小江回水區江段魚類群落結構的時空變動及驅動因素分析
2023-04-29 00:44:03鄭夢婷楊志胡蓮金瑤朱其廣鄒曦喬曄劉小燕唐會元
水生態學雜志 2023年3期
鄭夢婷 楊志 胡蓮 金瑤 朱其廣 鄒曦 喬曄 劉小燕 唐會元
摘要:調查小江下游回水區魚類資源,探討三峽水庫175 m正常蓄水后該江段魚類群落結構的時空變動特征及其驅動因素,為小江魚類資源保護提供參考依據。在小江下游渠口、養鹿、高陽和黃石設置4個采樣江段,1個位于水庫變動回水區,3個位于水庫常年回水區。基于2013年和2019年5-7月及10-11月開展的魚類資源調查數據,采用多元分析方法量化分析小江回水區江段非生物因子時空變動與魚類群落結構時空變動的關系。結果顯示:2013年和2019年共在小江回水區江段采集到魚類74種,隸屬于7目15科56屬,其中優勢種魚類15種,長江上游特有魚類7種,外來魚類5種;小江常年回水區的魚類群落結構在2019年發生了明顯改變,而變動回水區江段的魚類群落結構在2013年和2019年間無明顯差異;小江常年回水區江段的魚類群落結構在2013年顯示出明顯的季節變動特征,而在2019年季節性變動特征不明顯;變動回水區江段的魚類群落結構在2013年和2019年均存在明顯的季節變動;總磷含量、磷酸鹽含量、水溫和流速顯著影響小江回水區江段魚類群落結構的時空分布格局,其中磷酸鹽含量和水溫是最關鍵的驅動因素。盡量維持小江變動回水區的自然生境特征、采取措施控制小江常年回水區外來物種的數量以及嚴格控制外源營養物的輸入,對于保護小江土著魚類資源具有重要意義。
關鍵詞:魚類群落結構;環境因子;冗余分析;時間效應;小江
中圖分類號:S932.4? ? ? ? 文獻標志碼:A? ? ? ? 文章編號:1674-3075(2023)03-0042-12
河流筑壩形成了攔河水庫,原有的自然流水生境轉變為流水、緩流水以及靜水區梯次排列的生境,并迅速改變河流魚類群落結構的時空分布格局(Clavero et al, 2010;Pelicice et al, 2015;Poff, 2018)。探討魚類群落結構在庫區不同生境江段以及短期和長期時間尺度上的變動特征,一直是水生態學研究的重要內容之一(Gao et al, 2010;楊志等, 2015;Loures & Pompeu, 2019;Yang et al, 2021)。研究發現,水庫蓄水后,魚類群落的種類組成及其結構在空間縱向梯度上會沿著水庫河流帶到湖沼帶進行重新分布,并或多或少呈現隨庫區水位和蓄水運行周期變動的特征(Ferrareze et al, 2014;Lin et al, 2019)。許多喜流水性的魚類種類會根據庫區水位的周期性變動在水庫的河流帶和過渡帶之間進行遷徙,而適應靜水生境的肉食性魚類和浮游生物食性魚類則往往常年在水庫湖沼帶魚類群落中占據主導地位(Lin et al, 2019;Yang et al, 2021)。研究也發現,造成庫區不同江段魚類群落結構時空格局變動的因素,除魚類的遷徙特征和上游梯級水庫蓄水運行的綜合影響效應以外,庫區不同江段間的縱向梯度生境差異(如流速、營養鹽差異等)也起著重要的作用(Terra et al, 2011;Loures & Pompeu, 2019;Yang et al, 2021)。盡管如此,由于庫區不同支流在流域、局域和河段尺度上生境組成的差異以及在魚類種類組成上的差異,不同支流魚類群落結構在時空尺度上的變動特征及其驅動因素很可能也存在明顯的差異(劉燕山等, 2021)。在此背景下,對特定支流魚類群落結構的時空變動特征及其驅動因素進行研究,對于該支流魚類資源的保護具有重要的意義。
小江(又名澎溪河)是三峽庫區消落區面積最大、變動回水區較長以及常年流量較大的一條支流(史方等, 2017),孕育著豐富多樣的魚類資源(葉學瑤等, 2017),不僅是許多適應庫區靜緩流生境魚類的棲息、覓食和產卵區域(阮瑞等, 2017),也是許多產漂流性卵魚類的產卵區域(陳小娟等, 2020)。迄今,關于小江魚類群落結構的研究主要涉及種類組成及生物多樣性在蓄水前后的變動(葉學瑤等, 2017)以及部分區域(如漢豐湖等)魚類群落結構的季節性變動特征(丁慶秋等, 2015; 王敏等, 2017),而對其回水區江段群落結構在三峽水庫175 m正常蓄水后(2012年后)的時空變動特征及其驅動因素的定量分析研究缺乏。本文在小江下游回水區4個江段開展魚類資源調查,探討三峽水庫175 m正常蓄水后該江段魚類群落結構的時空變動特征及其驅動因素,以期為小江魚類資源保護提供參考依據。
1? ?材料與方法
1.1? ?研究區域
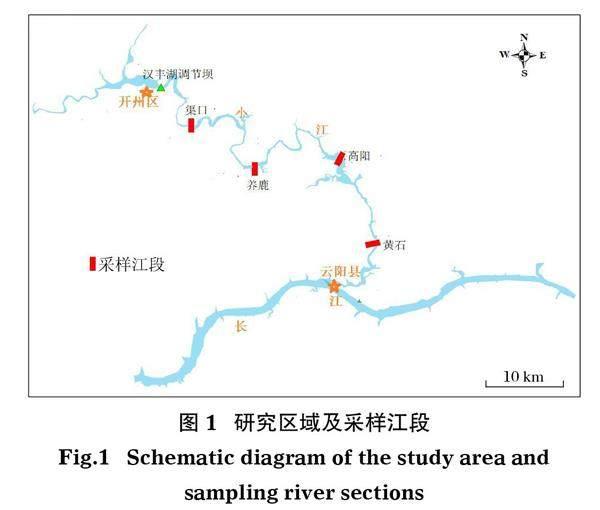
研究區域位于三峽庫區小江下游回水區河段(圖1)。小江全長182.4 km,流域面積5 172.5 km2,回水區長度約117.5 km,包括開州區漢豐湖調節壩以下江段。在三峽水庫低水位運行時,漢豐湖調節壩至渠口鎮下游白家溪江段為流水江段,而白家溪以下江段為靜緩流生境江段。采樣江段有4個,從上游到下游分別為渠口、養鹿、高陽和黃石江段,其中渠口采樣江段位于水庫變動回水區,該區域的底質基質主要由礫石和卵石組成,其他3個采樣江段位于水庫的常年回水區,底質基質主要由淤泥及大型巖石構成;高陽、養鹿和渠口江段均具有大面積的港灣生境,在春夏季時這些港灣覆蓋有大面積的水草植被,為許多粘草產卵魚類產卵場的主要分布區域(阮瑞等, 2017)。
1.2? ?樣本采集及環境因子測定
2013年和2019年5-7月以及10-11月,分別在4個采樣江段(渠口、養鹿、高陽和黃石)開展魚類資源調查(圖1)。2013年為三峽水庫達到175 m正常蓄水位后的第1年,而2019年為三峽水庫達到175 m正常蓄水位后的第7年。每次采樣,使用2種漁具(刺網和蝦籠)在每個采樣江段進行魚類樣本采集。刺網網目尺寸為20~250 mm(包括20、40、80、100、120、200和250 mm),不同網目規格的刺網網長為100~200 m,同一網目規格刺網的網長在不同斷面及不同月份間略有差異。刺網放置時,其一端被固定在沿岸帶,并沿著河流橫截面從沿岸帶放置到河道中間(Yang et al, 2021)。根據采樣點的最大水深,在黃石、高陽和養鹿江段放置網高為3、5、8、10、20、30和50 m的定置刺網,在渠口江段則放置網高為3、5、8和10 m的定制刺網,使得各個水層分布的魚類均能夠捕撈到。蝦籠被放置在非常淺的河道或巖石密布的區域,以捕捉小型營底棲生活的魚類。定置刺網和蝦籠開始放置時間通常為17:00-19:00,持續到第二天早上6:00-7:00,以覆蓋魚類活動最頻繁的時間段(Blabolil et al, 2017)。每次采樣時,相同網目規格刺網以及蝦籠在每個采樣江段的捕撈努力量大致相等。對所有采集到的魚類標本進行種類鑒定,并測量每尾個體的全長、體長(精確到1 mm)和體重(精確到0.1 g)(丁慶秋等, 2015)。
在漁獲物采樣的同時,使用LGY II型流速儀和多參數水質分析儀YSI 6600現場測量各個采樣江段的斷面平均流速(m/s)、水溫(WT,℃),溶解氧(DO,mg/L)和pH。同時,魚類資源調查期間,采集并測定各個采樣點的總氮(TN,mg/L)、總磷(TP,mg/L)、氨氮(AN,mg/L)、硝氮(NN,mg/L)、磷酸鹽(PP,mg/L)、高錳酸鹽指數(CODMn,mg/L)和葉綠素a(Chl-a,μg/L)含量。
1.3? ?數據統計與分析
對采集到的所有魚類的拉丁名,按照Eschmeyers Catalog of Fishes(Fricke et al, 2020)進行校對和更正,并確定所有魚類的流速偏好(曹文宣等, 2007;梁秩燊等, 2019)。同時,計算每個采樣江段魚類的相對豐度;計算魚類在漁獲物中的數量和重量比例,確定比例大于2%的主要種類組成。
基于Bray-Curtis相似性矩陣,采用組平均連接的聚類分析(Cluster analysis,CA)方法來確定魚類群落結構的潛在空間和時間分組(Clarke & Gorley, 2006)。在CA之前,為了避免稀有物種的不確定性影響,將每個采樣點相對豐度小于0.5%的魚類物種排除(Yang et al, 2018)。Bray-Curtis相似矩陣數據進行lg(x+1)轉換。采用One-way ANOSIM方法檢驗魚類群落結構在各組之間的差異是否在統計學上顯著,并采用百分比相似性分析(Similarity Percentage,SIMPER)獲得引起不同組間魚類群落結構差異的主要種類。同時,使用以下公式計算這些主要種類在不同聚類組間的相對豐度的變化:
Rav = G1ra-G2ra? ?①
式中:Rav為主要種類在不同聚類組之間的相對豐度的變化;G1ra和G2ra分別為主要物種在聚類組1和聚類組2中的相對豐度(%)。
使用11個環境變量和魚類的相對豐度數據來分析生物-非生物關系。11個非生物變量,包括總氮(TN)、硝氮(NN)、氨氮(AN)、總磷(TP)、磷酸鹽(PP)、葉綠素a(Chl-a)、高錳酸鹽指數(CODMn)、水溫(WT)、pH、溶解氧含量(DO)和流速(V),被用作解釋變量,而相對豐度數據為響應變量。所有數據在建模前都經過lg(x+1)轉換。在生物-非生物關系分析前,首先采用去趨勢對應分析(Detrended correspondence analysis,DCA)確定使用線性還是非線性模型(Yang et al, 2021)。對本文的數據而言,DCA前4個軸的最大軸長度小于3(范圍為1.493~1.971),表明選擇線性模型是合適的。因此,我們使用冗余分析(Redundancy analysis,RDA)模型來分析生物-非生物關系(Borcard et al, 2011)。在RDA構建之前,采用方差膨脹系數(Variance inflation factor,VIF)檢驗各變量間的共線性;VIF值大于10的變量具有明顯的共線性,其將被排除在RDA分析中。RDA分析時,采用正向選擇方法選擇解釋變量,以獲得最簡潔的RDA模型(Borcard et al, 2011),并通過置換檢驗(permutation test)確定RDA全局模型、每個RDA標準軸和每個解釋變量的顯著性。隨后,在此基礎上,采用層次分割方法(Hierarchical partitioning,HP)評估不同變量解釋小江下游魚類群落結構變動的相對比例。
使用軟件PRIMER-E (Version 6) 和 PERMANOVA+ (Anderson et al, 2008)進行CA、ANOSIM 和SIMPER分析。使用r軟件包“vegan 2.5-4”、 “ggplot2 3.3.2”、“ggrepel 0.8.2”和“rdacca.hp 0.1.0”進行DCA、 VIF、RDA、Permutation test和HP分析。本文的顯著性水平設置為P<0.05。
2? ?結果與分析
2.1? ?魚類種類組成及主要種類
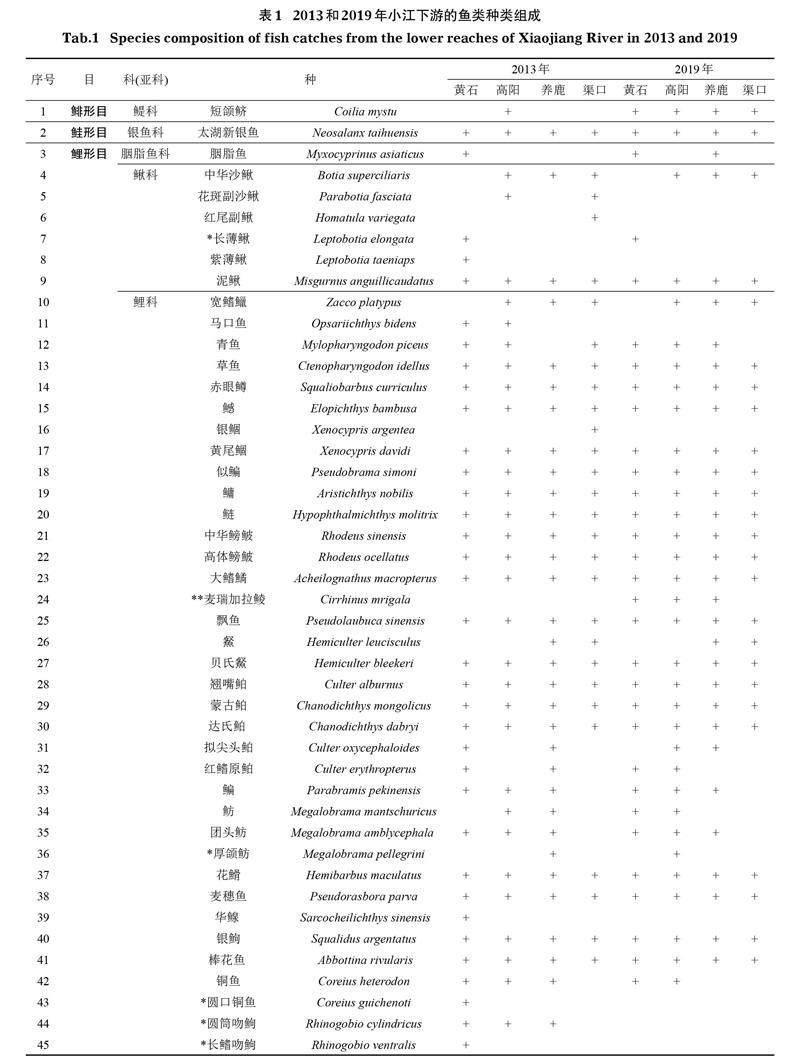
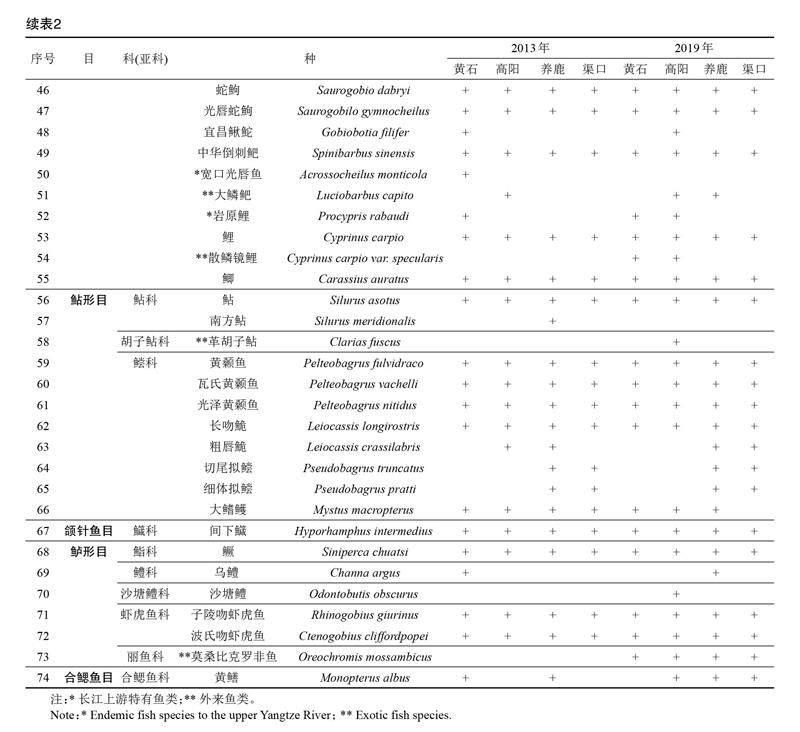
2013年和2019年共調查到漁獲物2 497.53 kg,47 305尾,鑒定出種類74種,隸屬于7目15科56屬(表1),其中2013年采集到魚類69種,2019年采集到魚類63種。2次調查中,采集到的鯉科魚類種類數最多,共有35屬46種,占總種類數的62.16%;其次為鲿科和鰍科,分別有4屬8種和5屬6種,各占10.81%和8.11%;最少為鳀科、銀魚科、胭脂魚科、胡子鲇科、鱵科、鮨科、鱧科、沙塘鱧科、麗魚科和合鰓魚科,均僅有1屬1種,各占1.35%(表1)。2次調查共采集到長江上游特有魚類7種,以及外來魚類5種(表1)。
2次調查,在漁獲物中數量百分比或重量百分比大于2%的魚類共有15種,如圖2。其中數量百分比最高的種類為似鳊(Pseudobrama simoni),占總采集尾數的28.74%,其次為蒙古鲌(Chanodichthys mongolicus)、飄魚(Pseudolaubuca sinensis)和光澤黃顙魚(Pelteobagrus nitidus),分別占10.06%、8.98%、8.93%和6.91%,最少為鰱(Hypophthalmichthys molitrix),占0.47%;重量百分比最高的種類為翹嘴鲌(Culter alburnus),占總重量的15.68%,其次為鯉(Cyprinus carpio)、蒙古鲌、鰱和似鳊,分別占13.13%、11.69%、9.06%和8.05%,最少為銀鮈(Squalidus argentatus),占0.43%(圖2)。
2.2? ?群落結構及其時空變動特征
2013年和2019年小江下游各江段魚類群落的聚類分析如圖3。在63.22%的Bray-Curtis相似性水平上可將2013年和2019年小江下游各江段的魚類群落結構分為3大組:組1包括黃石、高陽、養鹿江段2013年春夏季和秋冬季魚類,以及渠口江段2013年和2019年春夏季魚類;組2包括黃石、高陽、養鹿江段2019年春夏季和秋冬季魚類;組3包括渠口江段2013年和2019年秋季魚類。One-way ANOSIM檢驗顯示各大組間及任意兩大組間的魚類群落結構在統計學上差異顯著(各組間,r=0.711,P=0.1%,迭代次數999次;組1 vs組2,r=0.670,P=0.1%,迭代次數999次;組1 vs 組3,r=0.703,P=2.2%,迭代次數45次;組2 vs 組3,r=0.896,P=3.6%,迭代次數28次)。同時,組1又可以分為2小組,其中組1A包括黃石、高陽、養鹿江段2013年春夏季魚類,以及渠口江段2013年和2019年春夏季魚類,組1B包括黃石、高陽、養鹿江段2013年秋冬季魚類;組2也可以分為2小組,其中組2A包括黃石、高陽、養鹿江段2019年春夏季的魚類群落結構,而組2B包括黃石、高陽、養鹿江段2019年秋冬季的魚類群落結構(圖3)。One-way ANOSIM檢驗顯示魚類群落結構在組1A和組1B間差異顯著,而在組2A和組2B間差異不顯著(組1A vs組1B,r=0.723,P=1.8%,迭代次數56次;組2A vs 組2B,r=0.926,P=10.0%,迭代次數10次)。
用百分比相似性指數(SIMPER)分析組間群落結構差異(表2),結果顯示:引起組1和組2間魚類群落結構差異的主要物種共有18種,其中短頜鱭(Coilia mystu)在兩組間的豐度變動對組間魚類群落結構的差異貢獻率最高,為18.15%,其次為蒙古鲌 (6.96%)和蛇鮈 (6.84%);引起組2和組3間魚類群落結構差異的有17種,其中短頜鱭貢獻率最高,為12.14%,其次為似鳊 (11.30%)和光澤黃顙魚 (8.03%);引起組1和組3間魚類群落結構差異的共有15種,其中光澤黃顙魚貢獻率最高,為11.56%,其次為似鳊 (8.52%)和短頜鱭 (7.14%)。
2.3? ?群落結構與環境因子的關系
不同環境變量在小江各采樣江段不同采樣時間的平均值如表3所示。方差膨脹系數(VIF)檢驗顯示環境因子變量TN、NN和AN的值大于10(TN,64.516;NN,30.077;AN,119.450),表明與其他變量具有明顯的共線性,因此這些變量被排除,運用在后續的冗余分析中。通過變量的正向選擇,只有5個變量(TP、PP、Chl-a、WT和V)被最終選擇為RDA模型的解釋變量。這5個變量作為解釋變量在RDA中獲得的Adjust R2(0.484),幾乎與全變量模型獲得的Adjust R2(0.516)一樣大。最終的RDA模型在統計學上是顯著的(P<0.01),沒有明顯的共線性(各個變量的VIF值均小于10),具有2個顯著的規范軸(RDA1:F=9.750,P=0.001;RDA2:F=5.801,P=0.003)。所選的5個解釋變量中有4個解釋變量與小江下游4個采樣點魚類群落結構的時空變動顯著相關(TP,F=3.310,P=0.014; PP, F=5.720,P=0.001;WT,F=4.046,P=0.003;V, F=4.017, P=0.003)。最終的RDA分析結果見圖4。所選的5個解釋變量共同解釋了群落結構48.44%的變異,其中前2個規范軸共同解釋了群落結構39.46%的變異,第一個軸單獨解釋了群落結構24.74%的變異。
RDA1軸能夠將組2的魚類群落結構(2019年春夏季和秋冬季黃石、高陽和養鹿江段樣本)與組1(2013年春夏季和秋冬季黃石、高陽和養鹿江段樣本)和組3(渠口江段2013年和2019年秋季的魚類群落結構)的魚類群落結構分離開來,其中組2分布在RDA1>0的一側,而組1和組3則分布在RDA1<0的一側;組3以及組1B樣本分布在RDA2<0的一側,而組1B樣本分布在RDA2>0的一側,表明沿著RDA2軸2013年4個采樣江段的魚類群落結構呈現明顯的季節性變動特征;盡管如此,組2A和組2B樣本在RDA2=0的上下兩側均有分布,表明黃石、高陽和養鹿江段的魚類群落結構在2019年時的季節性變動特征不明顯。似鳊(PS)、銀鮈(SAR)、赤眼鱒(Squaliobarbus curriculus)(SCU)和蒙古鲌 (CM)等位于三序圖的左下角,其的相對豐度變動緊密聯系著更高的Chl-a和TP含量。巖原鯉(Procypris rabaudi)(PR)、圓口銅魚(Coreius guichenoti)(CG)、寬鰭鱲(Zacco platypus)(ZP)和華鳈(Sarcocheilichthys sinensis)(SSI)等位于靠近中心的左上角,其相對豐度在聚類組1春夏季樣本中最高,與較高的流速(V)和較高的水溫(WT)密切相關。同時,貝氏?(Hemiculter bleekeri)(HB)、光澤黃顙魚(PN)、蛇鮈(SD)等與變量WT正相關,表明這些魚類在水溫較高的時候大量出現;短頜鱭(CMY)、羅非魚(Oreochromis mossambicus)(OM)、黃顙魚(Pelteobagrus fulvidraco)(PF)等的相對豐度變動易受磷酸鹽(PP)含量變動的影響。其他種類如鯉(Cyprinus carpio)(CC),鲇(Silurus asotus)(SA)和瓦氏黃顙魚(Pelteobagrus vachelli)(PV)等聚集在三序圖的中央,表明它們能夠存在于水庫的大部分區域,并能夠適應不同采樣點的生境條件。總之,RDA分析的結果顯示第一個軸主要代表生物-非生物之間的年際變動關系(魚類群落結構在2次采樣間發生了明顯變動),而第二個軸主要描述生物-非生物之間的季節變動關系(圖4)。
RDA的層次分割(HP)結果顯示:磷酸鹽解釋的魚類群落結構時空變異比例最高,為34.58%,其次為和水溫和總磷,分別為25.31%和15.17%,最低為Chl-a,僅為10.76%(圖5)。
3? ?討論
3.1? ?魚類群落結構的時空變動特征
聚類分析以及One-way ANOSIM檢驗結果顯示在相同采樣季節,黃石、高陽和養鹿江段的魚類群落結構在不同年份間差異明顯,而渠口江段的魚類群落結構在不同年份間差異不明顯(圖3),表明小江常年回水區的魚類群落結構在三峽水庫175 m正常蓄水運行多年后發生了明顯的改變,而小江變動回水區江段(渠口江段)的魚類群落結構在2次調查間無明顯的差異。該結果與在三峽庫區干流常年回水區(Yang et al, 2021)以及變動回水區(Yang et al, 2012)獲得的結果相一致。魚類群落結構的年際變動在不同水文功能區域間的差異,很可能歸因于這2方面原因:(1)變動回水區在三峽水庫高、低水位時均具有一定的流速,該區域既分布有適應靜緩流生境的魚類種類,也分布有喜流水性的魚類種類,區域內的魚類群落結構易受上游江段流水性魚類下行補充的影響。尤其當變動回水區的優勢類群主要為喜流水性魚類時,只要這些魚類在上游自然連通的流水江段內有充足的群體,該變動回水區的魚類群落結構就會保持相對穩定(Yang et al, 2012; 2021);(2)常年回水區通常分布有大量適應靜緩流生境的魚類種類,雖然在一定程度上可以維持該區域魚類群落結構的穩定,但是當上游魚類群落結構發生變化時,由于生物間的相互作用,該區域的魚類群落結構也會受到影響而發生變動(董哲仁, 2010; Yang et al,2021)。除此之外,常年回水區通常是庫區非土著物種數量急劇增加的區域,其魚類群落結構更易受非土著物種增殖的影響(Lin et al, 2019)。正如本文表2所述,通過葛洲壩及三峽船閘引入的短頜鱭以及由于外來引種或放生進入的莫桑比克羅非魚(Oreochromis mossambicus)(楊志等, 2012)等非三峽庫區土著物種數量的劇烈變動主要發生在常年回水區江段。
同時,聚類分析以及One-way ANOSIM檢驗的結果也顯示常年回水區江段(黃石、高陽和養鹿)的魚類群落結構在2013年時有明顯的季節性變動特征,然而在2019年時季節性變動特征并不明顯(圖3),表明隨著三峽水庫正常蓄水運行的進行,常年回水區江段的魚類群落結構在不同季節間逐漸趨同。盡管如此,本文的結果也顯示變動回水區江段(渠口江段)的魚類群落結構在2013年和2019年均呈現明顯的季節性變動特征,表明變動回水區江段的魚類群落結構有其獨特的抗擾動和穩定機制,即靜水性和喜流水性魚類共存于不穩定的生境中,共同利用這個生境,從而使得該生境具有更多的稀有物種并最終達到群落的中度可持續性(Oliveira et al, 2003);生物與非生物因子共同構建魚類群落結構,在不穩定生境中,外界因子包括流速及水質的梯度等在魚類群落結構構建中的作用更顯著,其能夠避免常年回水區江段(更為穩定的生境)所面臨的生物間的顯著效應(如表2顯示外來物種、短頜鱭等非土著物種在渠口江段難以形成大規模的種群,并進而影響其種群的穩定)(Beesley & Prince, 2010)。這種機制在三峽庫區干流回水變動區江段也曾被觀測到(Yang et al, 2012)。
3.2? ?魚類群落結構與環境因子的關系
RDA結果顯示小江回水區江段魚類群落結構的時空變動與營養鹽含量(總磷、磷酸鹽)、葉綠素a、水溫及流速的時空變動密切相關,表明水質、水溫以及水文情勢特征在影響小江回水區江段魚類群落結構時空變化中起著重要的作用。許多研究已發現,水質特征包括總磷、磷酸鹽、溶解氧、pH、葉綠素a等顯著影響著河庫魚類群落結構的時空分布格局,且其關鍵水質影響要素隨著不同研究區域間的生境特征差異而有所不同(程琳, 2011; 孟睿等, 2013;劉燕山等, 2021;Lin et al, 2012; Gao et al, 2015; Yang et al, 2021)。在外源營養物質輸入較高的湖庫中,總磷和/或葉綠素a濃度通常是影響其魚類群落結構時空變動的關鍵因子之一(程琳, 2011; 孟睿等, 2013; Miranda et al, 2015; 周興安等, 2016; Yang et al, 2021)。有調查發現,小江回水區江段是三峽庫區營養鹽含量較高、浮游植物生物量較高的區域,因此小江回水區江段的魚類群落結構一定程度上會受到該區域營養鹽含量和浮游植物生物量變動的影響(陳薛偉杰, 2018)。本文中表2的結果也證實了這一點:調查區域的魚類主要為適應靜緩流生境或廣適性的魚類種類,而這些魚類種類的食物來源與其棲息區域營養鹽含量和浮游植物生物量存在密切的關系。
與TP相比,變量PP具有更高的解釋能力(圖5),表明小江回水區江段的魚類群落結構更易受磷酸鹽含量變動的影響。已有研究表明,磷酸鹽易被浮游植物吸收利用,是影響不同類型水域浮游植物數量消長的主要影響因子之一(李欽欽等, 2010;王麗卿等, 2011; 李佳俊等, 2015),因此磷酸鹽可以作為關鍵要素通過浮游植物-魚類這一上行效應影響魚類群落的結構特征。由于三峽水庫的磷酸鹽主要來源于隨降雨、徑流作用輸入的生活和生產性排污,包括農業面源輸入、城鎮污水輸入以及工業生產排污輸入等(曹承進等, 2008),因此通過控制污水輸入,很可能可以調控小江回水區江段的魚類群落結構,從而使得小江回水區江段魚類生物多樣性得以增加(例如環境耐受性高的部分外來物種數量的減少,以及水質要求高的部分土著物種數量的增加等)。
RDA結果顯示水溫主要影響調查區域魚類群落結構的季節分布特征,且這種影響主要與鯉、蛇鮈、鳊、光澤黃顙魚等在春夏季產卵的魚類種類的數量變動有關。已有研究表明,水溫是促使魚類產卵行為發生的驅動因素之一(Kayaba et al,2010;King et al,2016),而魚類產卵行為的發生又會使得魚類發生聚集行為,從而在特定時間特定江段內出現大量集群群體(Tao et al, 2017)。在本文中,許多魚類在春夏季節到小江回水區沿岸帶水草基質或流水淺灘上產卵繁殖,然而其在秋冬季時又會回到三峽水庫干支流深水灣沱處越冬,從而使得這些魚類在小江回水區江段的種群數量在不同季節間呈現明顯的變動特征。研究已發現,三峽水庫干支流回水區江段的魚類群落結構的季節性變動明顯,且該種變動與部分魚類受水位和水溫變動顯著影響的季節性遷徙有關(楊少榮等, 2010)。盡管如此,本文的結果還顯示在2019年時,常年回水區的3個采樣江段的魚類群落結構的季節性變動變得不明顯,這很可能與這些區域生境趨于相對穩定、定居性魚類比例較高,而遷徙性魚類比例較低有關。
RDA結果顯示流速顯著影響小江回水區江段魚類群落結構的時空變動,該結果與Yang 等(2021)在三峽水庫干流回水區江段獲得的結果相一致。盡管如此,與三峽庫區干流相比,流速對小江回水區江段魚類群落結構時空變動的解釋能力較弱(圖5),這很可能與小江回水區各調查江段流速梯度不明顯有關:三峽水庫低水位運行時,下游3個調查江段位于常年回水區,其上游及區間來水量通常較小,難以在此3個江段間產生明顯的流速梯度;三峽水庫高水位運行時,4個調查江段均位于小江回水倒灌區內,該回水區末端直達漢豐湖調節壩壩址,只有當調節壩放水時才會產生一定的流速梯度。該結果突顯了在辨識影響不同采樣江段魚類群落結構時空變動的關鍵生境因子時,需要充分考慮采樣江段間所存在的生境特征差異。
3.3? ?研究結果在小江下游魚類資源保護中的運用
本文的研究結果顯示處于變動回水區內的渠口江段的魚類群落結構在三峽水庫正常蓄水多年后仍保持穩定,而其他3個處于常年回水區內的江段的魚類群落結構則顯示相反的結果,表明小江變動回水區江段在保持原有魚類物種多樣性方面具有更大的作用。尤其是相較于渠口江段,下游3個江段的非土著物種(如短頜鱭、莫桑比克羅非魚等)在漁獲物中的相對豐度更大,其群落結構更易受到非土著物種特別是外來物種數量變動的影響。在上述背景下,盡量維持小江變動回水區的自然生境特征(如在三峽水庫低水位運行時,盡量維持其流水生境長度;在高水位運行時,盡量維持一定長度的流水緩沖區),以及采取措施控制小江常年回水區外來物種的數量對于保護小江回水區江段土著魚類資源具有重要的意義。
同時,RDA分析結果顯示營養鹽含量的時空變動對小江回水區江段魚類群落結構的時空變化有顯著的影響。因此,通過外源營養物的輸入控制,很可能會改變小江回水區江段營養物質的時空分布格局,從而促進小江回水區江段魚類群落結構的完善(如物種生物多樣性的上升、重要種質魚類數量的增加等)。此外,本文結果還顯示小江回水區江段的魚類群落結構在三峽水庫正常蓄水多年后仍在發生變動,表明在研究區域開展長期的魚類資源監測是非常有必要的。
參考文獻
曹承進, 秦延文, 鄭丙輝, 等, 2008. 三峽水庫主要入庫河流磷營養鹽特征及其來源分析[J]. 環境科學, 29(2): 310-315.
曹文宣, 常劍波, 喬曄, 等, 2007. 長江魚類早期資源[M]. 北京: 中國水利水電出版社.
程琳, 2011. 長江流域湖泊魚類群落與主要環境因子關系及其預測模型[D]. 北京: 中國科學院大學.
陳薛偉杰, 2018. 三峽庫區萬州江段和小江魚類食物網結構與功能研究[D]. 貴陽: 貴州大學.
陳小娟, 唐會元, 楊志, 等, 2020. 三峽水庫支流小江魚類早期資源現狀[J]. 三峽生態環境監測, 5(1): 42-47.
代亮亮, 李莉杰, 何梅, 等, 2020. 貴州草海秋季浮游植物群落結構與水質因子的關系[J]. 水生態學雜志, 41(2): 64-69.
丁慶秋, 彭建華, 楊志, 等, 2015. 三峽水庫高、低水位下漢豐湖魚類資源變化特征[J]. 水生態學雜志, 36(3): 1-9.
董哲仁, 孫東亞, 趙進勇, 等, 2010. 河流生態系統結構功能整體性概念模型[J]. 水科學進展, 21(4): 550-559.
李佳俊, 沈萍萍, 譚燁輝, 等, 2015. 南海東北部浮游植物對氮、磷加富的響應及與不同水團的關系[J]. 海洋學報, 37(10): 90-101.
李欽欽, 鄧建才, 胡維平, 等, 2010. 太湖金墅灣水源地浮游植物群落結構及其與環境因子的關系[J]. 應用生態學報, 21(7): 1844-1850.
劉燕山, 谷先坤, 唐晟凱, 等, 2021. 東太湖魚類群落結構特征及其與環境因子的關系[J]. 生態學報, 41(2): 769-780.
梁秩燊, 易伯魯, 余志堂, 2019. 江湖魚類早期發育圖志[M]. 廣州: 廣東科技出版社.
孟睿, 何連生, 過龍根, 等, 2013. 長江中下游草型湖泊浮游植物群落及其與環境因子的典范對應分析[J]. 環境科學, 34(7): 2588-2596.
阮瑞, 張燕, 沈子偉, 等, 2017. 三峽消落區魚卵、仔稚魚種類的鑒定及分布[J].中國水產科學, 24(6): 1307-1314.
史方, 陳小娟, 楊志, 等, 2016. 三峽水庫小江流域魚類營養層次研究[J]. 水生態學雜志, 37(4): 70-77.
王麗卿, 許莉, 盧子園, 等, 2011. 淀山湖浮游植物數量消長及其與環境因子的關系[J]. 環境科學, 32(10): 2868-2874.
王敏, 朱峰躍, 劉紹平, 等, 2017. 三峽庫區漢豐湖魚類群落結構的季節變化[J]. 湖泊科學, 29(2): 439-447.
楊少榮, 高欣, 馬寶珊, 等, 2010. 三峽庫區木洞江段魚類群落結構的季節變化[J]. 應用與環境生物學報, 16(4): 555-560.
楊志, 陶江平, 唐會元, 等, 2012. 三峽水庫運行后庫區魚類資源變化及保護研究[J]. 人民長江, 43(10): 62-67.
楊 志, 唐會元, 朱迪, 等, 2015. 三峽水庫175 m 試驗性蓄水期庫區及其上游江段魚類群落結構時空分布格局[J]. 生態學報, 35(15): 5064-5075.
葉學瑤, 陶敏, 朱光平, 等, 2017. 三峽庫區小江魚類群落特征及其歷史變化[J]. 長江流域資源與環境, 26(6): 841-846.
周興安, 喬永民, 王賽, 等, 2016. 洱海魚類群落結構特征及其與環境因子關系[J]. 生態學雜志, 35(6): 1569-1577.
Beesley L S, Prince J, 2010. Fish community structure in an intermittent river: the importance of environmental stability, landscape factors and within-pool habitat descriptors[J]. Marine & Freshwater Research, 61(5): 605-614
Blabolil P, [R]íha M, Ricard D, et al, 2017. A simple fish-based approach to assess the ecological quality of freshwater reservoirs in Central Europe[J]. Knowledge & Management of Aquatic Ecosystems, 418:53.
Borcard D, Gillet F, Legendre P, 2011. Numerical Ecology with R[M]. New York: Springer.
Clarke K R, Gorley R N, 2006. PRIMER V6: User manual/tutorial PRIMER-E[M]. Plymouth,UK: PRIMER-E.
Clavero M, Blanco-Garrido F, Prenda J, 2010. Fish fauna in Iberian Mediterranean river basins: Biodiversity, introduced species, and damming impacts[J]. Aquatic Conservation Marine & Freshwater Ecosystems, 14(6): 575-585.
Ferrareze M, Casatti L, Nogueira M G, 2014. Spatial heterogeneity affecting fish fauna in cascade reservoirs of the Upper Paraná Basin, Brazil[J]. Hydrobiologia, 738(1): 97-109.
Fricke R, Eschmeyer W N, Van der Laan R, 2020. Eschmeyer's catalog of fishes: Genera, species, references[P]. http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp.
Gao X, Zeng Y, Wang J W, et al, 2010. Immediate impacts of the second impoundment on fish communities in the Three Gorges[J]. Environmental Biology of Fishes, 87(2): 163-173.
Gao X, Zhang Y, Ding S, et al, 2015. Response of fish communities to environmental changes in an agriculturally dominated watershed (Liao River Basin) in northeastern China[J]. Ecological Engineering, 76: 130-141.
Kayaba T, Sugimoto T, Mori T, et al, 2010. Induced spontaneous spawning using an increased temperature stimulus in the cultured barfin flounder Verasper moseri[J]. Fisheries Science, 69(4): 663-669.
King A J, Gwinn D C, Tonkin Z, et al, 2016. Using abiotic drivers of fish spawning to inform environmental flow management[J]. Journal of Applied Ecology, 53: 34-43.
Lin C, Lek S, Lek-Ang S, et al, 2012. Predicting fish assemblages and diversity in shallow lakes in the Yangtze River Basin[J]. Limnologica, 42(2): 127-136.
Lin P C, Gao X, Liu F, et al, 2019. Long-term monitoring revealed fish assemblage zonation in the Three Gorges Reservoir[J]. Journal of Oceanology and Limnology, 37: 1258-1267.
Loures R C, Pompeu P S, 2019. Temporal changes in fish diversity in lotic and lentic environments along a reservoir cascade[J]. Freshwater Biology, 64(10): 1806-1820.
Miranda L E, Bies J M, Hann D A, 2015. Land use structures fish assemblages in reservoirs of the Tennessee River[J]. Marine & Freshwater Research, 66(6): 526-534
Pelicice F M, Pompeu P S, Agostinho A A, 2015. Large reservoirs as ecological barriers to downstream movements of neotropical migratory fish[J]. Fish and Fisheries, 16(4): 697-715.
Oliveira E F, Goulart E, Minte-Vera C V, 2003. Patterns of dominance and rarity of fish assemblage along spatial gradients in the Itaipu Reservoir, Paraná, Brazil [J]. Acta scientiarum: Biological sciences, 25(1):71-78.
Poff N L, 2018. Beyond the natural flow regime? Broadening the hydro-ecological foundation to meet environmental flow challenges in a non-stationary world[J]. Freshwater Biology, 63(8): 1011-1021.
Tao J P, Yang Z, Cai Y P, et al, 2017. Spatiotemporal response of pelagic fish aggregations in their spawning grounds of middle Yangtze to the flood process optimized by the Three Gorges Reservoir operation[J]. Ecological Engineering, 103: 86-94.
Terra B, Santos A, Araújo F G, 2011. Fish assemblage in a dammed tropical river: an analysis along the longitudinal and temporal gradients from river to reservoir[J]. Neotropical Ichthyology, 8(3): 599-606.
Yang S R, Gao X, Li M Z, et al, 2012. Interannual variations of the fish assemblage in the transitional zones of the Three Gorges Reservoir: Persistence and stability[J]. Environmental Biology of Fishes, 93(2): 295-304.
Yang Z, Tao J P, Qiao Y, et al, 2018. Multivariate analysis performed to identify the temporal responses of fish assemblages to abiotic changes downstream of the Gezhouba Dam on the Yangtze River[J]. River Research and Applications, 34(9): 1142-1150.
Yang Z, Pan X J, Hu L, et al, 2021. Effects of upstream cascade dams and longitudinal environmental gradients on variations in fish assemblages of the Three Gorges Reservoir[J]. Ecology of Freshwater Fish, 30(4): 503-518.
(責任編輯? ?鄭金秀)
Spatiotemporal Variation of Fish Community Structure and
Its Driving Factors in the Backwater Area of Xiaojiang River
ZHENG Meng‐ting1, YANG Zhi2, HU Lian2, JIN Yao2, ZHU Qi‐guang2, ZOU Xi2,
QIAO Ye2, LIU Xiao‐yan1, TANG Hui‐yuan2
(1. College of Animal Science and Technology, Hunan Agricultural University,
Changsha? ?410128, P.R. China;
2. Key Laboratory of Ecological Impacts of Hydraulic-projects and Restoration of
Aquatic Ecosystem of Ministry of Water Resources, Institute of Hydroecology,
Ministry of Water Resources and Chinese Academy of Sciences, Wuhan? ?430079, P.R. China)
Abstract:In this study, we explored the temporal and spatial variation characteristics of fish community structure and their driving factors in the backwater area of Xiaojiang River after the normal storage (175m after 2012) and operation of the Three Gorges Reservoir. It aimed to provide reference for the fish resource protection in the backwater area of Xiaojiang River. The study was based on the fish resource survey and habitat data collection conducted in four sections (Qukou in the variable backwater area , Yanglu, Gaoyang and Huangshi sections in the perennial backwater area) of the Xiaojiang River from May to July in 2013 and from October to November in 2019, and the relationship between the abiotic factors and the changes of fish community structure was quantitatively analyzed using multivariate analysis method. Results show that a total of 74 fish species belonging to 56 genera, 15 families and 7 orders were collected from the backwater area of Xiaojiang River in 2013 and 2019, including 15 dominant species, 7 endemic species and 5 exotic species. There were 69 fish species collected in 2013, while there were 62 fish species collected in 2019. The fish community structure in the perennial backwater area of Xiaojiang River has changed significantly in 2019, while no significant difference in the fish community structures in the river section of the variable backwater area was observed between 2013 and 2019. The fish community structure in the perennial backwater area of Xiaojiang River showed obvious seasonal variation in 2013, while the seasonal variation became less obvious in 2019. The fish community structure in the variable backwater area of Xiaojiang River showed obvious seasonal variation in 2013 and 2019. The total phosphorus content, phosphate content, water temperature and flow velocity significantly affected the temporal and spatial change pattern of fish community structure in the backwater area of Xiaojiang River, and the phosphate content and water temperature were two key factors driving the change of fish community structure. Our results indicate that maintaining the natural habitat characteristics of the variable backwater area of Xiaojiang River, taking measures to control the population size of alien species in the perennial backwater area of the Xiaojiang River as well as strictly controlling the input of exogenous nutrients are of great significance to protect the indigenous fish resources in the backwater area of Xiaojiang River.
Key words:fish community structure; environmental factor; redundancy analysis; temporal effect; Xiaojiang River
收稿日期:2021-07-28? ? ? 修回日期:2023-01-11
基金項目:國家重點研發計劃課題(2019YFC0408904);國家自然科學基金(51779157);湖南省自然科學基金面上項目(2021JJ30336);中國長江三峽集團有限公司科研項目“三峽庫區產黏沉性卵魚類自然繁殖的生態調度需求及調度方式研究”。
作者簡介:鄭夢婷,1995年生,女,碩士研究生,主要研究方向為漁業資源與環境。E-mail:247115837@qq.com
通信作者:唐會元。E-mail: Tanghy@mail.ihe.ac.cn
