秸稈高值利用合成中鏈脂肪酸研究進展
2023-05-17 08:54:14安柯萌趙立欣姚宗路于佳動李再興黃亞麗梁依申瑞霞
中國農業科技導報 2023年3期
安柯萌, 趙立欣, 姚宗路, 于佳動*, 李再興,黃亞麗, 梁依, 申瑞霞
(1.河北科技大學環境科學與工程學院,石家莊 050018;2.中國農業科學院農業環境與可持續發展研究所,農業農村部華北平原農業綠色低碳重點實驗室,北京 100081)
中國是農業大國,糧食、蔬菜、油料等作物收獲產生大量農作物秸稈。近5年全國主要農作物秸稈產生量在8億t左右,可收集資源量約7億t,2021年秸稈綜合利用率為88.1%[1-2]。秸稈含有豐富的有機物,厭氧發酵產沼氣和熱解炭氣聯產是秸稈能源化利用的2個重要技術手段,秸稈降解率可達到40%~60%,副產物還可用于生產有機肥[3]。但從能源及有機肥供需關系分析,目前農業廢棄物資源化利用全鏈條產業體系還不健全,收集運輸和儲存有一定難度,產品利用價值不高,阻礙了沼氣工程和熱解工程的發展。因此,基于秸稈沼氣和熱解工程功能,提升秸稈資源化處理產業鏈、價值鏈,遵循生物和熱化學轉化技術原理,尋找一種中間產物高值轉化途徑,生產高附加值產品,可助力產業健康發展。
近年來,一種生物合成中鏈脂肪酸(mediumchain fatty acids,MCFAs)技術成為有機廢棄物高值化利用的研究熱點,MCFAs在農業領域用作飼料添加劑、鹽堿土壤改良劑、生物農藥等,售價10 000~20 000元·t-1,是沼氣/有機肥價格的3~5倍。秸稈生物轉化和熱化學轉化過程的重要中間產物均可用于合成MCFAs,一方面,厭氧發酵水解酸化產物乙酸、丁酸、乳酸、乙醇等作為電子受體、電子供體,在功能微生物作用下通過碳鏈延長(chain elongation,CE)途徑合成MCFAs;另一方面,秸稈熱解炭氣聯產的合成氣富含CO2、H2和CO,通入厭氧發酵體系在功能微生物的作用下轉化為乙酸和乙醇,進行CE合成MCFAs。
在有機廢棄物厭氧發酵中間產物合成MCFAs方面,德國圖賓根大學、比利時根特大學及中國的復旦大學、哈爾濱工業大學等,利用污水和餐廚垃圾等原料合成MCFAs,選育了高效微生物菌種,研發了高效產酸調控工藝,己酸、辛酸產率為70%~80%[4-5];在秸稈熱解炭氣聯產合成氣轉化MCFAs方面,丹麥科技大學、北京化工大學等在以合成氣為底物的功能微生物馴化、抑制機理分析等方面做了較多研究,己酸、辛酸產率為30%~70%[6]。功能微生物挖掘、發酵機制探索、工藝優化等是理論研究與技術突破的主要關注點,但是以秸稈為主要原料的合成MCFAs特性及轉化機制研究較少。因此,本研究圍繞秸稈生物轉化和熱化學轉化技術,重點關注秸稈厭氧發酵中間產物(乙酸、乙醇等)以及熱解合成氣(CO2、H2、CO)在微生物作用下轉化合成MCFAs的研究進展,分析2條路徑轉化合成MCFAs的原理、功能微生物、工藝調控及強化等的研究進展,提出了秸稈合成MCFAs主流應用工藝,為秸稈綜合利用工作提供參考。
1 秸稈轉化合成MCFAs原理
秸稈轉化合成MCFAs主要分為2個階段,包括秸稈轉化成電子供體(乳酸、乙醇)/電子受體(乙酸、丙酸、丁酸等)階段和CE階段,其中,CE階段每輪反應實現2個碳原子個數的增加(圖1)。
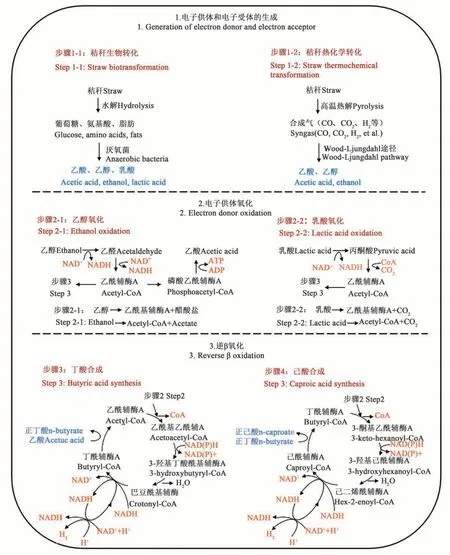
圖1 秸稈轉化合成MCFAs代謝過程Fig. 1 Metabolic process of MCFAs synthesis by straw transformation
秸稈可通過生物轉化(步驟1-1)和熱化學轉化(步驟1-2)生成電子供體和電子受體。秸稈生物轉化途徑主要是在厭氧條件下進行水解酸化產生乙酸、乳酸、乙醇等,其中參與秸稈水解酸化的酶主要有纖維素酶、胞外水解酶(如淀粉酶、蛋白酶和纖維素酶)、乙酰輔酶A、丁酰輔酶A等。秸稈熱化學轉化途徑是在700~800 ℃下熱解,產生混合熱解氣(30%~40% CO、30%~40% CO2、15%~20% CH4和5%~10% H2),氣體混合物經過濾去除焦油和固體顆粒等后,清潔氣體在厭氧微生物作用下通過 Wood-Ljungdahl途徑,將 CO、CO2和 H2催化轉化為乙酰輔酶A,并進一步轉化為乙酸和乙醇,其中參與Wood-Ljungdahl途徑的關鍵酶有甲酸脫氫酶、CO脫氫酶、乙酰輔酶A、乙醛脫氫酶和乙醇脫氫酶等。
CE反應是MCFAs生產的關鍵代謝路徑,核心是逆β氧化,包括電子供體氧化和電子受體的還原[7]。乙醇和乳酸作為電子供體其氧化產物分別為乙酰基輔酶A、醋酸鹽(步驟2-1)和乙酰基輔酶A、CO2(步驟2-2)。電子供體氧化后的乙酰輔酶A進入逆β氧化途徑,經過逆β氧化周期(乙酸增長為丁酸,步驟3),最終生成己酸(步驟4)。在己酸累積并且有乙醇/乳酸存在的條件下,可能會通過逆β氧化途徑向羧酸鏈中額外添加2個碳原子,將C6(己酸)延長至C8(辛酸)。隨著碳鏈(最多至C12)的延長,產物的毒性會隨之增加,對微生物群落也會產生一定的抑制作用[8]。因此,如果產物在反應過程中不及時提取,逆β氧化途徑就會成為CE的限制過程。
2 功能微生物
秸稈在功能微生物的作用下,經水解酸化或合成氣發酵轉化為乙醇、乳酸、乙酸、丁酸等,為合成MCFAs微生物提供可直接利用的底物。秸稈水解酸化階段的功能微生物主要有Clostridium_sensu_stricto、Bifidobacterium、Rummeliibacillus和Lactobacillus等菌屬[9-10]。功能微生物又分為純菌體系和復合菌體系,純菌培養試驗表明Clostridium autoethanogenum、Clostridium ljungdahlii和Clostridium ragsdalei等均能利用CO2和H2為底物產生乙醇。復合菌培養試驗證明,Clostridium和Bifidobacterium可以在微曝氣條件下協同作用生產乙酸和乳酸[11]。近年來,利用合成氣轉化為乙醇和乙酸過程中功能微生物的挖掘成為研究熱點,Struber等[12]和Tanner等[13]在中溫發酵體系下發現了Clostridium aceticum、Acetobacteriumwoodii、Clostridiumljungdahlii、Clostridium autoethanogenum等功能微生物,其中Clostridium ljungdahlii是合成氣發酵的功能菌株,研究證實該菌株通過Wood-Ljungdahl途徑利用生物酶將CO、CO2和H2催化轉化為乙酰輔酶A,進而轉化為乙醇和乙酸。但在目前的研究中,合成氣轉化乙酸、乙醇的產量較低,仍處于實驗室研究階段。
微生物利用電子供體和電子受體為底物,通過CE反應合成MCFAs。已報道的CE功能菌株包括Clostridium kluyveri、Ruminococcaceae bacterium、Clostridiumsp.BS-1和Megasphaera elsdenii,其中被廣泛應用的Clostridium kluyveri能夠利用乙酸、乙醇作為底物合成丁酸并進一步合成己酸,在連續高產MCFAs反應器中的豐度可達50%~70%[14]。Megasphaera elsdenii和Ruminococcaceae strainCPB6被證實是以乳酸為底物的CE細菌,研究發現了編碼催化CE反應酶復合物的完整基因組[15]。Zhu等[16]通過純菌培養試驗證實了Ruminococcaceae bacteriumCPB6可利用乳酸作為底物高效合成己酸,在乳酸濃度為45.1 g·L-1條件下,己酸產量達16.6 g·L-1。復合菌種也表現出合成MCFAs的優勢,朱孔云等[9]以秸稈為原料,發現在混菌厭氧發酵鏈延長體系中,Clostridia相對豐度達到40.3%,Clostridia菌群能夠將各種碳水化合物轉化為己酸、辛酸等。近年來,隨著生物信息學和厭氧微生物分離純化研究方法的不斷進步,亥姆霍茲環境研究中心和康斯坦茨大學分離出新型梭狀芽胞桿菌物種,與Clostridium kluyveri和Clostridium luticellarii具有極高的相似性,能夠將乳酸轉化為己酸,進一步豐富了MCFAs合成菌種資源[17](表1)。
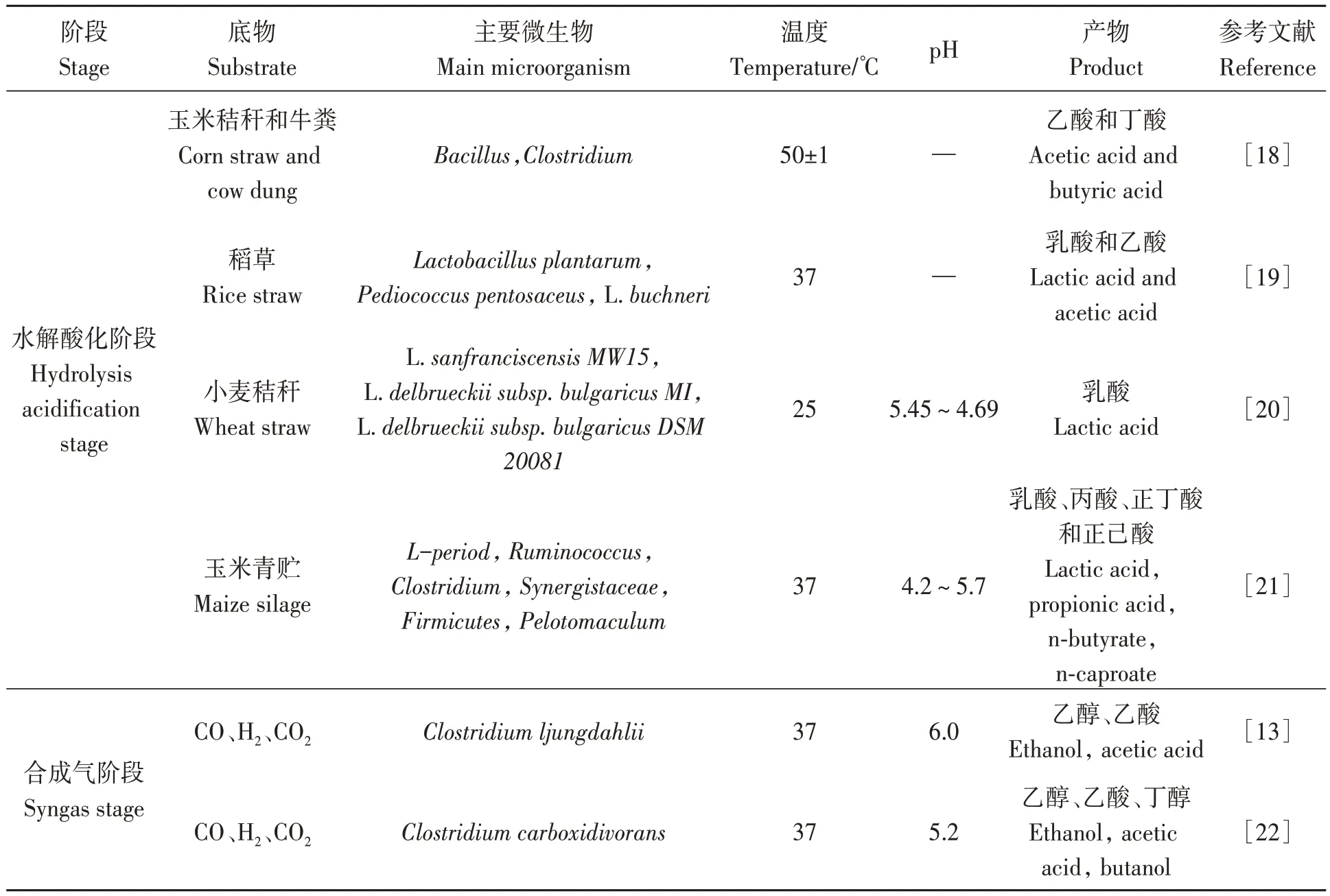
表1 秸稈轉化為MCFAs微生物工藝研究進展Table 1 Research progress on microbial technology of straw conversion to MCFAs
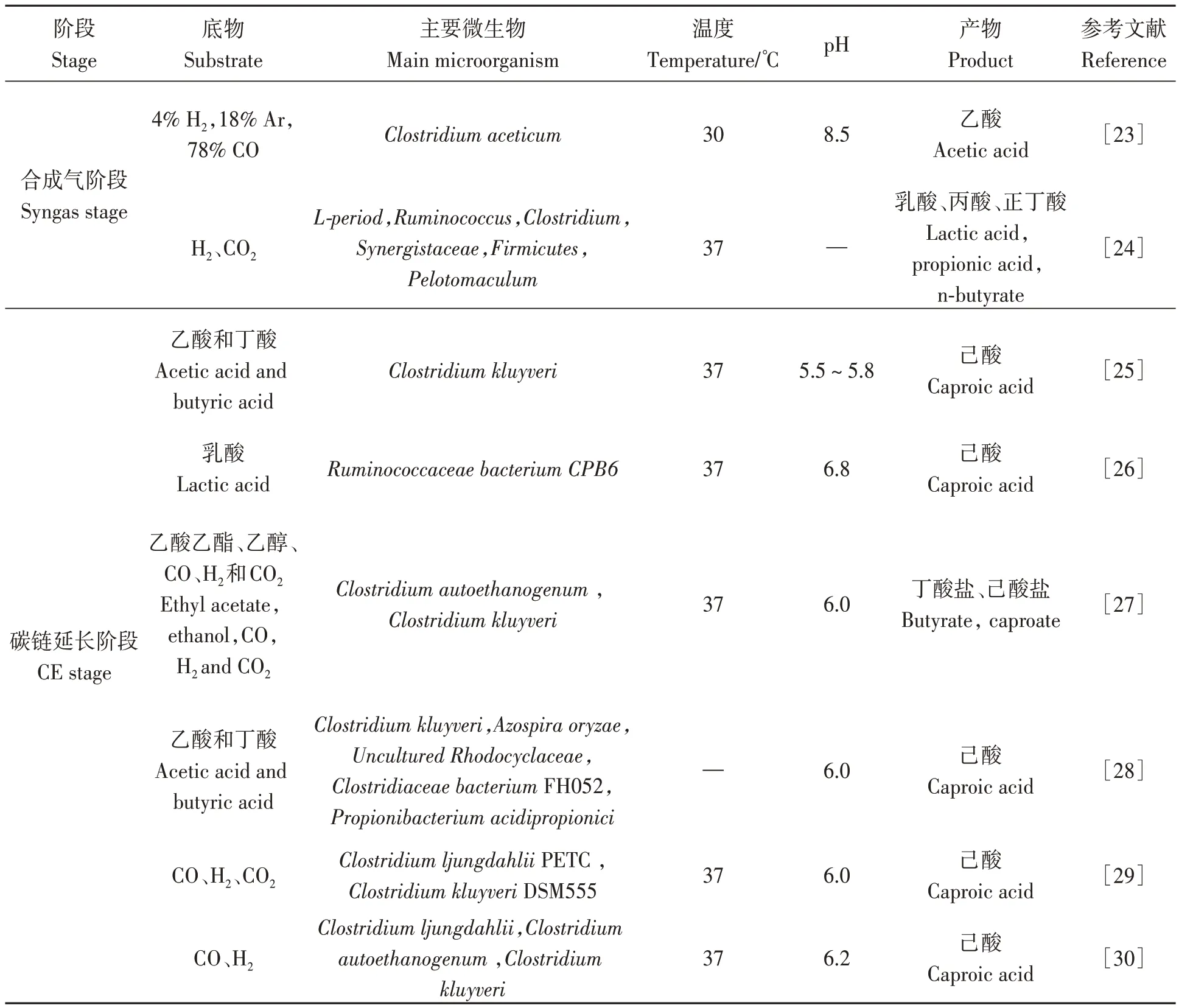
表1 秸稈轉化為MCFAs微生物工藝研究進展Table 1 Research progress on microbial technology of straw conversion to MCFAs 續表Continued
3 秸稈轉化生成電子供體和電子受體
3.1 秸稈水解酸化生成電子供體和電子受體
秸稈水解酸化為MCFAs合成提供電子供體和電子受體。秸稈中富含纖維素、半纖維素、木質素等難降解組分,導致水解酸化效率較低,可通過預處理的方式破壞木質纖維素結構、提高秸稈的降解效率,常用蒸汽爆破、柔絲處理或加入酸堿試劑等單一或復合手段[31-32]。秸稈自然水解酸化產物以乙酸、丙酸、丁酸為主,乙醇、乳酸占比較低(3.2%~21.4%),研究人員探索了水解酸化階段的調控工藝,如在秸稈水解酸化不同階段添加復合菌群或調控體系pH、溫度等關鍵參數來優化酸化產物組分,通過優化關鍵運行參數使酸化產物向乙醇型或乳酸型方向驅動,顯著提高了電子供體(乙醇、乳酸)、電子受體(乙酸、丁酸)協同生產效率,乙醇、乳酸占比提高到40%~55%[11,20]。研究發現[33],秸稈發酵體系的CE更傾向于利用乙醇和乙酸作為底物來生產MCFAs,因此通過對秸稈水解酸化體系定向調控促進產酸,可以為CE體系提供充足電子供體和電子受體(表2)。
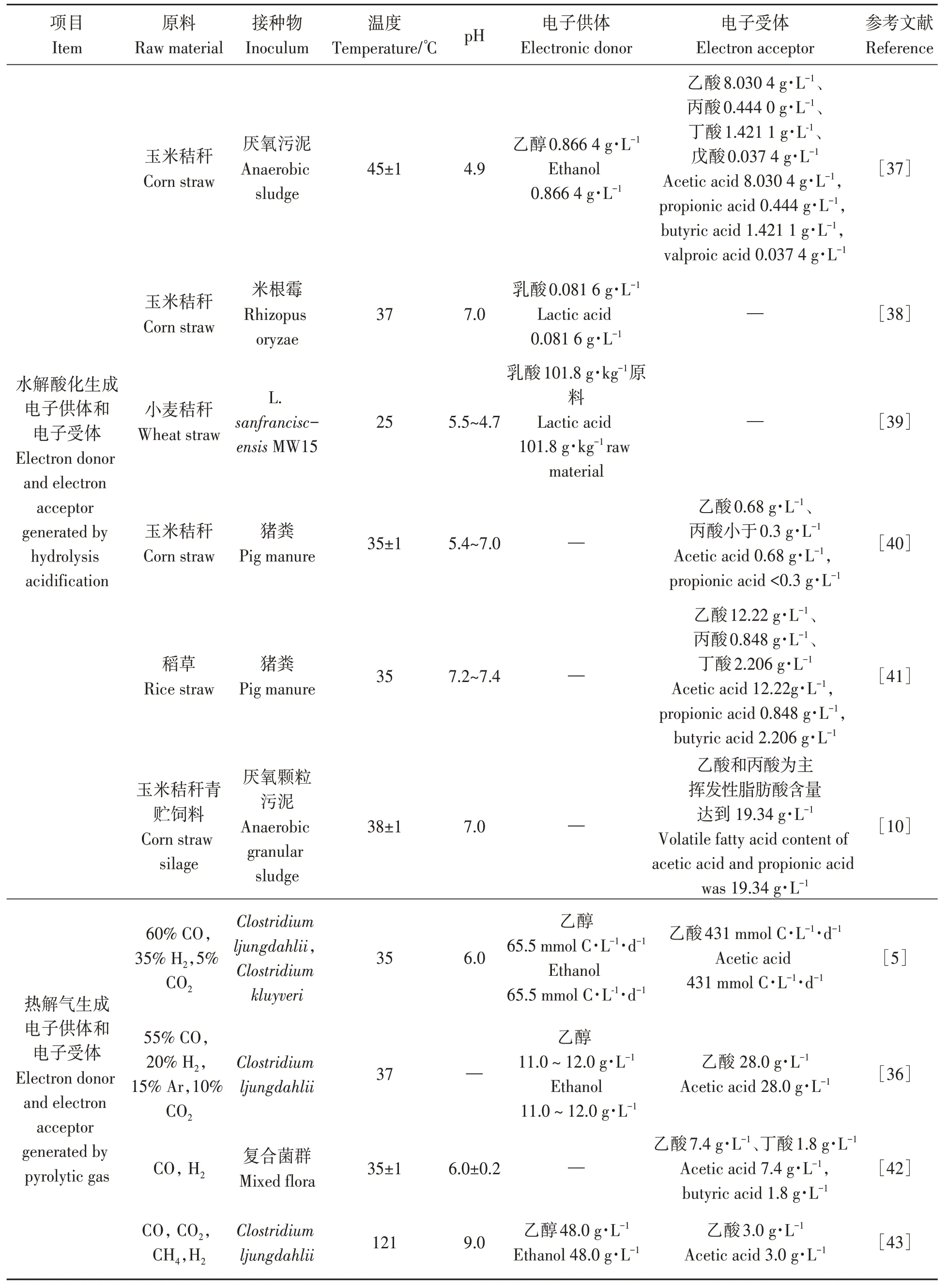
表2 生物轉化和熱化學轉化為電子供體和電子受體的研究進展Table 2 Advances in bioconversion and thermochemical conversion to electron donor and electron acceptor
3.2 秸稈熱解氣生成電子供體和電子受體
秸稈熱解生成合成氣(CO、H2、CO2)是電子供體和電子受體再生的關鍵步驟,發酵過程中合成氣組分決定了電子供體和電子受體的含量水平。秸稈熱解過程以空氣為氣化劑時,產物合成氣中含有體積分數15%~22%的CO、8%~12%的H2、10%~15%的CO2以及50%左右的N2[34]。原料含水率也是影響合成氣組分的重要因素,于杰等[35]研究發現,隨著秸稈含水率增大(5%~25%),合成氣中的CO2與H2含量逐漸升高,CO含量下降。發酵過程中的總壓力可影響發酵微生物活性,進而對電子供體和電子受體的含量水平產生影響。Najafpour等[36]在 81~182 kPa 下將 CO(55%)、CO2(10%)、H2(20%)和 Ar(15%)通入反應器進行批次試驗,發現在Clostridium ljungdahlii作用下,乙醇含量隨氣壓的增加而增加,在182 kPa時乙醇含量最高,達12 mg·L-1,乙酸含量達2~4 mg·L-1,在相同的工藝條件下進行連續發酵,會獲得更高的乙醇和乙酸含量(12~15 mg·L-1)。目前,合成氣發酵體系還需在發酵過程穩定性控制、提高產醇產酸效率等方面加強攻關。
4 CE調控強化工藝研究
4.1 CE產MCFAs調控技術
醇酸比、pH、溫度、水力停留時間等是影響CE合成MCFAs效率的主要因素,調控不同因素對厭氧發酵產MCFAs的影響見表3。
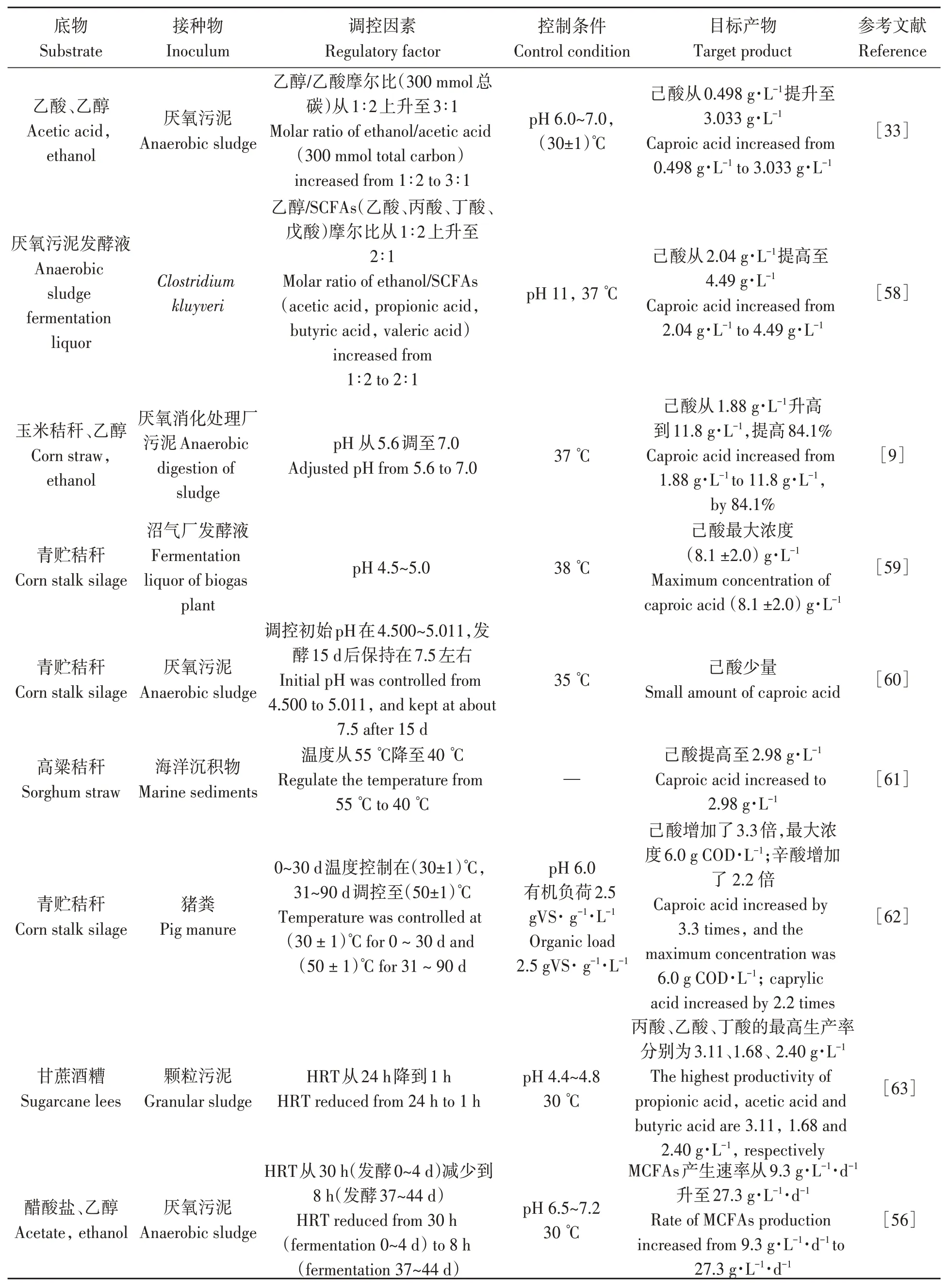
表3 生產MCFAs調控工藝研究進展Table 3 Research progress in regulation process of MCFAs production
4.1.1 醇酸比 電子供體和電子受體的含量水平和比例會對MCFAs生成效率產生影響。研究表明[44-45],醇酸比過低或過高均不利于MCFAs的合成,醇酸比較低時可能缺少CE所需的電子供體,但過高的醇酸比會對MCFAs合成產生毒害作用。Liu等[33]研究了不同的醇酸比對CE體系MCFAs的產量和組分的影響,在乙醇/乙酸的摩爾比大于2∶1(300 mmol總碳)時,能夠為CE反應提供足夠的電子供體,有利于MCFAs的產生,當乙醇/乙酸的摩爾比從3∶1(乙醇濃度(226.63±2.52) mmol·L-1)增加到5∶1(乙醇濃度(226.63±2.52)mmol·L-1)時,MCFAs的產生量增加38%[46],醇酸比大于5∶1時,MCFAs產量開始下降。有研究利用Clostridium kluyveri strain3231B 和Clostridium kluyveri strainK.l的純培養微生物,對比了乙醇/乙酸摩爾比0.78~12.00的產酸性能,結果顯示乙醇/乙酸比例系數越大,其產物己酸含量越高[39,47]。合理設計電子供體和電子受體的含量水平和比例是工藝調控的重要內容。
4.1.2 pH pH可對CE功能微生物細胞內酶的活性和代謝途徑產生影響。大多產己酸菌最適pH在4~7,例如以青貯秸稈為底物的完全混合式反應器(continuous stirred tank reactor,CSTR)中pH保持在4.2~5.7,在Megasphaera elsdenii菌株作用下 ,己 酸 產 量 達 3.1 g·L-1[21],Ruminococcaceae bacteriumCPB6菌株在pH 5.0~6.5的酸性環境下,己酸最大產量達16.6 g·L-1[48]。目前研究表明,以乙酸為電子受體時,Clostridium kluyveri在pH 6.4~7.5時有較高的CE活性[49],并且添加碳酸氫鹽等無機碳源調控pH,己酸含量會提升。pH對混菌發酵過程中的微生物菌群具有直接選擇性,不同的微生物菌群最適pH范圍變化較大,如產甲烷菌的最適pH在6.5~7.5,而產酸菌pH在4.0~10.0可保持較高活性[50],在CE體系中保持pH 5.5左右可有效抑制產甲烷菌的活性,有利于己酸、辛酸等MCFAs的生成。
4.1.3 溫度 在CE體系中,發酵溫度直接影響CE功能微生物代謝產酸速率,有研究表明,在中溫條件(30~40 ℃)表現出更高的產MCFAs效率[41]。當系統溫度為(37±1) ℃時,產MCFAs效果最好,表明參與CE反應過程的丁酰輔酶A脫氫酶和鐵氧還蛋白氧化還原酶的活性較高,與己酸具有顯著的正相關性[51]。Zhu 等[16]研究了溫度(20、30、40、50 ℃)對Ruminococcaceae strainCPB6產己酸效能的影響,發現40 ℃是該菌種利用乳酸轉化己酸的最適溫度。Hollister等[52]比較了中溫(40 ℃)和高溫(55 ℃)體系產酸的差異,發現中溫和高溫下Clostridia、Bacill和Thermoanaerobacterium明顯富集,但中溫下產己酸菌的豐度更高,可獲得更高的己酸產量。此外有研究發現,在中低溫環境下(20~30 ℃)也有較高的MCFAs生產效率,低溫環境抑制部分產甲烷菌、硫化氫還原菌的功能,而CE功能菌群溫度適應范圍廣,在適宜的理化培養環境下易被富集[53-54]。
4.1.4 水力停留時間 調控水力停留時間(hydraulic retention time,HRT)會顯著影響酸化產物組分及含量,引起微生物豐度和數量的明顯差異,是連續運行MCFAs調控研究關注較多的影響因素之一。Grootscholten等[55]研究表明,當上流式厭氧反應器HRT為17 h時,連續發酵80 d,己酸產率達16.6 g·L-1·d-1。進一步研究通過縮短水力停留時間(提高向上流動的速度),產甲烷菌豐度及關鍵酶活性均下降30%以上,MCFAs的最大體積產率達到 57.4 g·L-1·d-1,MCFAs選擇性仍在80%以上[56],表明在較短的水力停留時間下,有利于CE功能菌的生長。Liu等[57]研究發現,HRT指示菌與不斷變化的環境和相應的功能之間存在很強的關聯性,縮短HRT,增加了HRT指示菌的豐度,而這部分指示菌90%以上被證明具有MCFAs合成功能,產甲烷菌豐度降至1%以下,HRT從8 d縮短到2 d,己酸、辛酸含量達到64.1、5.9 mmol C·L-1·d-1。在工藝調控上,可根據CE細菌生長速率快的特點,營造適宜的發酵環境,通過控制水力停留時間限制性培養,提高MCFAs產量。
4.2 CE產MCFAs強化技術
CE強化主要以理化強化和生物強化為主,理化強化是通過改善微生物發酵環境,達到MCFAs增產的目的。理化強化方法為熱沖擊強化、氣調強化、加入生物炭與微量元素等,例如,Clostridium kluyveri具有耐熱孢子,對熱沖擊具有一定的耐受力,對接種物進行熱沖擊處理可以相對富集MCFAs功能微生物,特別是在偏酸性或中性條件下連續發酵過程中抑制產甲烷菌的生長,減少底物競爭[64]。氣調強化通過調節氫分壓在3~10 kPa,可避免乙醇過度氧化,促使更多CE功能菌利用乙醇,一定程度上抑制了乙醇氧化菌、嗜氫產甲烷菌的生長[65]。添加生物炭、微量元素等,也可顯著提高MCFAs的生產效率。Liu等[66]研究發現,添加生物炭可通過吸附有機化合物、重金屬等提高MCFAs的轉化效率,粒徑小于5 μm的生物炭能夠強化微生物種間電子傳遞,促進丁酸向己酸的轉化,并能縮短CE反應周期,CE功能微生物明顯得到富集。微量元素在CE混合培養體系中,對微生物生長和代謝過程也表現出促進作用,Heike等[59]發現,在青貯秸稈發酵體系中,加入鐵元素可以促進丙酮酸鐵氧還蛋白氧化還原酶和梭菌氫化酶的活性,梭狀芽胞桿菌的豐度增加了55%~65%,MCFAs轉化效率提高了89%。
生物強化是通過添加功能微生物菌劑促進CE體系MCFAs的生產效率。CE過程中研究較多的微生物強化技術有加入CE菌劑[67]、培養馴化功能微生物[68]以及加入有助于微生物作用的酶等使CE過程加強。目前,研究報道的CE菌劑主要為具備生產MCFAs的純菌種和混合菌種,其中常見的純菌種見表1。另外,馴化厭氧消化污泥、土壤沉積物、產MCFAs反應器的污泥得到具有產MCFAs的混合菌種,通過加入甲烷抑制劑(溴乙烷磺酸鈉、氯仿等)或降低CE體系的pH至酸性(5.0~5.5)來抑制產甲烷菌的活性,從而使CE功能微生物成為混合體系的優勢菌種,馴化后的混合菌種產酸能力提高2~8倍。微生物強化相對于理化強化方法而言,具有能耗降低、二次污染減少等優勢,但CE轉化的穩定性仍需進一步提高[69]。
5 應用工藝
秸稈可通過生物轉化工藝和熱化學轉化工藝得到MCFAs,2種工藝下,適用條件、反應器和優化條件以及生產的MCFAs在農業和工業上的應用見圖2。
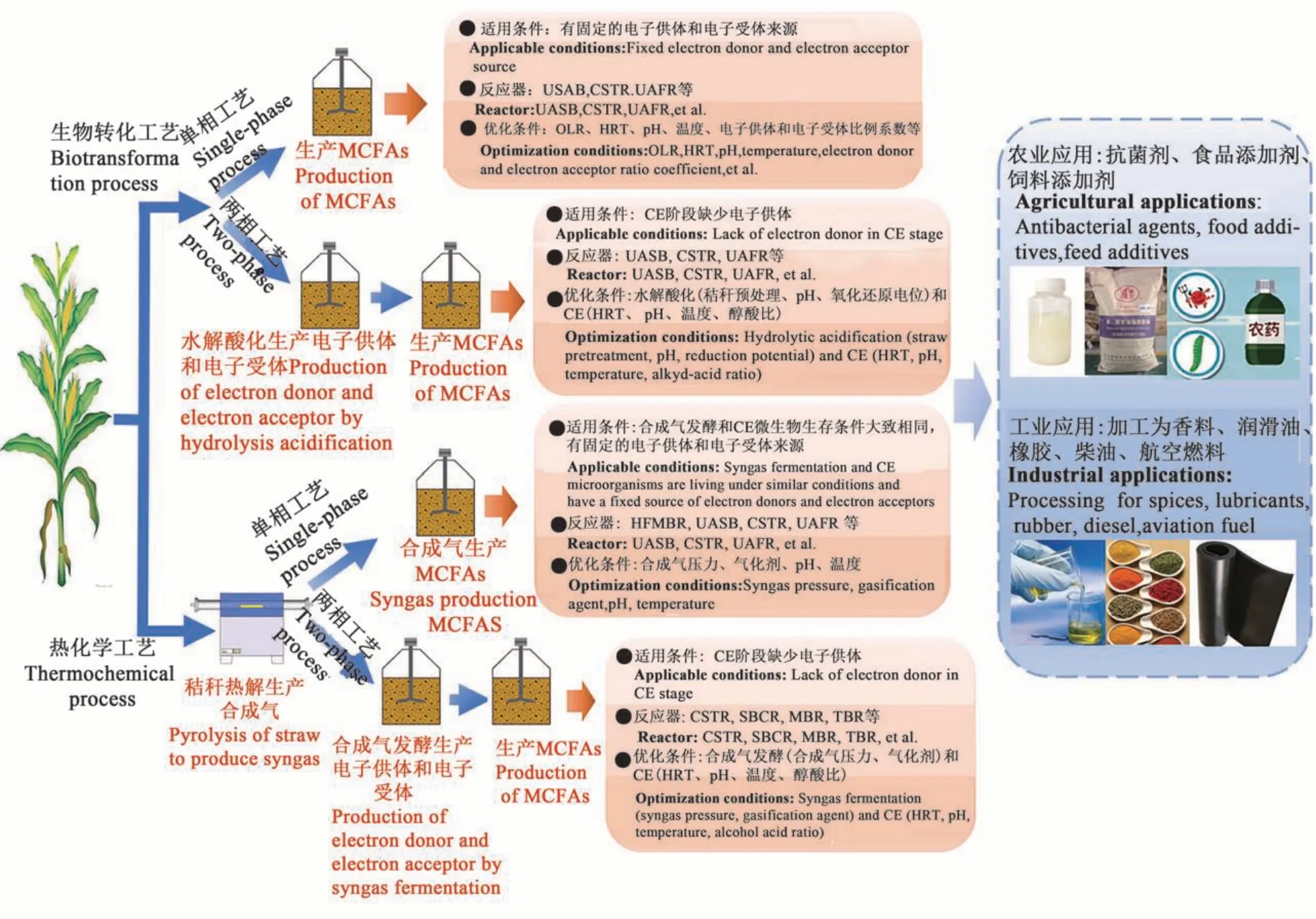
圖2 秸稈轉化合成MCFAs應用工藝Fig. 2 Application technology of straw transformation to synthesize MCFAs
5.1 生物轉化合成MCFAs工藝
生物轉化合成MCFAs可通過單相厭氧發酵工藝或兩相厭氧發酵工藝完成,常用的反應器有升流式厭氧污泥床(upflow anaerobic sludge bed,UASB)、完全混合式反應器(CSTR)、升流式厭氧過濾反應器(up-flow anaerobic sludge bed, UAFR )等,可根據秸稈發酵體系的電子供體和電子受體的量來選擇適用的發酵體系。當反應體系有充足的外源電子供體添加時,可在單相反應器內進行CE反應,與秸稈水解酸化生成的電子受體(乙酸)反應合成MCFAs;當電子供體來源不足時,搭建兩相厭氧發酵體系,將創造的秸稈轉化乙醇或乳酸的發酵體系與水解酸化體系結合,在第二相反應器中合成MCFAs。兩相反應器可實現電子供體、受體生產與CE反應均在最適條件下進行,具有提高MCFAs生產的穩定性,減少有機負荷、氫分壓、CO2含量等不匹配帶來負面影響的優勢。Zhang等[70]評價了實驗室規模兩相反應器中外加電子供體MCFAs的生產效率,將CSTR中生成的8.4 g COD·L-1乳酸加入CE反應器(UAFR)中,連續發酵90 d,己酸酯的最大產率為1.5 g COD·L-1·d-1,選擇性為68.3%,對其進行經濟評價分析發現,該過程每噸新鮮生物質的總收益約為62.9美元。目前,兩相反應模式已在工業上得到應用[71],德國Darmstadt有機垃圾處理廠(處理量 13 200 t·年-1)將秸稈、食物垃圾等有機廢棄物進行收集分選,破碎后進行水解酸化,酸化后的固體用于好氧堆肥,液體進入厭氧反應器進行CE反應產MCFAs,同時對產品進行提取分離,該工廠每年提煉MCFAs 15.5~22.5 t,脂肪酸轉化產值6萬歐元,沼氣5 000 m3,固體有機肥1 760 t。
5.2 熱化學轉化合成MCFAs工藝
秸稈熱解生成合成氣(H2、CO2和 CO),并進一步轉化為MCFAs,其中,合成氣轉化MCFAs可通過單相或兩相工藝實現。當合成氣發酵和CE的溫度范圍、pH、微生物所需的生長營養物大致相同時,可以在單相反應器中完成;當CE階段缺少電子供體和電子受體時,秸稈熱解后進行合成氣發酵生成乙醇和乙酸,然后加入CE反應相,通過兩相反應器完成MCFAs的合成。其中,CSTR、鼓泡塔反應器(slurry bubble column reactor,SBCR)、生物膜反應器(membrane bio-reactor,MBR)、滴流床反應器(trickle-bed reactor,TBR)是合成氣發酵常用的反應器,除此之外,中空纖維膜生物反應器(hollow fiber membrane reactor,HFMBR)憑借生物膜對微生物的富集作用以及裝置氣體損耗小的優勢,被廣泛應用到合成氣轉化MCFAs體系。Zhang等[42]首次證實了混合微生物利用H2和CO2合成氣生成MCFAs,己酸鹽和辛酸鹽的含量分別0.98和0.42 g·L-1,其中氣體率接近100%,表明了HFMBR是具有發展潛力的合成氣發酵技術。有研究[34]通過兩相工藝,利用Clostridium kluyveri的純培養物,從合成氣發酵產物中獲得了更高的己酸 生 產 率 (40 mmol·L-1)、辛 酸 生 產 率 (約2.19 mmol·L-1)和>90%的碳轉化率。到目前為止,將難于生物降解的秸稈進行熱解氣化,得到富含CO2、CO和H2的合成氣,再利用合成氣作為底物發酵生產MCFAs是一項具有巨大應用潛力的生產技術,很多的研究依舊處于實驗室階段,距離達到商業化還存在諸多限制。
6 對我國秸稈綜合利用的啟示
秸稈有機質含量豐富,是厭氧合成MCFAs的理想原料,研究MCFAs合成技術的主要目的是為秸稈離田利用價值尋找新的突破口,為秸稈產業鏈循環經濟提供新的增長點。當前,我國處理秸稈的主要方式仍是還田,面對秸稈產量大、耕地少、種植岔口緊的供需矛盾,在保證土壤健康及不影響下茬作物種植的情況下,加大還田量難度較大,且還田產生的經濟效益低,難以帶來收益,根本出路仍是離田利用。許多研究和案例已經證明,影響秸稈離田做肥料、做飼料、做能源、做基料甚至做原料的產業鏈的限速步驟均為收儲運成本高、產品價值低、市場空間小,如秸稈飼料需混配其他營養物質調質后才有較好的市場價值、沼氣/生物天然氣并網發電或做車用燃料市場準入困難等。因此,傳統的秸稈綜合利用方式亟需升級,高值利用將是秸稈綜合利用的一個重要方向。
秸稈兩大轉化路徑(生物化學、熱化學)均具備高效合成MCFAs的潛力,發酵技術設備與厭氧發酵產沼氣幾乎相同,一些電子供體/受體富集、接種、水力停留時間、pH等調控技術趨于成熟,秸稈產酸速率不斷提高,可實現MCFAs的定向連續生產,現有技術模式示范證明了秸稈轉化MCFAs可取得較好的經濟效益,為秸稈的高值利用提供了良好的技術路徑和技術儲備。秸稈高效合成MCFAs技術仍需進一步探索,復雜發酵體系中間產物定向代謝路徑、微生物功能網絡形成機制等科學問題尚不清楚,產酸周期長、發酵體系緩沖能力不足、過程不穩定等技術問題亟需突破。因此,下一步研究將根據秸稈原料特性,研究協同生產乙醇-乙酸、乳酸-乙酸及CE轉化MCFAs過程的代謝流,建立不同生態位層次的微生物關系圖譜,通過信息學、生物學、統計學交叉融合等方式挖掘關鍵微生物及產酸潛力,結合反應器結構優化,攻克接種物富集、抗酸沖擊等關鍵技術,提高秸稈連續穩定產酸水平。
