水稻種子胚形態(tài)與胚乳組成對稻米留胚特性的影響
2023-05-18 02:14:36馬兆惠石一涵程海濤宋文雯路連吉劉仁廣呂文彥
中國水稻科學 2023年3期
馬兆惠 石一涵 程海濤 宋文雯 路連吉 劉仁廣 呂文彥*
水稻種子胚形態(tài)與胚乳組成對稻米留胚特性的影響
馬兆惠1,#石一涵1,#程海濤1宋文雯1路連吉1劉仁廣2呂文彥1,*
(1沈陽農(nóng)業(yè)大學農(nóng)學院,沈陽 110866;2本溪市農(nóng)業(yè)綜合發(fā)展服務中心, 遼寧 本溪 117022;*通信聯(lián)系人,email:lwyzxr@syau.edu.cn)
【目的】稻米留胚能夠提高稻米營養(yǎng)品質(zhì),但稻米留胚特性與胚性狀、胚乳成分的關(guān)系等相關(guān)生物學機制并不明確。因此,有必要深入研究不同類型稻米品種留胚率差異及其影響因素。【方法】選取粳稻中軟米型和普通型兩類典型留胚材料及非留胚材料為試材,首先切片觀察兩類品種籽粒胚發(fā)育特點,繼而對比表觀直鏈淀粉含量(apparent amylose content, AAC)和離層果膠含量,最后以外源細胞分裂素(cytokinin, CTK)調(diào)控,比較胚性狀、胚乳組成及留胚率差異。【結(jié)果】與非留胚材料相比,軟米型和普通型留胚材料具有胚內(nèi)幅和外幅較大,胚發(fā)育速度快,胚體積較大的特點;籽粒充實度好,粒形更寬、厚;同時離層果膠含量也較高。外施CTK后,軟米型留胚材料中后期胚發(fā)育速度加快,普通型材料胚發(fā)育減緩;兩類材料的AAC、果膠含量、胚比重及留胚率均提高。【結(jié)論】除胚性狀對兩類品種留胚率均有明顯影響外,胚乳組成主要影響軟米型材料留胚特性,而粒形是影響普通型材料留胚特性的重要因素。CTK則通過影響籽粒不同性狀對不同類型材料留胚率進行調(diào)控。這為后續(xù)高留胚水稻品種的培育和栽培調(diào)控提供了理論依據(jù)和指導方向。
留胚率;胚發(fā)育;胚乳成分;粒形;細胞分裂素
隨著生活水平的提高,人們越來越重視大米的營養(yǎng)品質(zhì)。稻胚含有脂肪、氨基酸、維生素等豐富的營養(yǎng)元素[1],但在加工過程中大多因碾磨而隨胚丟失,導致精米無法實現(xiàn)營養(yǎng)與食味兼?zhèn)洌诿卓诟星芳选H粝朐诒WC稻米優(yōu)良口感之余進一步提高其營養(yǎng)品質(zhì),留胚米毫無疑問成為了一個折衷的選擇,留胚品種的研究成為了稻米研究的一個新方向。探索不同類型留胚品種的留胚機制,有助于進一步明確留胚品種高留胚率的特異性及其形成規(guī)律,對于選育留胚米具有重要意義。
留胚米通常指糙米經(jīng)過碾磨加工之后,留胚率在80%以上或胚的質(zhì)量占整體質(zhì)量2%以上的稻米[2]。從表型來看,稻米留胚就是胚與胚乳粘連沒有被完全破壞,因此,凡是胚體有殘留,不論留有多少,均屬于留胚米。稻米含胚與加工精度、品種、粒形和胚發(fā)育情況等因素有關(guān)。一般稻米的留胚率達80%以上時,長寬比小于1.45,即一般短粗形稻米利于留胚[3]。反田嘉博[4]研究發(fā)現(xiàn),通常胚越長,胚的伸出(胚的外側(cè))大,而嵌入(胚的內(nèi)側(cè))小,則胚越容易脫落。籽粒發(fā)育過程中,盾片外側(cè)胚乳分化出一層特化細胞(即離層),胚通過離層與胚乳相聯(lián)系[5],離層組織的粘連強度決定胚與胚乳的結(jié)合力[6]。猜測離層(即胚與胚乳間)存在某種黏性物質(zhì),而這類黏性物質(zhì)含量的差異導致水稻留胚率的差異。水稻淀粉可能是胚與胚乳之間粘連物質(zhì)之一,直鏈淀粉含量(AAC)較高則黏性較低[7],AAC較低的品種一般黏性較大[8]。植物細胞之間最初主要是由果膠類物質(zhì)粘連在一起[9],而水稻的胚與胚乳之間是否也存在果膠類的黏性物質(zhì)?這類報道在谷類研究中尚不多見,值得進一步探討。已經(jīng)證實激素可通過調(diào)節(jié)纖維素酶和果膠酶活性來控制脫落,細胞分裂系(CTK)含量的變化不僅會引起果膠類物質(zhì)含量和離層特性的改變,從而引發(fā)胚脫落[10],而且對胚器官細胞發(fā)育、胚乳淀粉積累及籽粒發(fā)育也有一定影響[11-12]。因此,細胞分裂素是否也對留胚特性有影響?課題組前期研究發(fā)現(xiàn),灌漿好、胚乳充實度高是提高留胚率的關(guān)鍵因素,據(jù)此推測,CTK可能通過對胚乳發(fā)育的影響從而間接影響胚的著生、發(fā)育和脫落,但水稻上的相關(guān)研究還很少。
因此,本研究選擇留胚率有顯著差異的典型材料,首先通過觀察胚及籽粒發(fā)育動態(tài),得到兩類材料胚形態(tài)、體積、結(jié)構(gòu)差異,分析留胚材料的胚發(fā)育特性,繼而測定籽粒AAC和離層果膠含量,研究胚與胚乳粘連程度和特點,最后以外源激素CTK調(diào)控,比較胚性狀、理化指標及留胚率差異,以期初步探明水稻高留胚率的生物學特征。本研究旨在揭示影響水稻留胚率的主要因素,并根據(jù)不同類型水稻品種的不同留胚影響因素,確定提高稻米留胚率的栽培調(diào)控方法,以期應用于水稻高留胚育種和栽培工作。
1 材料與方法
1.1 試驗材料
選擇2個留胚和2個非留胚粳稻材料為試材,分別為彥粳軟玉1號(軟米留胚型,8月4日齊穗)、1419-8(軟米非留胚型,為彥粳軟玉1號后代,8月3日齊穗)、2045-5(普通留胚型,8月5日齊穗)和19-3(普通非留胚型,8月4日齊穗)。
1.2 田間栽培管理
試驗于沈陽農(nóng)業(yè)大學試驗水田(41°83'N、123°57'E)展開,試驗地基本情況為堿解氮65.70 mg/kg,有效磷13.59 mg/kg,速效鉀120.40 mg/kg,有機質(zhì)25.63 g/kg,土壤pH 5.53,電導率(土壤含鹽量與水土比為5:1) 274.70mS/m。2020年4月中旬播種,5月下旬插秧。田間采用隨機區(qū)組排列,3次重復,每個材料3 m行長,6行區(qū),行株距為30 cm×13.3 cm,每穴2株苗。試驗田基施硫酸鉀型復合肥50 kg/hm2。
在孕穗期時,葉片噴施細胞分裂素(6-芐基氨基嘌呤)。細胞分裂素用95%的乙醇完全溶解后,配制成400 mg/L母液,施用時再稀釋成各梯度濃度,對照為純凈水加入等體積的95%乙醇。處理時間選擇在16∶00-18∶00,每個處理噴施溶液1 L。經(jīng)前期預備試驗對比,最終采用CK(空白對照)、H(20 mg/L)兩個處理。激素處理后,標記同一天開花、長勢均勻一致的籽粒,根據(jù)稻胚發(fā)育進程,分別于標記后9、12、30 d取樣,每次取穗中上部強勢粒40粒,放入裝有FAA固定液(90 mL 70%乙醇+5 mL 38%甲醛+5 mL冰醋酸+5 mL甘油)的離心管后置于4℃冰箱內(nèi)貯存,用于測定有關(guān)性狀。
1.3 性狀測定
1.3.1 千粒重
稻谷自然干燥后,采用稻谷計數(shù)板量出3份稻谷,每份200粒,稱量后取平均值,將結(jié)果乘以5,獲得水稻樣品的千粒重。
1.3.2 粒形
用萬深SC-E型種子大米外觀品質(zhì)檢測儀測定長寬比、粒長和粒寬。隨機選取10~15粒籽粒,用游標卡尺測定其厚度,求平均值。
1.3.3 胚比重
將100粒糙米稱重(1),將胚部用鑷子剝離,稱量其重量(2),其胚比重=(2/1)×100%,重復3次,取平均值。
1.3.4 留胚率
稻谷利用韓國KMT-400B6脫殼機脫殼,Keet精米機碾精(碾精度在90%±1%)得到整精米,然后按照四分法取樣:10 g整米試樣排成一粒米厚度的正方形,用分樣板在樣品上劃兩條對角線,分成4個三角形,從每個三角形中,各隨機數(shù)出25粒,100粒計為一個試樣,然后取3份平均試樣,將試樣攤在白色潔凈的搪瓷盤里,在明亮處(應采用背向光)逐粒鑒別,并按水稻留胚級別[13]分類,半胚及半胚以上米粒數(shù)之和計為留胚米,即該試樣留胚率。
1.3.5 米粉的制備
整糙米采用FSⅡ旋風式粉碎磨制成米粉,分成兩等份,一份過80目篩,用于總果膠含量的測定;另一份過100目篩,用于表觀直鏈淀粉含量的測定。
1.3.6 直鏈淀粉含量的測定
參照《ISO 6647-2:2007直鏈淀粉含量測定方法》,其中標準樣品來自于中國水稻研究所(CRRI)。
1.3.7 總果膠含量的測定
總果膠含量測定參照張雪丹[14]的方法。
1.3.8 胚形態(tài)的觀察
胚形態(tài)觀察采用石蠟切片和徒手切片相結(jié)合的方法,石蠟切片制作參照李娜[15]的方法;徒手切片方法具體如下:將糙米放在蒸餾水中浸泡15 min,取出后放在吸水紙上吸干水分。將整個籽粒從中部截斷,然后將含有胚的半個籽粒立在桌面上,從籽粒上端中間與胚橫截面水平角度用雙面刀片切開,取兩側(cè)第一片,用于顯微鏡觀察,測量胚部各性狀。每個材料做25~30片,除去異常值(|測量值-平均值|/平均值>15%),選擇形態(tài)、大小類似的作為最終計算數(shù)值。
1.3 數(shù)據(jù)分析
運用DPS 15.10數(shù)據(jù)處理系統(tǒng)進行多重比較和相關(guān)分析,用Sigmaplot作圖軟件繪圖。
2 結(jié)果與分析
2.1 不同類型材料胚性狀差異比較
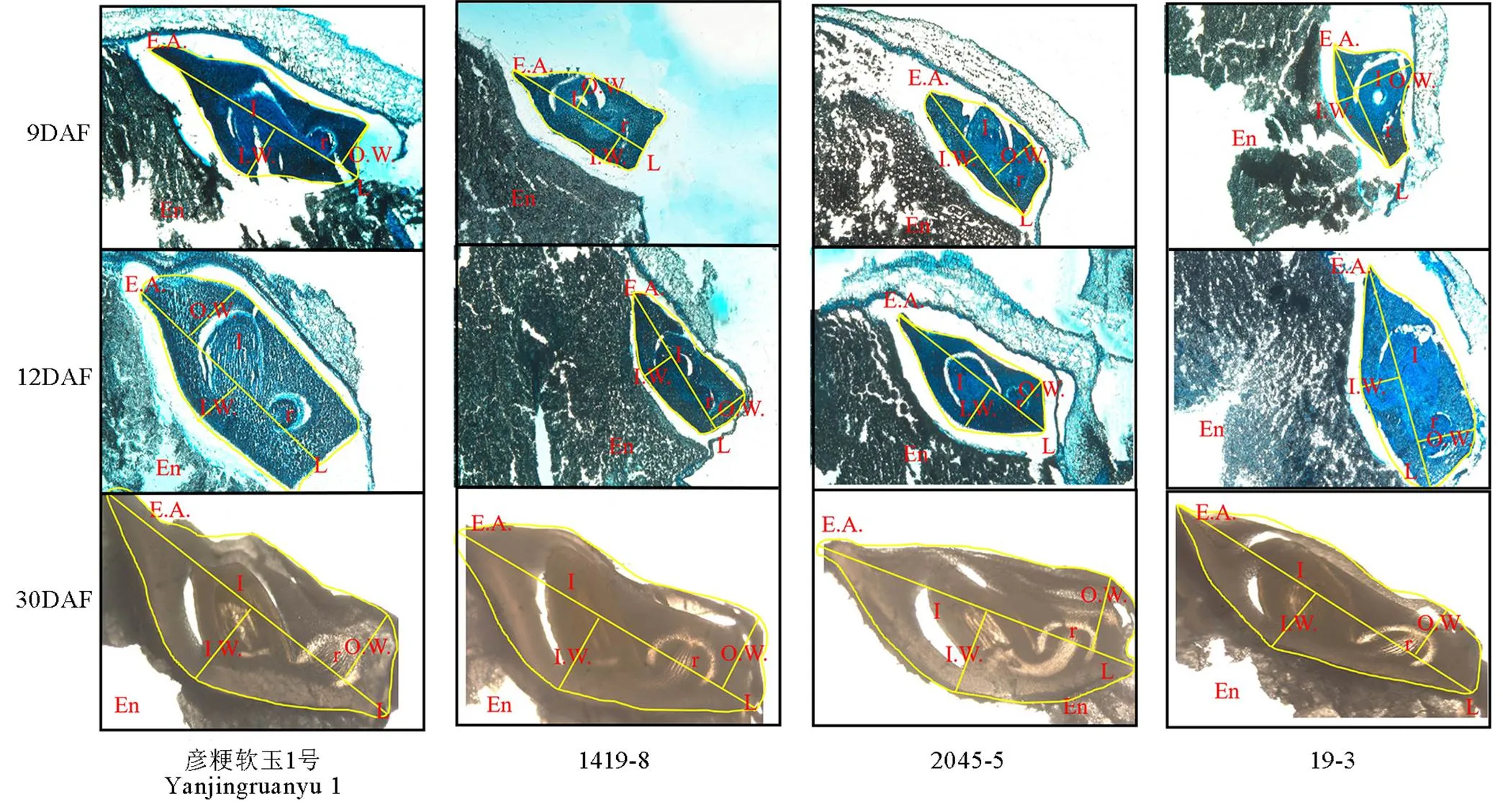
切片觀察軟米型和普通型兩類材料花后9 d、12 d和30 d胚部形態(tài)特征(圖1,表1),結(jié)果表明,在胚發(fā)育前期與中期,軟米型留胚材料彥粳軟玉1號的內(nèi)幅、胚長、外幅、胚面積及胚發(fā)育速度均顯著大于非留胚材料1419-8,并在整個時期保持了較大的胚體積(圖1)。普通型留胚材料2045-5在花后9 d胚部各指標顯著小于非留胚材料19-3,但其在整個生育期的胚發(fā)育速度較19-3快,因此在花后12 d至成熟期,2045-5胚的內(nèi)幅、外幅和面積顯著大于19-3,表現(xiàn)出大胚的特點。
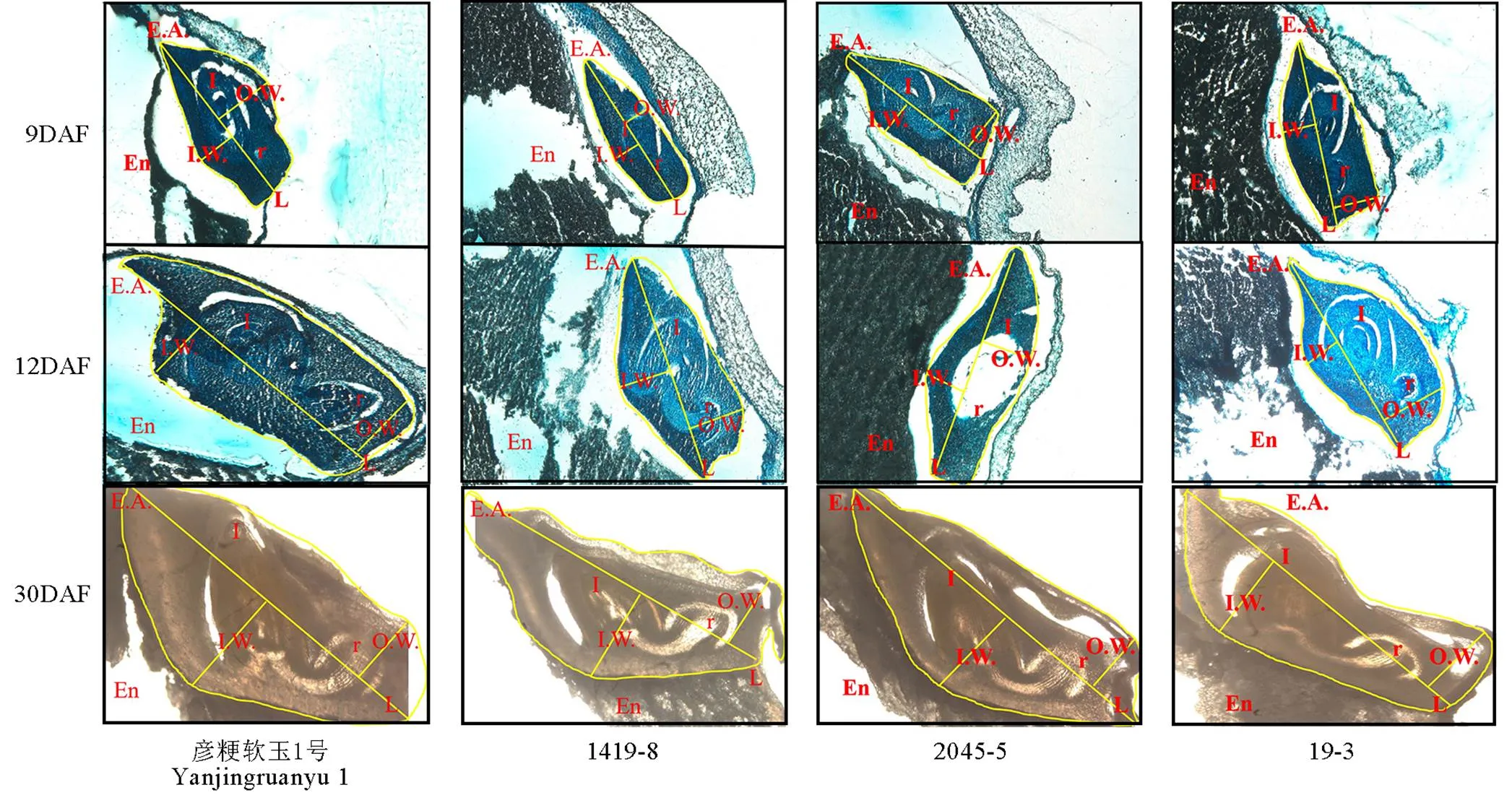
EA-胚面積;En-胚乳;L-胚長;I-胚芽;IW-內(nèi)幅;OW-外幅;R-胚根。DAF-花后天數(shù)。
Fig. 1. Embryo-specific morphologies of different materials at different developmental stages.
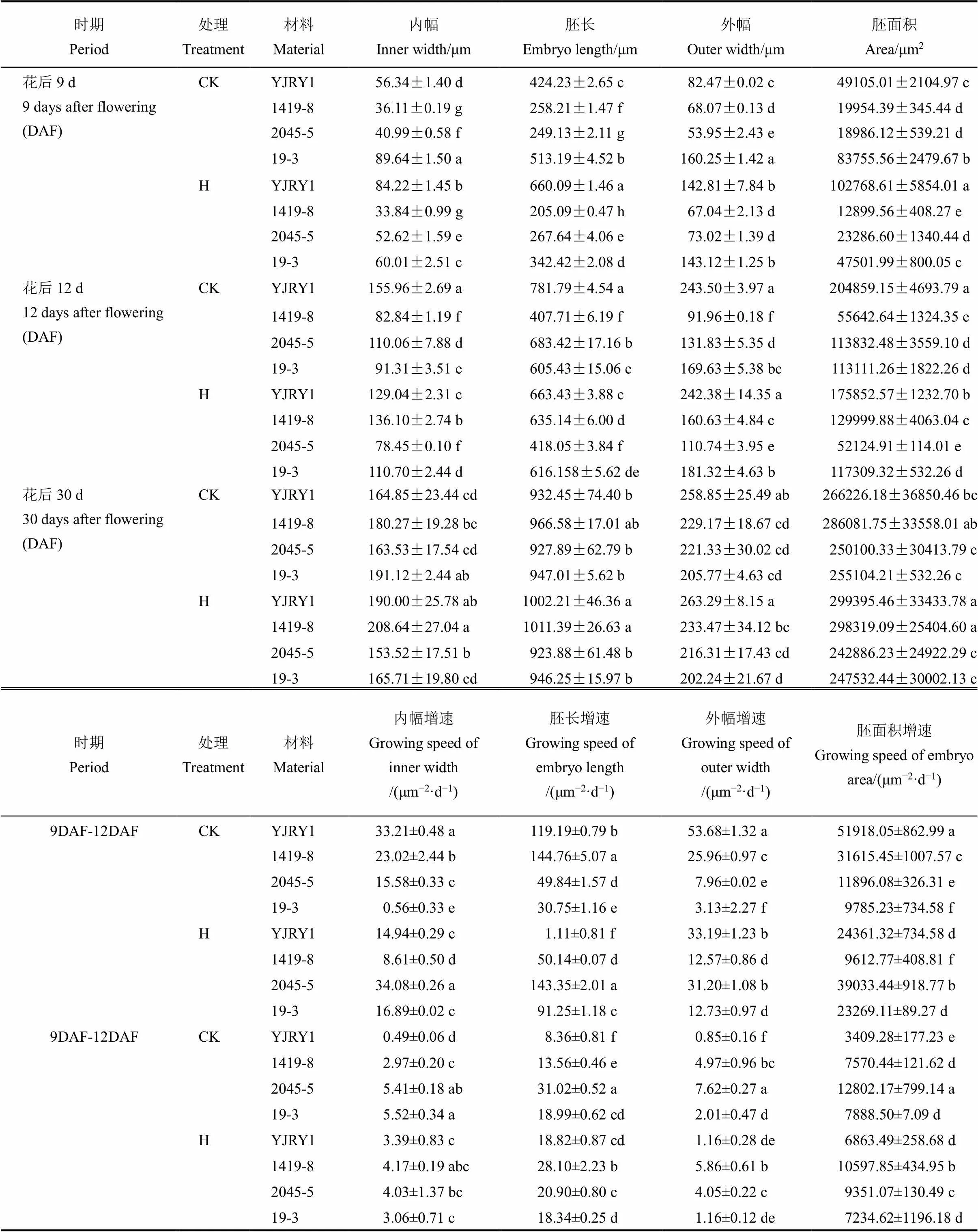
表1 施用細胞分裂素后不同時期留胚和非留胚材料胚部性狀及發(fā)育速率對比
不同小寫字母表示在0.05水平上差異顯著。CK-對照;H-施用細胞分裂系。
Values flanked by various lowercase letters are significantly different at the level of 0.05. CK, Control; H, Cytokinin application.
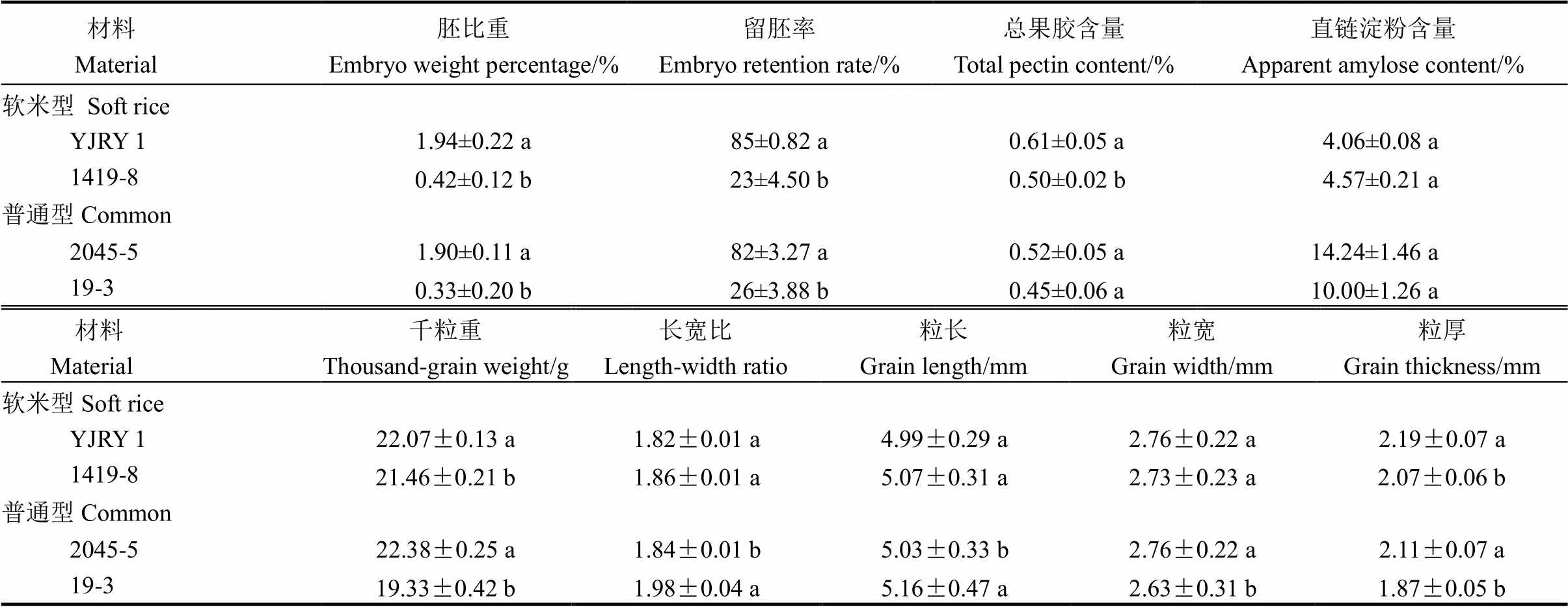
表2 不同類型材料留胚特性及籽粒性狀比較
YJRY 1-彥粳軟玉1號;不同字母表示同一類型品種間在5%水平上差異顯著。下同。
YJRY 1, Yanjingruanyu 1. Different letters indicate significance at the 5% level. The same below.
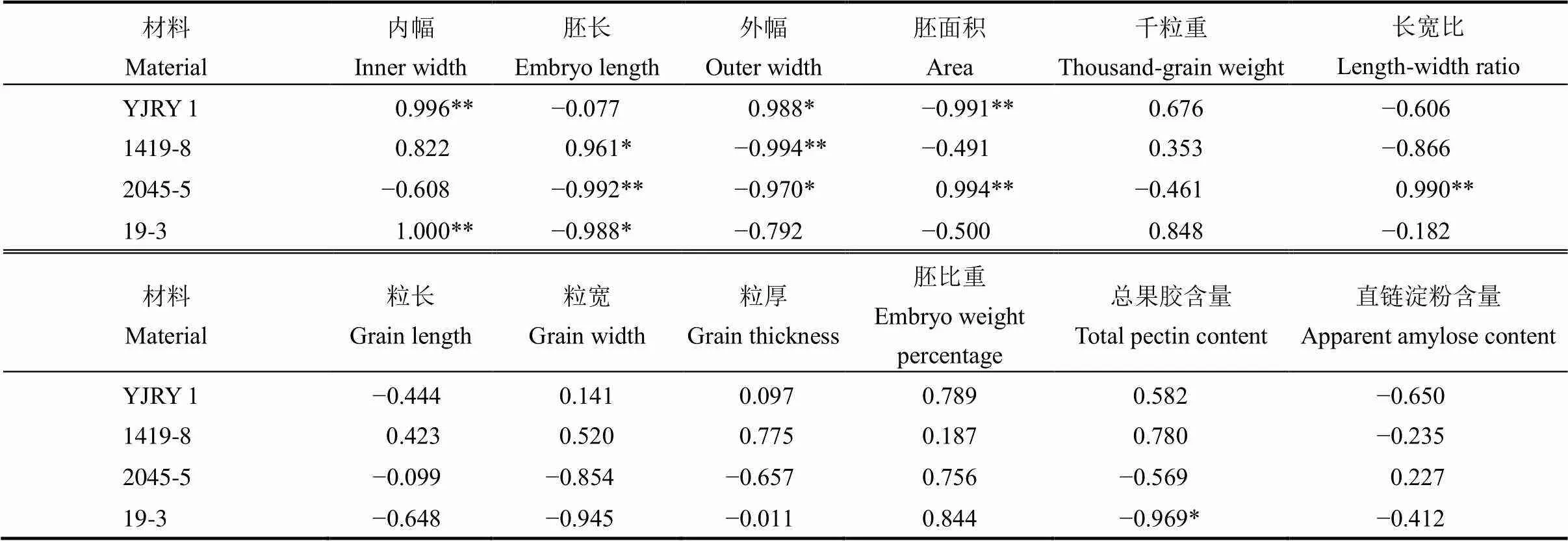
表3 不同類型材料籽粒性狀與留胚率的關(guān)系
=5。**表示該性狀與留胚率達極顯著相關(guān);*表示顯著相關(guān)。
=5. **,<0.01; *,<0.05.
2.2 不同類型材料離層物質(zhì)含量及留胚特性比較
兩類留胚材料的胚比重和留胚率均顯著高于兩類非留胚材料(表2),其中軟米型留胚材料的胚比重和留胚率最大;在離層物質(zhì)含量上,留胚材料的總果膠含量均大于同類型的非留胚材料,其中軟米型材料間的果膠含量差異達顯著水平。從直鏈淀粉含量看,材料間差異不顯著,但軟米型留胚材料的直鏈淀粉含量較非留胚材料低,因此胚乳黏性較高,這可能是其高留胚率的原因之一。
2.3 不同類型材料粒形及千粒重差異比較
研究結(jié)果表明,留胚材料與非留胚材料之間千粒重差異均顯著(表2),說明無論是軟米型還是普通型,留胚材料的籽粒充實度高于非留胚材料。從粒形看,留胚材料的籽粒更寬、厚,但粒長和長寬比值較非留胚材料低,這種差異在普通型材料間達到顯著,說明留胚材料更趨近于短圓粒形。
2.4 不同類型材料籽粒性狀與留胚率的關(guān)系
籽粒性狀中,胚形態(tài)與留胚率關(guān)系最密切(表3)。軟米型留胚材料的內(nèi)幅、外幅與留胚率達到顯著正相關(guān);普通型留胚材料的胚面積與留胚率極顯著正相關(guān)。此外,增加胚比重對提高留胚率起促進作用。從粒形看,長寬比對普通型留胚材料的留胚率影響更明顯;粒厚增加能提高軟米型材料留胚率。從離層成分看,軟米型材料果膠含量與留胚率呈正相關(guān),直鏈淀粉含量與留胚率呈負相關(guān),而普通型材料果膠含量與留胚率呈負相關(guān),說明離層成分對留胚率的影響因材料類型而異。
EA-胚面積;En-胚乳;L-胚長;I-胚芽;IW-內(nèi)幅;OW-外幅;R-胚根。
Fig. 2. Comparison of embryo morphology of different types of embryo-remaining material after cytokinin application.
2.5 外施CTK對籽粒各性狀及留胚特性的影響
與對照相比,外施CTK后使發(fā)育前期(花后9 d)留胚材料的胚各性狀值均有所增加(圖2,表1),胚發(fā)育速度加快(花后9~12 d)。在發(fā)育中后期(12~30 d),CTK抑制普通型材料的胚發(fā)育速度,加速軟米型材料胚發(fā)育,因此最終成熟時,軟米材料胚各性狀值較高,普通型材料胚性狀值降低。此外,外施CTK增加了胚比重和留胚率(表4),其中兩類非留胚材料增幅較大,增幅達36.1%和46.5%。果膠含量因外施CTK而增加,其中普通型留胚材料的增幅明顯(30.9%)。外施CTK使各材料的千粒重降低,軟米型材料粒厚減小,普通型材料粒厚增加,留胚材料粒寬下降而長寬比增加,非留胚材料粒寬增加而長寬比下降。
2.6 CTK施用量與留胚特性的相關(guān)性分析
外施CTK對不同類型材料的胚發(fā)育特性、籽粒性狀及胚乳成分含量有不同程度的影響(表4)。CTK施用量與留胚材料各胚性狀正相關(guān),但CTK施用量與非留胚材料的胚性狀負相關(guān)。從粒形看,留胚材料的粒長、粒寬、粒厚均與CTK施用量負相關(guān),其中粒長、粒厚受CTK負影響較大。離層成分中,果膠含量和直鏈淀粉含量與CTK用量均呈顯著或極顯著正相關(guān)。留胚特性中,留胚率和胚比重均與CTK施用量表現(xiàn)為正相關(guān),其中留胚材料與CTK施用量的相關(guān)更顯著。
3 討論
3.1 不同類型留胚材料胚性狀及籽粒性狀差異比較
前人對胚的發(fā)育進程研究雖已基本明確[16-19],但不同類型、不同留胚率品種間胚發(fā)育是否存在差異?本研究結(jié)果表明,軟米型留胚材料在早期胚發(fā)育速度快,后期發(fā)育速度慢,且胚體積始終大于非留胚材料,這不僅符合早期胚迅速生長發(fā)育的特征[20],更證明早期是軟米型材料胚發(fā)育及胚體積形成的關(guān)鍵時期。而普通型留胚材料在整個生育期均表現(xiàn)出較非留胚材料快的發(fā)育速度,因此最終成熟期時,二者胚大小近乎一致,說明普通型材料的胚建成貫穿整個胚發(fā)育時期,這驗證了前人的結(jié)論[17-18]。同時,還發(fā)現(xiàn)留胚米的胚質(zhì)地較為緊密,非留胚米胚極易碎裂,難以取得完整胚。因此,進一步研究可以從留胚米胚部質(zhì)地方向著手。前期研究發(fā)現(xiàn),留胚率高的籽粒具有較低的AAC[21],說明低AAC有助于留胚,而本研究中只有軟米型材料符合這一特征,因此AAC作為判斷留胚率高低的指標應因材料類型而異。Michal等[22]通過紅醋栗實驗證明了果膠含量與黏度特性之間存在關(guān)系,但關(guān)于果膠與黏性的報道主要集中在漿果類,在谷物類研究中尚屬空白。我們的研究發(fā)現(xiàn),留胚材料的總果膠含量較大,故此推測總果膠含量差異使胚與胚乳粘連程度不同,是影響留胚的原因之一,此結(jié)果與張雪丹[14]的研究結(jié)果相似。
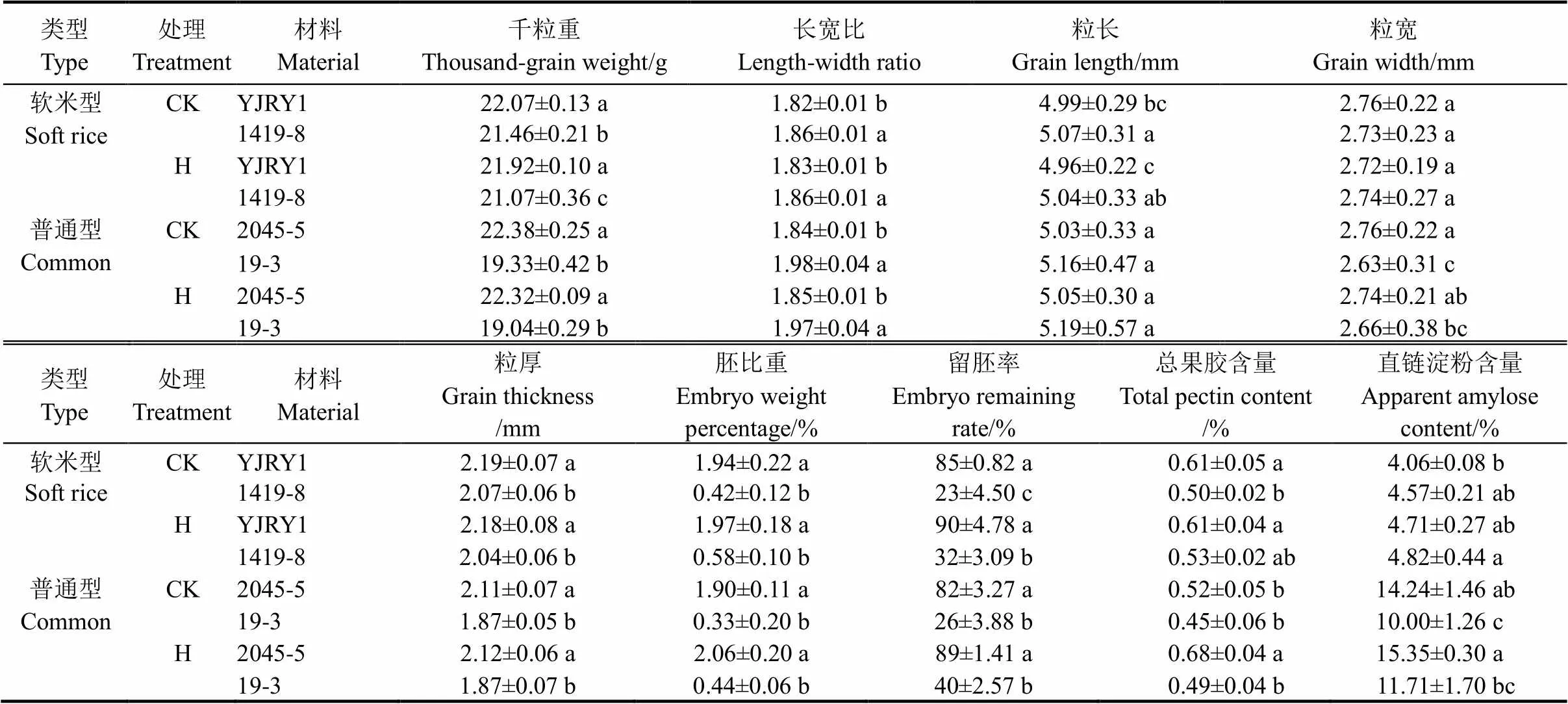
表4 不同類型材料外施細胞分裂素后籽粒性狀及理化性質(zhì)的變化
不同字母表示在5%水平上差異顯著。
Different letters indicate significance at the 5% level.
高留胚品種充實度好,千粒重大,籽粒寬厚,這與前期研究結(jié)果相同[23],且這種特點在普通型留胚品種表現(xiàn)明顯,說明粒形和充實度是影響普通型品種間留胚率的重要因素。此外,通過分級碾精加工發(fā)現(xiàn)(圖3),隨碾磨時間增加,留胚材料的留胚類型由全胚(A)和平胚(B),轉(zhuǎn)變成少量半胚(C)和大部分的殘胚(D);而非留胚材料的留胚類型由半胚(C)轉(zhuǎn)變成殘胚(D)和無胚(E)。這種留胚類型的差異,證明留胚材料的胚與胚乳粘連緊密且抗碾磨。而本研究中發(fā)現(xiàn)該類材料在胚發(fā)育、籽粒性狀及胚乳成分方面與非留胚材料相比,存在一定特點和差異,從而具有高留胚率的特性。
3.2 影響不同類型品種留胚率的因素分析
影響稻米留胚的因素多樣[24],留胚首先與胚形態(tài)結(jié)構(gòu)有關(guān),當胚嵌入米粒較淺且自身體積大時,則受碾磨能力弱,易脫落[4]。本研究發(fā)現(xiàn)留胚材料具有米胚外形鈍圓,且胚整體向粒內(nèi)伸展長度/向外伸展長度比例較大的特點,因此更抗碾磨。從相關(guān)分析看,軟米型留胚材料的胚嵌入方式是影響留胚率的主要因素,而普通型留胚材料的留胚率主要由胚面積決定,說明不同類型留胚材料的留胚率由不同的胚形態(tài)性狀影響。胚乳的主要組成成分是淀粉,其中AAC是影響米粉黏性的主要因子[25-26],通常AAC低更有利于胚乳與胚相粘連。本研究中,與非留胚材料相比,軟米型留胚材料的AAC低且其與留胚率負相關(guān),進一步證明較低的AAC是保證軟米型材料高留胚率的因素之一。果膠作為植物細胞之間最初的粘合物質(zhì),其濃度越大則黏性越強[27]。本研究也發(fā)現(xiàn)總果膠含量高是導致軟米型材料具有高留胚率的重要因素,但普通型材料總果膠含量高不利于留胚率,是否因果膠種類及占比不同而造成差異,還有待進一步研究。
水稻粒形一般指的是粒長、粒寬、粒厚以及長寬比[28],稻米加工時,粒形影響籽粒碾磨的接觸面積,因此是留胚的一個決定因素。有研究表明,水稻留胚率與粒長、長寬比顯著負相關(guān),與粒厚、粒寬正相關(guān)[21],也有研究得出留胚率與粒長、寬、長寬比不顯著負相關(guān)的結(jié)果[29]。課題組前期研究發(fā)現(xiàn),對于一般材料而言,粒形越長的稻米,耐磨性越差[23],長寬比增加,留胚率則下降[30]。而本研究中,普通型留胚材料的長寬比與留胚率呈極顯著正相關(guān),說明對于較秈稻粒形更短圓的粳稻品種而言,適當提高長寬比可以提高留胚率。此外,粒重對胚比重有顯著影響[31],本研究中兩類留胚材料的胚比重與留胚率均正相關(guān),這說明,粒重通過影響胚比重進而影響留胚率,即籽粒充實度是影響留胚率的基本因素之一。
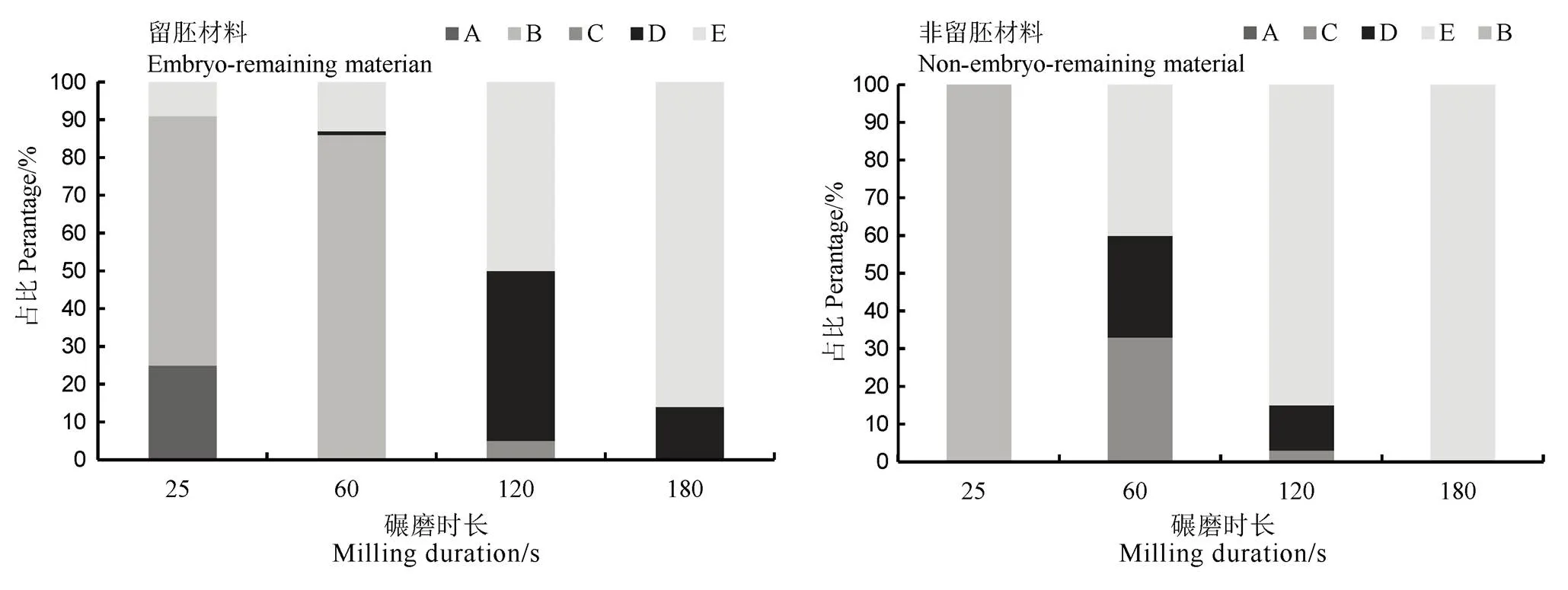
A-全胚, 糙米研磨后,胚幾乎沒有磨損;B-平胚,糙米碾碎后,剩下的胚與米尖相切;C-半胚, 糙米磨碎后,將一半胚磨掉;D-殘胚, 糙米研磨后,只剩下部分胚;E-無胚, 糙米研磨后,所有的胚都被研磨掉,沒有任何殘留物。
Fig. 3. Changes of embryo remaining types of embryo-remaining rice and non-embryo-remaining rice under different milling time.

表5 CTK施用量與不同類型材料籽粒性狀及留胚率的相關(guān)性
**表示極顯著相關(guān);*表示顯著相關(guān)。
**,<0.01;*,<0.05.
3.3 外施CTK對不同類型材料籽粒性狀及留胚特性的影響
細胞分裂素(CTK)對籽粒發(fā)育有重要影響[32],可使胚增大[33]。本研究也發(fā)現(xiàn),與對照相比,CTK處理加速了軟米型材料中后期的胚發(fā)育,使其形成大胚;但普通型材料在CTK處理下整個生育期胚發(fā)育速率下降,胚值減小,說明CTK處理可能抑制普通型材料胚細胞的分生組織[34]。即外源施用CTK不僅對胚發(fā)育的影響時期不同,影響結(jié)果也因材料類型而異。CTK誘導的水稻基因Cga1下調(diào)表達時,籽粒變得細長[35-36],即當CTK濃度較低時,籽粒發(fā)育會趨于細長形。本研究中,與對照相比,CTK處理提高了留胚材料的長寬比,而普通型留胚米的長寬比與留胚率呈極顯著正相關(guān),因此,普通型材料可通過CTK調(diào)控粒形進而提高留胚率。
前人研究已經(jīng)證實,內(nèi)源激素能通過調(diào)節(jié)纖維素酶和果膠酶活性引起果膠類物質(zhì)含量和離層黏性的改變,進而影響留胚[37-39]。對于留胚米的高留胚率機理,有研究推測其胚和胚乳間具有較高含量的果膠,導致離層粘連特性變化。本研究發(fā)現(xiàn)外施CTK對果膠含量提高有明顯的促進作用,軟米類材料可通過外施CTK,顯著提高果膠含量,進而提高留胚率。此外,CTK有調(diào)控同化物積累和胚乳發(fā)育[40],促進淀粉積累[41]等作用,但其對水稻留胚特性的影響方面尚未見報道。本研究中,兩類材料的AAC與CTK量顯著正相關(guān),證明CTK在水稻中也具有促進淀粉積累的作用,并通過調(diào)控AAC含量影響不同類型材料的留胚率。
4 結(jié)論
本研究結(jié)果表明,留胚材料總體上胚發(fā)育速度快,胚體積較大,胚質(zhì)地緊密。軟米型材料胚嵌入胚乳部位較深,并具有較高果膠含量。普通型材料的胚面積大,粒形寬厚。通過外源CTK調(diào)控發(fā)現(xiàn),施用CTK可以促進軟米型材料胚內(nèi)幅、外幅發(fā)育,并提高離層黏性物質(zhì)含量;而對于普通型材料,CTK主要增加胚比重和粒長。因此,CTK對不同類型材料留胚率的調(diào)控是通過影響水稻籽粒不同性狀實現(xiàn),可根據(jù)材料特性進行留胚率調(diào)控。
[1] 朱一帆, 馮亞斌, 林軒, 周美圓, 王中華. 胚芽米營養(yǎng)成分研究[J]. 科技通報, 2013,29(9): 51-54.
Zhu Y F, Feng Y B, Lin X, Zhou M Y, Wang Z H. Nutritional components analysis of germ-remaining rice[J].,2013,29(9): 51-54. (in Chinese with English abstract)
[2] 宋幼良, 吳殿星. 高留胚糯稻突變體原胚糯的選育與營養(yǎng)成分分析[J]. 核農(nóng)學報, 2017, 31(9): 1672-1677.
Song Y L, Wu, D X. Breeding and nutrient content analysis of glutinous rice mutant Yuanpeinuo with high remained germ[J].,2017,31(9): 1672-1677. (in Chinese with English abstract)
[3] Liang J, Li Z, Tsuji K. Milling characteristics and distribution of phytic acid and zinc in long-, medium-and short-grain rice[J]., 2008, 48(1): 83-91.
[4] Tanda Y. Research on embryo detachment by polishing early cultivated rice[J].1962, 30: 9-13. (in Japanese)
[5] Yang W, Gao M, Yin X, Liu J, Xu Y, Zeng L, Li Q, Zhang S, Wang J, Zhang X, He Z. Control of rice embryo development, shoot apical meristem maintenance, and grain yield by a novel cytochrome P450[J]., 2013, 6(6): 1945-1960.
[6] Wan Z, Xia J, Kang D. The Research progress and prospect analysis of germ-remaining rice whitener[J].:,2008, 15(2): 84-87.
[7] Kong X, Zhu P, Sui Z, Bao J. Physicochemical properties of starches from diverse rice cultivars varying in apparent amylose content and gelatinization temperature combinations[J]., 2015,172: 433-440.
[8] Zhou L, Sheng W, Jun W, Wang C, Liu Q, Deng Q. Differential expressions among five Waxy alleles and their effects on the eating and cooking qualities in specialty rice cultivars[J]., 2015, 14(6): 1153-1162.
[9] Müller-Maatsch J, Bencivenni M, Caligiani A, Tedeschi T, Bruggeman G, Bosch M, Petrusan J, Droogenbroeck BV, Elstf K, Sforza S. Pectin content and composition from different food waste streams[J]., 2016,201: 37-45.
[10] Yuan R C, Burns J K. Temperature factor affecting the abscission, response of mature fruit and leaves to CMN-Pyrazole and ethephon in Hamlin oranges[J].,2004,129(3): 287-293.
[11] Jameson P E, Song J. Cytokinin: A key driver of seed yield[J]., 2015, 67: 593-606.
[12] Han Y, Yang H, Jiao Y. Regulation of inflorescence architecture by cytokinins[J]., 2014,5: 669.
[13] 李愛華. 米胚的營養(yǎng)價值和胚芽米的加工技術(shù)[J]. 糧食與飼料工業(yè), 1997(5): 9-12.
Li A H. The nutritive value of rice germ and the processing technology of germ-remaining rice[J]., 1997(5): 9-12. (in Chinese with English abstract)
[14] 張雪丹. 蘋果果膠制備工藝及研究進展[J]. 落葉果樹, 2009, 41(2): 22-25.
Zhang X D. Preparation technology and research progress of apple pectin[J]., 2009, 41(2): 22-25. (in Chinese with English abstract)
[15] 李娜.基因?qū)π∷牖ㄆ鞴侔l(fā)育的影響研究[D]. 沈陽: 沈陽農(nóng)業(yè)大學, 2015.
Li N. Functional analysis ofgene in spikelet development[D]. Shenyang: Shenyang Agricultural University, 2015. (in Chinese with English abstract)
[16] 楊弘遠. 水稻生殖生物學[M]. 杭州: 浙江大學出版社, 2005: 102-125.
Yang H Y. Rice Reproductive Biology[M]. Hangzhou: Zhejiang University Press, 2005: 102-125 (in Chinese).
[17] 王敏. 關(guān)于水稻穎界發(fā)育的研究[D]. 揚州: 揚州大學, 2011.
Wang M. Caryopsis development of rice[D]. Yangzhou: Yangzhou University, 2011. (in Chinese with English abstract)
[18] Itoh J I, Nonomura K I, Ikeda K. Rice plant development: From zygote to spikelet[J]., 2005,46(1): 23-47.
[19] Xu H, Zhang W, Gao Y, Zhao Y, Guo L, Wang J. Proteomic analysis of embryo development in rice () [J]., 2012, 235(4): 687-701.
[20] 章清杞, 陳健勇, 黃榮華, 張書標. 巨胚稻胚發(fā)育的解剖學觀察[J]. 核農(nóng)學報, 2008(2): 122-126.
Zhang Q Q, Chen J Y, Huang R H, Zhang S B. Anatomical observations on the developmental process of embryo in giant embryo rice[J]., 2008(2): 122-126(in Chinese with English abstract)
[21] 陳恒雪, 石一涵, 呂文彥, 馬兆惠, 程海濤, 陳云. 稻米留胚率測定方法篩選及留胚米相關(guān)特性研究[J]. 沈陽農(nóng)業(yè)大學學報, 2018, 49(3): 337-341.
Chen H X, Shi Y H, LW Y, Ma Z H, Cheng H T, Chen Y. Determination methods of rice plumule ratio and related characteristics of germ-remained rice[J]., 2018, 49(3): 337-341.
[22] Michal P, Joanna K, Marcin L, Magdalena W, Mateusz K, Jolanta J, Anna P. Red currant pectin: The physicochemical characteristic of pectin solutions in dilute and semi dilute regimes[J]., 2021, 113: 106420.
[23] Ma Z H, Chen H X, LW Y, Cheng H T, Chen Y, Wang Y B. Comparison of the chemical and textural properties of germ-remaining soft rice grains from different spikelet positions[J]., 2019, 96: 1137-1147.
[24] 熊海錚, 張寧, 孫健, 王寅, 舒小麗, 吳殿星. 水稻留胚米的營養(yǎng)價值、加工技術(shù)及產(chǎn)品開發(fā)研究進展[J]. 核農(nóng)學報, 2012, 26(7): 1031-1036.
Xiong H Z, Zhang N, Sun J, Wang Y, Shu X L, Wu D X. Research progress on nutritional value, processing technology and product exploration of embryo-retaining milled rice[J]., 2012, 26(7):1031-1036. (in Chinese with English abstract)
[25] Ahmed N, Tetlow I, Nawaz S, Iqbal A, Mubin M, Rehman M, Butt A, Lightfootc D, Maekawa M. Effect of high temperature on grain filling period, yield, amylase content and activity of starch biosynthesis enzymes in endosperm of basmati rice[J]., 2015, 95: 2237-2243.
[26] Teng B, Zeng R, Wang Y, Liu Z, Zhang Z, Zhu H, Ding X, Li W, Zhang G. Detection of allelic variation at thelocus with single segment substitution lines in rice (L) [J]., 2012, 30 (1):583-595.
[27] Xing F, Owen G J, Bradley L R, Osvaldo H C. Soluble pectin acts as a particle stabilizer of tomato suspensions: The impact on tomato products rheological characterization[J]., 2021, 139(3): 110508.
[28] 周海慶. 水稻粒形及千粒重的遺傳研究[D]. 吉林省延邊市: 延邊大學, 2014.
Zhou H Q. Genetic studies on the rice grain shape and 1000-grain weight[D]. Yanbian: Yanbian University, 2014. (in Chinese with English abstract)
[29] 趙飛. 粳稻稻米品質(zhì)及其與產(chǎn)量關(guān)系的遺傳分析[D]. 沈陽: 沈陽農(nóng)業(yè)大學. 2014.
Zhao F. Genetic analysis on relationship between quality and yield of Japanese rice[D]. Shenyang: Shenyang Agricultural University, 2014.(in Chinese with English abstract)
[30] 王少元, 孟慶虹, 嚴松, 張志宏, 高揚, 袁超, 盧淑雯, 河野元信. 北方粳稻留胚米碾磨工藝的研究[J]. 中國稻米, 2014, 20(5): 31-35.
Wang S Y, Meng Q H, Yan S, Zhang Z H, Gao Y, Yuan C, Lu S W, Motonibu K. Research on the milling processing with germ- remained japonica rice of northern China[J]., 2014, 20(5): 31-35. (in Chinese with English abstract)
[31] 張標金, 魏益華, 張祥喜, 羅林廣. 育成巨胚稻與其親本的糙米粒型性狀和礦物質(zhì)含量的比較分析[J]. 中國農(nóng)學通報, 2014, 30(6): 182-185.
Zhang BJ, Wei YH, Zhang XL, Luo LG. Comparative analysis on grain traits and mineral elements of brown rice from bred giant embryo rice and its parents[J]., 2014, 30(6): 182-185.
[32] 李志康, 嚴冬, 薛張逸, 顧逸彪, 李思嘉, 劉立軍, 張耗, 王志琴,楊建昌, 顧駿飛.細胞分裂素對植物生長發(fā)育的調(diào)控機理研究進展及其在水稻生產(chǎn)中的應用探討[J]. 中國水稻科學, 2018, 32(4): 311-324.
Li Z K, Yan D, Xue Z Y, Gu Y B, Li S J, Liu L J, Zhang H, Wang Z Q, Yang J C, Gu J F. Regulations of plant growth and development by cytokinins and their applications in rice production[J]., 2018, 32(4): 311-324. (in Chinese with English abstract)
[33] Werner T, Schmülling T. Cytokinin action in plant development[J]., 2009, 12(5): 527-538.
[34] 孫貝貝. 生長素和細胞分裂素的內(nèi)源合成控制根從頭發(fā)生的機制研究[D]. 上海: 上海師范大學, 2017.
Sun B B. Mechanism of specific function of plant hormones auxin and cytokinin during adventitious root regeneration[D]. Shanghai: Shanghai Normal University. 2017. (in Chinese with English abstract)
[35] Hudson D, Guevara D R, Hand A J. Rice cytokine in GATA transcription Factor1 regulates chloroplast development and plant architecture[J]., 2013, 162(1): 132-144.
[36] Li J, Nie X, Tan J L. Integration of epigenetic and genetic controls of seed size by cytokine in[J]., 2013, 110 (38): 15479-15484.
[37] Ashikari M, Wu J, Yano M. Rice gibberellin-in-sensitive dwarf mutant geneencodes the alpha-subunit of GTP-binding protein[J]., 1999, 96(18): 10284-10289.
[38] Fujisawa Y, Kato T, Ohki S. Suppression of the hetero trimetric G protein causes abnormal morphology, including dwarfism, in rice[J]., 1999, 96(13): 7575-7580.
[39] Nayar S, Sharma R, Tyagi A, Kapoor S. Functional delineation of rice MADS29 reveals its role in embryo and endosperm development by affecting hormone homeostasis[J]., 2013, 64: 4239-4253.
[40] Zhao Y, Hu Y, Dai M, Huang L, Zhou D. The WUSCHEL-Related homeobox geneis required to activate Shoot-Borne crown root development in rice[J]., 2009, 21(3): 736-748.
[41] 朱艷梅, 羅興錄, 樊吳靜.木薯內(nèi)源細胞分裂素含量對塊根淀粉積累的影響[J]. 南方農(nóng)業(yè)學報,2016, 47(8): 1279-1284.
Zhu Y M, Luo X L, Fan W J.Effects of endogenous cytokinin content on starch accumulation in root tuber of cassava[J]., 2016, 47(8): 1279-1284.(in Chinese with English abstract).
Effects of Embryo Morphology and Endosperm Composition on Embryo-remaining Characteristics in Rice
MA Zhaohui1,#, SHI Yihan1,#,CHENG Haitao1, SONG Wenwen1, LU Lianji1, LIU Renguang2, Lü Wenyan1,*
(Agronomy College, Shenyang Agricultural University, Shenyang 110866, China; Benxi Agricultural Comprehensive Development Service CenterBenxi 117022, China; Corresponding author, email:)
【Objective】The embryo of embryo-remaining rice has high nutritional value and ensures the taste of rice. However, related biological mechanisms, such as the relationship among embryo-remaining characteristics, embryo traits, and endosperm components remains unclear. Therefore, it is necessary to further study the difference of embryo-remaining rate among different types of rice varieties and its influencing factors.【Method】Therefore, typical embryo-retaining and non-embryo-remaining varieties were selected from softand common rice as experimental materials to investigate rice embryo-remaining characteristics. Here, the embryo development characteristics of the two types of varieties were observed by slicing; then, apparent amylose content (AAC) and abscission pectin contents were compared. Finally, the differences in embryo-remaining rate, embryo traits and endosperm composition were studied under external cytokinin (CTK) application. 【Result】Compared with non-embryo-remaining varieties, embryo-remaining softvarieties and common rice had larger inner and outer embryo width, faster embryo development, large embryo volume, better grain plumpness, wider and thicker grain shape and higher abscission pectin content. After the application of CTK, the development speed of the soft rice varieties was accelerated in the middle and late stages of development, while the embryo development of the common varieties slowed down. The AAC, pectin content, embryo weight percentage and embryo-remaining rate of the two types were increased.【Conclusion】In general, apart from embryo traits, endosperm composition mainly affected the embryo-remaining characteristics of soft rice varieties, whereas grain shape was an important factor influencing the embryo-remaining characteristics of common varieties.CTK regulates the embryo-remaining rate of different types of rice varieties by influencing different traits of rice seeds. These results will lay a new theoretical basis for the breeding and cultivation regulation of high embryo-remaining rice varieties.
embryo-remaining rate; embryo development; endosperm composition; grain shape; cytokinin
10.16819/j.1001-7216.2023.220809
2022-08-25;
2022-12-26。
國家自然科學基金資助項目(32201892);遼寧省教育廳面上項目(LJKMZ20221016);遼寧省科技重大專項(2019JH1/10200001);沈陽農(nóng)業(yè)大學成果轉(zhuǎn)化項目(H2017035)。
