青海湖入湖河流大型底棲動物時空分布及漁產潛力估算
2023-04-29 03:38:29謝廷鋒和雅靜趙永晶徐東坡俞錄賢王洪鑄崔永德
水生態學雜志 2023年3期
謝廷鋒 和雅靜 趙永晶 徐東坡 俞錄賢 王洪鑄 崔永德
摘要:探究青海湖及其入湖河流底棲動物的群落結構與時空分布,根據底棲動物現存量估算漁產潛力,可為青海湖裸鯉(Gymnocypris przewalskii)的增殖放流和種群持續恢復提供科學依據。以青海湖5條主要入湖河流(黑馬河、布哈河、泉吉河、沙柳河、哈爾蓋河)為研究對象,于2019年在青海湖裸鯉洄游前期、中期和后期對18個斷面進行采樣調查。結果表明,調查共記錄底棲動物101種(屬),隸屬于5門7綱9目44科。空間分布上,黑馬河種類最多,有56種(屬),哈爾蓋河最少僅27種(屬),且EPT(蜉蝣目、襀翅目和毛翅目)自上游至下游呈減少趨勢;密度和生物量均以泉吉河最高(544 個/m2,4.77 g/m2),生物多樣性各指數以黑馬河最高。時間尺度上,青海湖裸鯉洄游前期、中期和后期,大型底棲動物種類數無明顯變化,但密度和生物量均呈增加趨勢,生物多樣性各指數呈降低趨勢。影響大型底棲動物密度和生物量分布的主要因素為底質類型、海拔高度、水溫、濁度和電導率(P?0.05)。基于大型底棲動物估算的漁產潛力以布哈河最高(285 129 kg),其次是泉吉河(20 694 kg)和沙柳河(18 857 kg),與青海湖裸鯉密度及產卵場空間分布較為一致,這與親魚洄游產卵需要大量的食物來源有很大關系。
關鍵詞:底棲動物;時空分布;漁產潛力;青海湖裸鯉
中圖分類號:Q178? ? ? ? 文獻標志碼:A? ? ? ? 文章編號:1674-3075(2023)03-0070-09
青海湖流域位于青藏高原東部,是我國生態環境地位極其重要但同時又很脆弱的區域。由于自然環境條件變化和人為活動的綜合影響,流域內生態環境不斷惡化,主要表現為青海湖水質惡化、水土流失嚴重、珍稀野生動物瀕臨滅絕和漁業資源瀕臨枯竭(馮宗煒和馮兆忠,2004)。長期過度捕撈導致魚類資源量銳減、魚體變小、魚類洄游通道受阻、天然產卵場喪失、資源再生能力下降(史建全等,2000;2004)。
青海湖裸鯉(Gymnocypris przewalskii)是青海湖流域特有魚類,占漁獲物總量的95%以上(陳民琦等,1990);其在咸水和淡水中均能生長,但生長緩慢,個體重達到500 g以上需要10年(中國科學院西北高原生物研究所,1989)。青海湖裸鯉具有明顯的生殖洄游特性,入湖河流是親魚洄游產卵的重要通道和場所,每年4-7月溯河產卵,產卵場主要在黑馬河、布哈河、泉吉河、沙柳河和哈爾蓋河等入湖淡水河流,通常在礫石河灘處產卵,但繁殖力低(張信等,2005)。不同時期的裸鯉食性不同,幼魚階段對餌料有較為明顯的選擇性,成體階段幾乎攝食水中所有生物性食物(青海省生物研究所,1975)。蔣小明(2012)認為大型底棲動物在食物網中具有承上啟下的作用,并成為許多魚類的優質天然餌料;汪洋(2019)研究發現,搖蚊幼蟲輔助飼喂明顯提高青海湖裸鯉仔稚魚增重率。
有關青海湖入湖河流大型底棲動物分布格局及時空動態的研究不多(孟星亮等,2014),漁產潛力也僅限于青海湖湖區(楊洪志和王基琳,1997;姚維志等,2011);而入湖河流作為青海湖裸鯉重要的產卵場亟需關注。國內關于漁產潛力估算的研究多針對湖泊(閆云君,1998),河流漁產潛力的研究相對較少。本文以青海湖5條入湖河流為研究對象,描述了大型底棲動物群落結構的時間動態與空間變化特征,計算大型底棲動物的現存量,并根據經驗模型,估算入湖河流大型底棲動物的漁產潛力,以期為青海湖裸鯉的人工放流和種群資源恢復以及流域內生物多樣性保護與生態治理提供技術支撐和科學依據。
1? ?材料與方法
1.1? ?區域概況
青海湖流域西北部面積遼闊,河網較為密集,較大的河流多分布于此;流域東南部陸地較為狹窄,河流短小,多為時令河,水量貧乏。湖周直接入湖且流域面積大于5 km2的河流有48條,本研究選取其中較大的5條河流,即黑馬河、布哈河、泉吉河、沙柳河和哈爾蓋河,這5條河流水量占據青海湖入湖總流量的80%(Li et al, 2007)。黑馬河全長20 km,河道平均比降5.3%;布哈河全長286 km,河道落差1 318 m,河床多為砂卵石組成;泉吉河全長65 km,河道落差1 113 m,平均比降1.7%,上中游河道順直,中游河床由砂卵石組成,下游流經廣闊的湖濱灘地;沙柳河全長106 km,河道落差1 505 m,中上游河床主要由大卵石組成,下游河道比較平緩。5條河流汛期(6-9月)主要依靠降水補給(李岳坦等,2010;時盛博等,2017)。
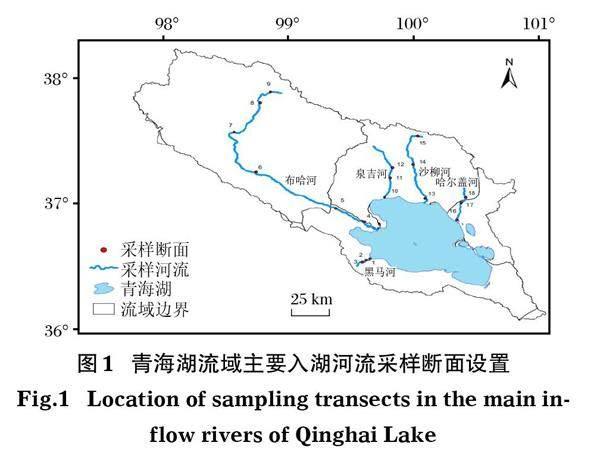
1.2? ?采樣斷面設置
本研究分別于2019年在5條入湖河流的上游、中游及下游共設置18個采樣斷面,進行大型底棲動物調查(圖1)。 調查時段分別為青海湖裸鯉洄游前期(5月)、洄游中期(7月)和洄游后期(8月)。
1.3? ?樣品采集與處理
在每個采樣斷面100 m 范圍內,用篩網孔徑 420 μm、網口寬度 30 cm 的D型抄網采集大型底棲動物。定量樣品在不同生境中采集,總樣方面積為0.9 m2,定性樣品通過D型抄網和手撿方式獲得。采集到的底棲動物帶回室內進行人工挑揀,分揀出的底棲動物標本放入樣品瓶中,并加入75%的酒精固定保存。樣品帶回實驗室后,參考相關文獻資料(劉月英等,1979;Morse et al, 1994;王洪鑄,2002;唐紅渠,2006;崔永德,2008;何雪寶,2011;周長發等,2015),在體視鏡和顯微鏡下進行種類鑒定、準確計數后,先用吸水紙吸干樣品表面的液體,再用精度為萬分之一的電子天平稱重(軟體動物為帶殼濕重)。
1.4? ?環境因子測定
使用手持GPS測定海拔高度(Altitude);通過哈希HQ40D便攜式多參數分析儀對電導率(Cond)、溶解氧(DO)、水溫(WT)、pH等水體理化指標進行現場測定;濁度(Tur)使用哈希2100Q便攜式濁度儀現場測定;水深(WD)用測深尺現場涉水測量。在采樣點的表、底層取水樣混合,帶回實驗室分析,化學需氧量(COD)、總氮(TN)、總磷(TP)、氨氮(NH4-N)、硝酸氮(NO3-N)、亞硝酸氮(NO2-N)等營養鹽,根據《水和廢水監測分析方法》測定(國家環保局,1989)。底質類型劃分依據Barbour等(1999)的標準。
1.5? ?數據分析
采用多元統計軟件PRIMER 6.1計算青海湖流域5條入湖河流大型底棲動物的Margalef豐富度指數(d)、Shannon-Wiener 多樣性指數(H')、Pielou均勻度指數(J)、Simpson多樣性指數(D)和優勢度指數(Y),計算公式(Shannon & Weaver, 1949; Pielou, 1966; Margalef, 1968)如下:
式中:S 為樣品物種總數,N 為總個體數,Pi為第 i 種的個體數與總個體數的比值,fi為第i 種在各點位的出現頻率,Y≥0.02 的種類確定為優勢種。
為探討底棲動物群落結構與環境因子之間的關系,采用Canoco 5.1對數據進行約束性排序。分析前對環境參數和底棲動物密度、生物量數據進行lg(x+1)轉化,以使數據呈正態分布,基于蒙特卡羅檢驗(Monte Carlo permutation test),通過前選法(Forward selection)排除貢獻小的因子。采用典范相關分析(Canonical Correspondence Analysis, CCA)探究底棲動物群落結構與篩選出來的環境因子之間的關系。
1.6? ?漁產潛力估算
漁產潛力(F)由動物可利用的資源量(R),即其生產量(P,一般以年計)與其現存量(B)的差額,乘以其對魚的轉化效率(C,餌料系數之倒數)得到(閻云君等,2005)。基于以上算法的經驗公式(王洪鑄等,2005)如下:
Ftotal = FO+FG+FI = 0.7BO+0.4BG+0.4BI? ?⑥
式中:Ftotal 為總漁產潛力,FO、FG、FI分別為寡毛類、鉤蝦、水生昆蟲的漁產潛力,BO、BG、BI分別為寡毛類、鉤蝦、水生昆蟲的現存量。鉤蝦的換算系數參考水生昆蟲。
根據《中國河湖大典》中提供的河長及河寬(不同河段均值)數據,計算出5條河流的面積,結合本次調查得到的平均生物量,計算各類群的現存量。調查已知漁獲物中青海湖裸鯉平均體重為177.7 g/尾,計算底棲動物對各河流洄游親魚的載魚量時,以此為參照。
2? ?結果
2.1? ?群落組成的時空分布
本次調查共采集到大型底棲動物101種(屬),隸屬于5門7綱9目44科。其中,寡毛類6科12種(屬),占總數的11.9%;水生昆蟲26科86種屬,占總數的85.1%;軟體動物、線蟲和渦蟲各有1種,共占總數的3%。空間分布上,黑馬河大型底棲動物種類數最高,有56種(屬),泉吉河次之,有49種(屬),哈爾蓋河最低,為27種(屬)(圖2);自上游到下游,EPT分類單元(蜉蝣目、襀翅目和毛翅目)減少,鉤蝦出現頻率增加。時間動態上,從青海湖裸鯉洄游前期至后期,大型底棲動物種類數呈現遞減的趨勢。前期最高為101種(屬),水生昆蟲為86種屬,占83%;中期為59種(屬),水生昆蟲47種(屬),占79%;后期最低,為54種(屬),水生昆蟲44種(屬),占81%。
2.2? ?密度和生物量
從底棲動物總密度的時空分布來看(圖3-a),在青海湖裸鯉洄游后期最高,各樣點平均密度為362 個/m2,中期次之,為253 個/m2,前期最低,平均密度只有149 個/m2。除哈爾蓋河以外,其他河流的總密度變化趨勢相一致,從前期至后期逐漸上升。從空間上看,泉吉河大型底棲動物全年各時期總密度均最高,平均密度達544 個/m2,布哈河和沙流河次之,哈爾蓋河最低,平均密度為85 個/m2。
從底棲動物總生物量的時空分布來看(圖3-b),裸鯉洄游后期最高,平均生物量為3.82 g/m2,中期最低,僅0.96 g/m2。各條河流平均生物量在3個時期無明顯變化規律。空間分布上,泉吉河全年總生物量最高,為4.77 g/m2,布哈河與沙柳河次之,哈爾蓋河最低,僅0.31 g/m2。
2.3? ?優勢種及多樣性時空變化
以優勢度Y≥0.02為標準,青海湖裸鯉洄游前期、中期和后期的優勢種分別為5種、4種和3種(表1)。其中,寡毛綱和軟甲綱各1種(啞口仙女蟲和鉤蝦屬),其余4種均為昆蟲綱。鉤蝦屬、四節蜉屬是不同時期的共同優勢種,也是5條河流的廣布種。
整體上看,Shannon-Wiener 多樣性指數(圖4-a)、Pielou均勻度指數(圖4-c)和Simpson多樣性指數(圖4-d)在每條河流呈現出較為一致的時間變化趨勢,而Margalef豐富度指數(圖4-b)在3個時期無明顯變化。除哈爾蓋河外,其他河流的生物多樣性幾乎都呈現從洄游前期、中期到后期逐漸降低的趨勢,哈爾蓋河的生物多樣性指數變化趨勢相反。
2.4? ?底棲動物與環境因子的相關性
青海湖入湖河流各采樣斷面的環境特征差異較大。5條河流上游斷面的底質中含有較多石塊和卵石,生境異質性高,受干擾程度小;中下游底質類型逐漸過渡為以礫石和細沙為主,且下游河口段河岸帶固化嚴重(表2)。上游斷面水體溶氧(DO)總體高于河口段,而電導率(Cond)明顯低于下游斷面。但總氮(TN)、總磷(TP)、氨氮(NH4-N)含量并無明顯差異(表3)。
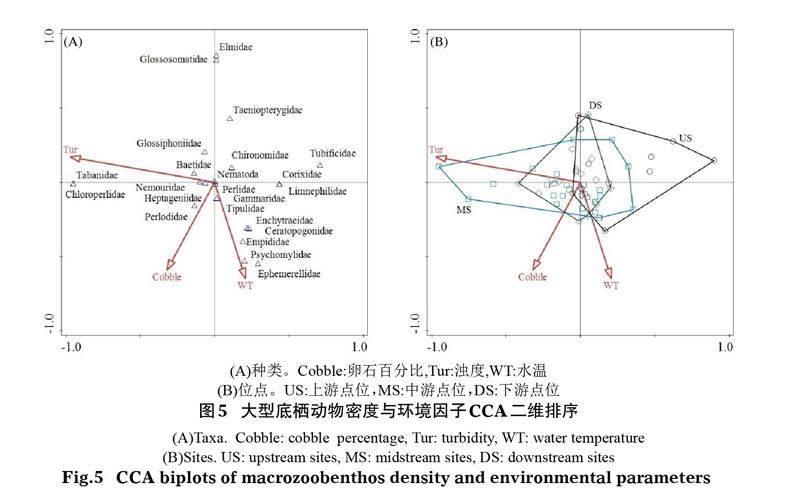
典型對應分析的前選法和蒙特卡羅轉置法分析表明,影響青海湖入湖河流大型底棲動物密度的關鍵環境因子有水溫(WT)、卵石百分比(Cobble)和濁度(Tur)(P<0.05),第一軸和第二軸的特征值分別為0.192和0.166。從CCA二維排序圖5可以看出,水溫和卵石百分比主要貢獻于第二軸,濁度主要貢獻于第一軸,其中水溫和卵石百分比與第二軸呈負相關關系,濁度與第一軸呈負相關關系。分析環境因子對大型底棲動物空間分布的影響,中游和下游底棲動物密度的空間分布與濁度密切相關,底棲動物種類主要包括舌蛭科(Glossiphoniidae)、虻科(Tabanidae)、綠襀科(Chloroperlidae)和顫蚓科(Tubificidae);上游斷面則與卵石百分比和水溫關系更密切,底棲動物種類主要包括小蜉科(Ephemerellidae)、舞虻科(Empididae)、管石蛾科(Psychomylidae)、舌石蛾科(Glossosomatidae)和溪泥甲科(Elmidae)。
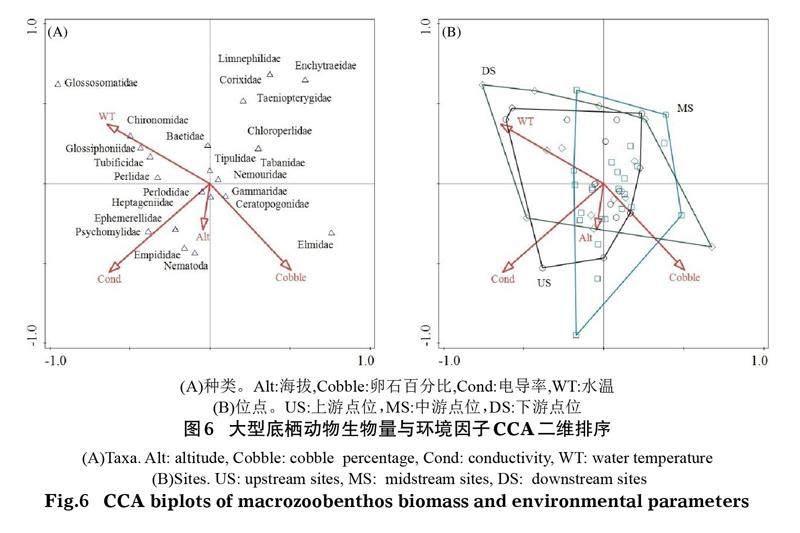
相同方法分析還表明,影響青海湖入湖河流大型底棲動物生物量的關鍵環境因子有電導率(Cond)、海拔(Alt)、水溫(WT)和卵石百分比(Cobble)(P<0.05)(圖6)。第一軸和第二軸的特征值分別為0.401和0.324。從CCA二維排序圖可以看出,卵石百分比、水溫和海拔主要貢獻于第二軸,電導率主要貢獻于第一軸;其中卵石百分比和海拔與第二軸呈負相關關系,水溫與第二軸呈正相關關系,電導率與第一軸呈正相關關系。從環境因子對大型底棲動物空間分布的影響看,上游大型底棲動物生物量的空間分布與水溫和卵石百分比密切相關,底棲動物種類主要包括四節蜉科(Baetidae)、舌石蛾科和搖蚊科(Chironomidae);中游底棲動物生物量的空間分布與海拔密切相關,底棲動物種類主要包括沼石蛾科(Limnephilidae)、劃蝽科(Corixidae)和線蚓科(Enchytraeidae);下游大型底棲動物生物量的空間分布與電導率密切相關,底棲動物種類主要包括小蜉科(Ephemerellidae)、舞虻科(Empididae)和蠓科(Ceratopogonidae)。
2.5? ?大型底棲動物的漁產潛力
根據漁產潛力經驗公式估算,黑馬河、布哈河、泉吉河、沙柳河、哈爾蓋河大型底棲動物的年均漁產潛力差別很大。以布哈河最高,其次是泉吉河和沙柳河(表4)。在這些河流中,鉤蝦對漁產潛力的貢獻比均最高,5條主要河流的大型底棲動物載魚量也差別很大。
3? ?討論
3.1? ?高海拔河流大型底棲動物的分布特點
黑馬河、布哈河、泉吉河、沙柳河和哈爾蓋河地處青藏高原東北部,屬典型的高原河流,具有海拔高、水溫低、日照時間長、水熱同期、降水量年內分布不均、河道落差大、微生境異質性高和有機碎屑降解慢等特點(吳瑞琛,1990;崔步禮等,2011)。高海拔造成的水溫、光照、地形等差異會塑造多樣化的大型底棲動物群落(Tomanova et al, 2007)。高海拔地區,河流水溫低,有機質降解慢,大型底棲動物食物來源多為粗顆粒有機碎屑,因此冷水性且功能攝食類群為撕食者的種類占優勢(王昱等,2021),如本研究中的優勢種鉤蝦屬;另外,海拔高、光照強、河水濁度低、著生藻生長較快,加之河道落差較大、水流較急,一些行攀援或粘附生活的刮食者也是大型底棲動物主要組成,如身體扁平的微動蜉屬和粘附基質生活的直突搖蚊屬。高原河流的海拔落差較大,水流對河床底質的沖刷、搬運作用強烈,且河床底質多為卵石和大石塊,因而生境異質性較高,生物多樣性也相對較高,底棲動物主要為節肢動物(簡東等,2015),如雅魯藏布江流域大型底棲動物的種類組成中水生昆蟲占比超過74%(Xu et al,2018)。本研究中,蜉蝣目、襀翅目、毛翅目、鞘翅目和雙翅目等水生昆蟲種類數占比大于80%,這也與其他高海拔河流大型底棲動物的相關研究一致(Mishra et al, 2013;趙娜等,2020)。
3.2? ?河流底棲動物漁產潛力與漁業資源保護
近年來,青海湖裸鯉資源量銳減,2004年青海湖裸鯉被列入《中國物種紅色名錄》瀕危物種名單。魚卵和仔魚是魚類資源補充和可持續利用的基礎(武云飛,1992)。黑馬河、布哈河、泉吉河、沙柳河和哈爾蓋河等入湖河流是親魚產卵的重要通道,調查發現這些河流各河段均有產卵場存在(付生云等,2019)。攝食是影響魚類早期資源生長發育、存活與死亡的重要因素(劉熠,2020),本次魚類調查統計得出泉吉河與沙柳河洄游親魚總量分別為176萬尾和121萬尾左右,大型底棲動物的漁產潛力估算值折合能供給親魚量分別為12萬尾和11萬尾,分別占洄游親魚總量的6.8%和9.1%,是親魚重要的能量補給站。本次調查雖然未估算黑馬河、布哈河和哈爾蓋河的洄游親魚量,但從湖區青海湖裸鯉密度空間分布得知,布哈河入湖河口區域的密度最高,產卵場也最多,因而本研究估算出大型底棲動物漁產潛力空間分布最大值出現在布哈河是較為合理的。
青海湖裸鯉人工增殖放流工作于2002年開始,截至2020年,其資源量增加了38倍,其中增殖放流貢獻占比約為23%。要想實現漁業的可持續發展,除增殖放流外,還需要合理利用漁產潛力(洪榮華等,2005),保護主要入湖河流水域生態系統的生物多樣性和生態完整性,可為青海湖裸鯉完成世代繁衍提供優渥的生態環境。
參考文獻
陳民琦,林建國,應百才,1990. 青海湖封湖3年對裸鯉種群結構的影響初探[J]. 青海大學學報(自然科學版), (1):50-56.
崔步禮,李小雁,李岳坦,等,2011. 青海湖流域河川徑流特征及其對降水的滯后效應[J]. 中國沙漠, 31(1):247-253.
崔奕波,2005. 長江流域湖泊的漁業資源與環境保護[M]. 北京:科學出版社:304-307.
崔永德, 2008. 云南湖泊寡毛類環節動物研究[D]. 北京:中國科學院研究生院.
馮宗煒,馮兆忠,2004. 青海湖流域主要生態環境問題及防治對策[J]. 生態環境學報, 13(4):467-469.
付生云,周衛國,張濤,等,2019. 青海湖裸鯉洄游排卵、受精高峰期的繁殖特征[J]. 畜牧獸醫科技信息, (11):35-36.
國家環保局,1989. 水和廢水監測分析方法[M]. 3版. 北京:中國環境科學出版社.
何雪寶, 2011. 西藏和四條大型河流水棲寡毛類區系研究[D]. 北京:中國科學院研究生院.
洪榮華,劉其根,陳來生,2005. 千島湖的漁產潛力估算及可持續發展途徑[J]. 水產科技情報, 32(4):175-177.
簡東,黃道明,常秀嶺,等,2015. 拉薩河中下游底棲動物群落結構特征分析[J]. 水生態學雜志, 36(1):40-46.
蔣小明,2012. 中西部河流大型無脊椎動物多樣性格局及其環境評價[D]. 北京:中國科學院研究生院.
李岳坦,李小雁,崔步禮,等,2010. 青海湖流域50年來(1956-2007年)河川徑流量變化趨勢——以布哈河和沙柳河為例[J]. 湖泊科學, 22(5):757-766.
劉熠,2020. 長江下游刀鱭(Coilia nasus)仔稚魚的時空分布及其食性差異研究[D]. 上海:上海海洋大學.
劉月英,張文珍,王躍先,等,1979. 中國經濟動物志:淡水軟體動物[M]. 北京:科學出版社.
孟星亮,何玉邦,宋卓彥,等,2014. 青海湖區大型底棲動物群落結構與空間分布格局[J]. 水生生物學報, 38(5):819-827.
青海省生物研究所,1975. 青海湖地區的魚類區系和青海湖裸鯉的生物學[M]. 北京:科學出版社:37-45.
時盛博,戴虎德,李萬志,等,2017. 青海省海北州水分資源變化[J]. 中國農學通報, 33(35):119-127.
史建全,祁洪芳,楊建新,2004. 青海湖自然概況及漁業資源現狀[J]. 淡水漁業, 26(5):3-5.
史建全,祁洪芳,楊建新,等, 2000. 青海湖裸鯉資源評析[J]. 淡水漁業, 30(11):38-40.
唐紅渠,2006. 中國搖蚊科幼蟲生物系統學研究[D]. 天津:南開大學.
汪洋,2019. 青海湖裸鯉仔稚魚工廠化培育中投喂搖蚊幼蟲對生長的影響[J]. 科學養魚, (3):71-72.
王洪鑄,2002. 中國小蚓類研究——附中國南極長城站附近地區兩新種 [M]. 北京:高等教育出版社.
王昱,左一鋒,馮起,等,2021. 黑河中上游大型底棲動物棲息地適宜度評估[J]. 生態學雜志, 40(4):1116-1127.
吳瑞琛,1990. 西藏高原水文特性[J]. 水文, (2):53-56.
武云飛,1992. 青藏高原魚類[M]. 成都:四川科學技術出版社.
閆云君,1998. 淺水湖泊大型底棲動物生態能量學及生產量的研究[D] . 武漢:中國科學院水生生物研究所.
楊洪志,王基琳,1997. 青海湖底棲生物及其生產力分析[J]. 青海科技, 4(3):36-39.
姚維志,史建全,祁洪芳,等,2011. 2006-2010年夏季青海湖浮游植物研究[J]. 淡水漁業, 41(3):22-28.
張信,熊飛,唐紅玉,等,2005. 青海湖裸鯉繁殖生物學研究[J]. 漁業科學進展, 26(3):61-67.
趙娜,周罕覓,李志威,等,2020. 黃河源典型卵石河床底棲動物的小尺度分布[J]. 水生態學雜志, 41(1):22-29.
中國河湖大典編纂委員會,2014. 中國河湖大典:西北諸河卷[M]. 北京:中國水利水電出版社.
中國科學院西北高原生物研究所,1989. 青海經濟動物志:魚篇[M]. 西寧:青海人民出版社.
周長發,蘇翠榮,歸鴻,2015. 中國蜉蝣概述[M]. 北京:科學出版社.
Barbour M T, Gerritsen J, Snyder B D, et al, 1999. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish[M]. Second Edition, Washington, USEPA DC.
Li X Y, Xu H Y, Sun Y L, et al, 2007. Lake-Level Change and Water Balance Analysis at Lake Qinghai, West China during Recent Decades[J]. Water Resources Management, 21(9):1505-1516.
Margalef R, 1968. Perspective in Ecological Theory[M]. Chicago: University of Chicago Press.
Mishra A S, Nautiyal P, Semwal P, 2013. Distributional patterns of benthic macro-invertebrate fauna in the glacier fed rivers of Indian Himalaya[J]. Our Nature, 11(1):36-44.
Morse J C, Yang L, Tian L, 1994. Aquatic Insects of China Useful for Monitoring Water Quality[M]. Nanjing: Hohai University Press.
Pielou E C, 1966. The Measurement of Diversity in Different Types of Biological Collections[J]. Journal of Theoretical Biology, 13(1):131-144.
Shannon C E, Weaver W, 1949. Mathematical Theory of Communication[M].? Illinois:University of Illinois Press.
Tomanova S, Tedesco P A, Campero M, et al, 2007. Longitudinal and altitudinal changes of macroinvertebrate functional feeding groups in neotropical streams: a test of the Stream Continuum Concept[J]. Fundamental and Applied Limnology, 170:233-241.
Xu M Z, Zhao N, Zhou X D, et al, 2018. Macroinvertebrate Biodiversity Trends and Habitat Relationships within Headwater Rivers of the Qinghai-Tibet Plateau[J]. Water, 10(9):1214.
(責任編輯? ?萬月華)
Tempora-spatial Distribution and Potential Fish Production Capacity
of Macrozoobenthos in the Main Inflow Rivers of Qinghai Lake
XIE Ting‐feng1,2, HE Ya‐jing2,3, ZHAO Yong‐jing2, XU Dong‐po4, YU Lu‐xian5,
WANG Hong‐zhu2, CUI Yong‐de2
(1. Tibet University, Lhasa 850000, P.R. China;
2. State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology,
Chinese Academy of Sciences, Wuhan 430072, P.R. China;
3. University of Chinese Academy of Sciences, Beijing 100049, P.R. China;
4. Freshwater Fisheries Research Center of Chinese Academy of Fishery Sciences, Wuxi 214081, P.R. China;
5. Qinghai Key Laboratory of Qinghai-Lake Naked Carps Breeding and Conservation,
Rescue and Rehabilitation Center of Naked Carps of Qinghai Lake, Xining 810016, P.R. China)
Abstract:In this study, the five primary inflow rivers to Qinghai Lake (Heima River, Buha River, Quanji River, Shaliu River, Ha'ergai River) were investigated. The temporal-spatial distribution characteristics of the macrozoobenthos community in the five rivers were characterized, and potential fishery production was then estimated based on the macrozoobenthos stock. The aim was to provide scientific guidance for resource enhancement and the sustainable recovery of Gymnocypris przewalskii in Qinghai Lake. In 2019, during the early, middle and late migration period of G. przewalskii, the microzoobenthos and environmental parameters were investigated at 18 sampling transects in the upper, middle and lower reaches of the five rivers. A total of 101 microzoobenthos taxa (species/genus) belonging to 44 families, 9 orders, 7 classes and 5 phyla were observed during the investigation, with absolute dominance by aquatic insects (86 species/genus, accounting for 85.1% of the total species). Spatially, species richness was highest in Heima River (56 species/genus) and lowest in Ha'ergai River (27 species/genus). The taxa number of EPT (Ephemeroptera, Plecoptera and Trichoptera) decreased from upstream to downstream. Microzoobenthos density and biomass were both highest in Quanji River (544 ind/m2, 4.77 g/m2) and lowest in Haergai River (85 ind/m2, 0.31 g/m2), while the Shannon-Wiener and Simpson diversity, Pielou evenness and Margalef richness indices were highest in Heima River. Temporally, total taxa number and biodiversity indices of microzoobenthos decreased with each of the three successive migration periods, while the density and biomass increased. The primary environmental parameters influencing the distribution of macrozoobenthos density and biomass were substrate, altitude, water temperature, turbidity, and conductivity. The estimated potential fishery production capacity, based on macrozoobenthos, was highest in Buha River(285 129 kg), followed by Quanji River(20 694 kg) and Shaliu River(18 857 kg), and consistent with the distribution of G. przewalskii density in the different spawning grounds. Our findings indicate that macrozoobenthos can provide an adequate source of food for parent G. przewalskii during migration and spawning.
Key words:zoobenthos; temporal-spatial distribution; potential fishery production; Gymnocypris przewalskii
收稿日期:2021-08-15? ? ? 修回日期:2022-11-09
基金項目:農業農村部財政專項“西北地區重點水域漁業資源與環境調查”。
作者簡介:謝廷鋒,1994年生,男,碩士研究生,研究方向為底棲動物生態學與濕地生態系統生態學。E-mail:xietf@ihb.ac.cn
通信作者:崔永德,1978年生,男,副研究員,主要從事寡毛類分類學與生態學、底棲動物生態學及生態系統健康評價研究。E-mail:ydcui@ihb.ac.cn
