茶樹CsAS1和CsAS2基因的克隆及功能分析
2023-05-23 14:20:18袁連玉張麗童華榮代洪葦鄭姝婷
西南大學學報(自然科學版) 2023年5期
袁連玉, 張麗, 童華榮, 代洪葦, 鄭姝婷
西南大學 食品科學學院,重慶 400715
主持人簡介
曾亮, 西南大學涪陵研究院院長, 西南大學食品科學學院教授, 博士研究生導師. 主要從事茶及飲料植物資源功效評價與利用、 茶葉風味物質化學與感官評價等方面的研究. 主持國家級和省部級等各類項目30余項, 以第一或通信作者發表相關學術論文70余篇, 獨立譯著、 副主編和參編教材8部, 以第一發明人獲得授權國家發明專利4件; 獲國際學術“Best Researcher Award”獎1項、 獲神農中華農業科技獎三等獎、 重慶市科學技術進步獎三等獎、 中國茶葉學會“帝芙特杯”青年科技獎、 重慶市高校中青年骨干教師、 首屆全國評茶員職業技能競賽總決賽優秀裁判員、 西南大學科研工作先進個人、 優秀教師等各類榮譽; 兼任中國農村專業技術協會茶葉專委會秘書長、 重慶市茶葉學會副理事長、 重慶市營養學會副理事長、 重慶市第三屆食品安全地方標準審評委員會委員、 重慶市第一屆林業標準化技術委員會委員、 重慶市第一屆青年科技領軍人才協會會員、 精制川茶四川省重點實驗室學術委員會委員、 茶葉標準與檢測技術四川省重點實驗室學術委員會委員、 《茶葉科學》和《中國茶葉》編委等.
葉片是植物的主要營養器官, 是植物光合作用和蒸騰作用等生理代謝過程的重要場所. 葉片的發育過程是非常關鍵的植物形態建成過程, 受生理和外界環境等多種因素的綜合調控[1]. 葉片的發育過程主要包括葉片的發生、 葉片的形態特征決定和葉片的極性分化3個重要過程. 葉片的極性分化包括即背-腹軸(近-遠軸極性)、 基-頂軸和中-側軸3個方向的分化[2-3], 其中近-遠軸極性的建立能夠決定葉片背腹特征的形成, 形成葉片背側的海綿組織和腹側的柵欄組織的分化, 使得葉片由輻射對稱變成兩側對稱, 這是葉片發育的關鍵過程之一, 是保障葉片正常進行光合作用、 蒸騰作用等重要生理過程的結構基礎[4-6].
多種轉錄調控因子相互協調表達形成復雜的調控網絡, 共同調控了葉片的近-遠軸極性的建立過程, 其中HD-ZIP基因家族(包括REVOLUTA、PHAVOLUTA和PHABULOSA基因)、ARP基因家族(ASYMMETRICLEAVES1(AS1)、 玉米ROUGHSHEATH(RS2)及金魚草PHAN基因)和LOB(AS2)基因家族是主要參與葉片近軸極性建立的重要基因[7];KANADI基因家族(KAN1-4)和ARF基因家族(ARF3和ARF4)主要參與葉片的遠軸極性建成[8]; 小RNA包括的siRNAs、 ta-siRNAs和miRNAs等也可通過調控靶基因的表達參與植物葉片的近-遠軸極性的建成[9-10].ASYMMETRICLEAVES1(AS1)和ASYMMETRICLEAVES2(AS2)互為等位基因, 編碼形成的AS1與AS2蛋白均可形成轉錄因子復合體參與葉片近-遠軸極性的建成過程. AS1屬于ARP蛋白家族, 是一類MYB-R2R3型轉錄因子, 是參與DNA結合的結構蛋白[11]; AS2蛋白屬于植物特有的Lateral Organ Boundaries(LOB)蛋白家族, 在植物中廣泛存在, 能夠直接或間接參與植物生長發育及環境響應的轉錄調控過程[12-14].AS1和AS2基因的突變會導致葉片褶皺、 葉片不對稱和葉邊緣下卷等葉片缺陷的癥狀;AS1和AS2基因的超表達則會使葉片出現葉片邊緣上卷和葉片收縮等近似近軸化極性的特征[15].AS1和AS2基因家族還參與調控植物的其它發育和抗逆境脅迫等代謝調控過程, 如莖的發育、 維管束和根的發育等過程[16-17].
茶樹是我國重要的葉用經濟作物, 葉片的發育及形態建成直接影響茶葉的產量和質量, 所以研究茶樹葉片發育調控機制對提高茶樹經濟價值的重要意義. 目前, 茶樹中AS1和AS2基因還未被關注, 本研究從茶樹基因組中鑒定克隆了CsAS1和CsAS2基因, 對其進行了全面的生物信息學分析和表達分析, 可為進一步研究CsAS1和CsAS2基因家族在茶樹葉片發育過程中的重要功能提供借鑒.
1 材料與方法
1.1 材料
茶樹材料為2年生“福鼎大白茶” (Camelliasinensiscv. Fuding Dabaicha), 種植于重慶市北碚區天生路2號西南大學校內教學實驗基地. 轉錄組表達數據下載于“舒茶早”茶樹基因組數據庫TPIA(http: //tpdb.shengxin.ren/). 用于表達分析的茶樹不同組織材料, 包括成熟葉、 莖、 根及花等, 取于 “福鼎大白茶”, 液氮速凍后, 存儲于-80 ℃超低溫冰箱. 試驗試劑和材料主要包括: 植物RNA提取試劑盒(產于天根生化科技(北京)有限公司)、 反轉錄酶及配套緩沖液(產于寶生物工程(大連)有限公司)、 Taq DNA 聚合酶、 pMD18-T、 DH5α大腸桿菌感受態細胞、 DNA切膠回收試劑盒(產于上海源葉生物科技有限公司)和熒光定量PCR試劑超混液(產于重慶鼎國生物技術有限公司). 引物合成和基因測序由生工生物工程(上海)有限公司完成.
1.2 茶樹CsAS1和CsAS2基因的克隆
按照植物RNA提取試劑盒說明書的步驟, 分別提取“福鼎大白茶”茶樹的根、 莖材料的總RNA, 采用反轉錄試劑盒進行mRNA的反轉錄, 合成不同組織的cDNA, 作為基因克隆及表達分析的材料. 克隆茶樹AS1和AS2基因的PCR擴增體系為: 94 ℃, 5 min; 94 ℃, 30 s; 50 ℃, 30 s; 72 ℃, 2 min; 35個循環后, 72 ℃, 5 min, DNA純化試劑盒進行純化回收測序和分析. 所有使用的引物詳情見表1.

表1 熒光定量PCR引物信息表
1.3 茶樹CsAS1和CsAS2蛋白的生物信息學分析
分別從擬南芥數據庫TAIR和JGI-Phytozome 13 數據庫(https: //phytozome-next.jgi.doe.gov/blast-search)中下載CsAS基因在9種不同植物中的核苷酸和蛋白質序列, 利用TPIA數據庫工具進行Blast搜索, 鑒定茶樹CsAS基因. 并進行了編碼蛋白的分子量、 等電點的蛋白性質分析; 分別采用SOPM和SWISS-MODEL軟件分析了CsAS1和CsAS2蛋白的結構; 用Clustal X1.8, MEGA 4.0和DNAMAN等生物學軟件分析了茶樹CsAS蛋白的氨基酸序列, 采用鄰接法(Neighbor-joining)方法構建進化樹, Bootstrap參數為1 000, 其余參數設置為默認值; 分別通過MEME、 Plant-mPLoc和Plantcare軟件分析了茶樹CsAS蛋白的保守基序(基序數量設為5)、 亞細胞定位和CsAS基因上游2 kb啟動子序列中含有的順式作用元件; 利用模式植物擬南芥ATAS蛋白的相互作用網絡, 在數據庫STRING中建立了茶樹CsAS蛋白的相互作用關系網絡.
1.4 茶樹AS受體蛋白基因的表達分析
本研究從TPIA數據庫中下載了茶樹葉片中CsAS基因在鹽、 冷、 干旱和MeJA等不同非生物逆境脅迫條件下的轉錄表達水平的數據, 并利用Tbtools繪制熱圖. 為了驗證數據庫中的基因表達量, 本研究還采用熒光定量PCR的方法檢測CsAS基因在茶樹5個組織部位(花、 芽、 成熟葉、 莖、 根)、 不同發育時期的芽、 不同葉位的葉片及在外源GA3、 IAA處理條件下的表達量. 本研究采用水培的方式對2年生“福鼎大白茶”扦插苗進行外源激素脅迫處理, 分析了CsAS基因的表達特異性, 每盆24株, 在盆中加入配制好的響應的營養液, 加氣泵通入氧氣, 每7天更換一次營養液, MS培養30天后進行MS/(100 mg/L)GA3和MS/(50 mg/L)IAA脅迫處理, 分別在24 h和48 h后取樣, 液氮固樣, -80 ℃保存. 采用艾德萊植物RNA提取試劑盒進行樣本的總RNA的提取, 并反轉錄成cDNA 模板, 用于CsAS基因表達特異性分析. 用Primer 3軟件設計熒光定量引物(表1), 內標基因為茶樹肌動蛋白基因Actin1. 10 μL Real-Time PCR 反應體系為: SsoFast EvaGreen Supermix 5 μL, 濃度為10 μmol/L的上下游引物各 0.20 μL, cDNA 1 μL, ddH2O 3.5 μL, 充分混勻, 于Bio-Rad CFX 96實時定量PCR儀上進行擴增分析, 反應程序為95 ℃, 10 s; 95 ℃, 5 s; 55 ℃, 5 s, 進行40個循環. 每個樣品進行3次生物學重復和3次試驗重復, 采用2-ΔΔCT法分析結果, 分別使用SPSS和ORIGIN軟件進行差異顯著性分析和作圖.
2 結果與分析
2.1 茶樹AS蛋白基因的鑒定及克隆
本研究利用模式植物中AS蛋白的核苷酸和氨基酸序列信息, 在茶樹基因組數據中進行本地Blast搜索鑒定, 共獲得茶樹中同源的AS1和AS2蛋白基因各1個:CsAS1和CsAS2. 如圖1, 本研究分別以“福鼎大白茶”茶樹莖和根為材料獲得的cDNA為模板, 克隆獲得了CsAS1和CsAS2基因, 長度分別為1 033 bp和688 bp. 如表2,CsAS1和CsAS2基因分別編碼344 aa和229 aa長度的氨基酸殘基, 分子量分別為39.45和25.33; 蛋白的等電點分別為10.04和7.36; 亞細胞定位預測分析顯示2個AS蛋白均定位于細胞核, 這與其它植物中的AS蛋白的定位模式一致. 如圖2, 染色體定位分析結果顯示2個CsAS基因分別定位于茶樹的4號和10號染色體上.
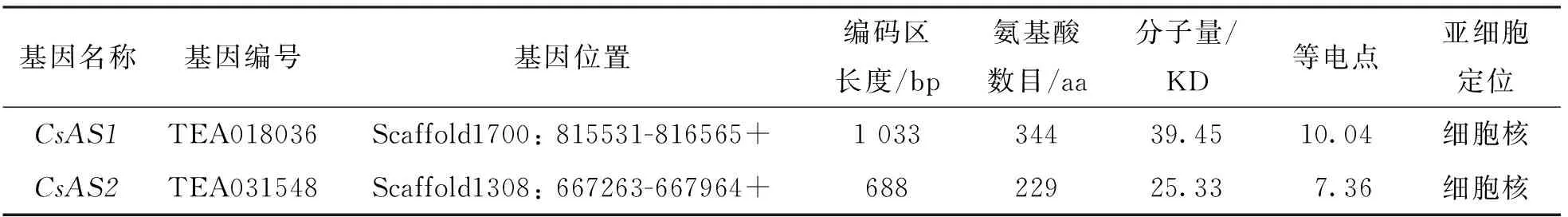
表2 茶樹CsAS1和CsAS2蛋白的理化性質
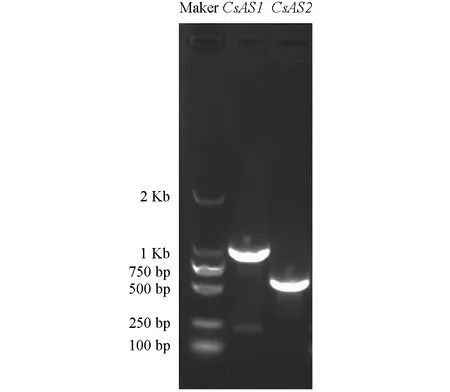
圖1 CsAS1和CsAS2基因的PCR擴增
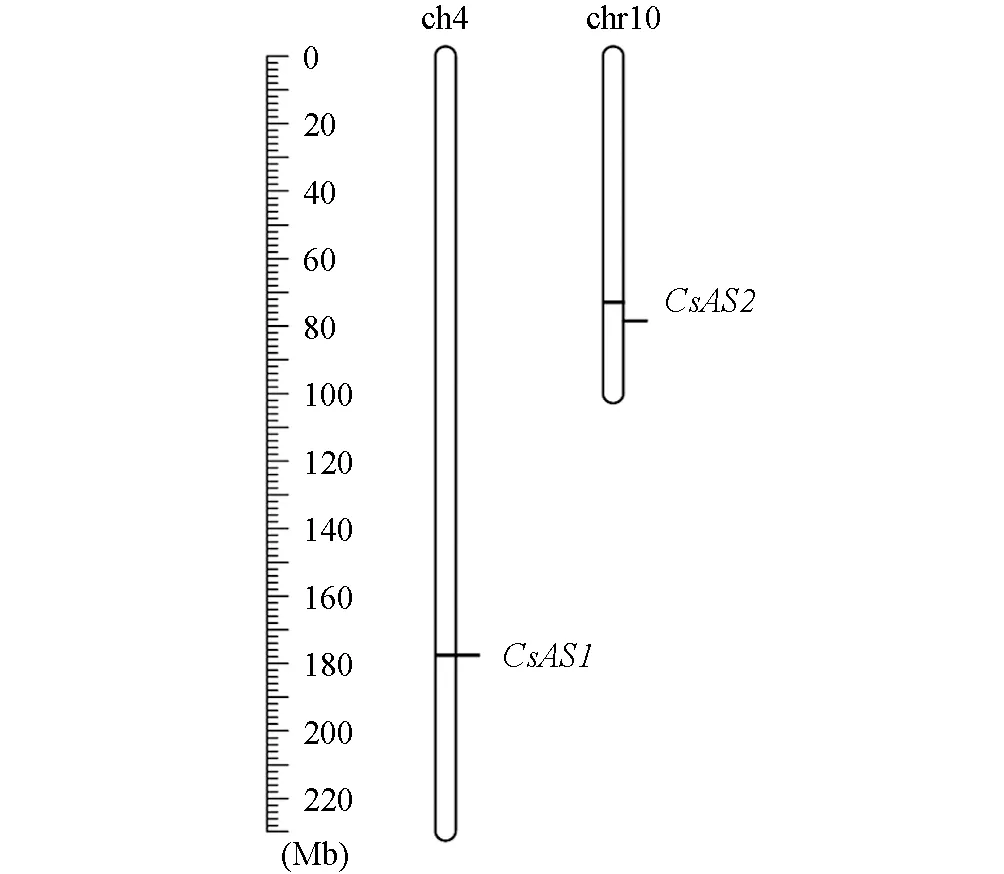
圖2 CsAS1和CsAS2基因的染色體定位
2.2 茶樹CsAS1和CsAS2蛋白系統進化及蛋白保守結構域分析
系統進化樹分析結果顯示(圖3), 茶樹CsAS1和CsAS2蛋白與其它植物中的該類蛋白氨基酸序列具有高度的保守性, 茶樹CsAS1蛋白與葡萄VitAS蛋白的親緣關系最近, 而CsAS2與水稻OsAS蛋白的親緣關系最近. 保守基序分析顯示(圖4a), AS1和AS2蛋白在不同植物中均具有高度的保守性, 不同植物CsAS1和CsAS2蛋白序列中保守基序的種類和數量也高度一致, 但CsAS1和CsAS2蛋白之間差異比較明顯, 分屬于不同的亞家族. 基因結構分析顯示(圖4b),CsAS1基因只有1個外顯子,CsAS2基因含有2個外顯子和1個內含子. CsAS1蛋白屬于ARP蛋白家族, CsAS2蛋白屬于LOB蛋白家族. 多種植物的AS1和AS2蛋白的多序列比對結果顯示(圖5), 不同植物中AS1蛋白序列一致性為64.91%, AS2蛋白的一致性為53.69%; CsAS1蛋白氨基酸序列中含有2個保守的DNA結合結構域: DNA結構域Ⅰ和Ⅱ; CsAS2蛋白氨基酸序列中含有LOB家族蛋白所共有的半胱氨酸結構域CX2CX6CX3C(ZF基序)、 GAS(Gly-Ala-Ser)、 負責蛋白質二聚體的類亮氨酸拉鏈的螺旋基序LX6LX3LX6L、 ICG和LZL等特殊結構域, 均與該蛋白家族的功能有直接關系. 茶樹的CsAS1和CsAS2蛋白均為典型的AS蛋白, 推測可能與已經報道的其它植物AS蛋白具有相似生物學功能.
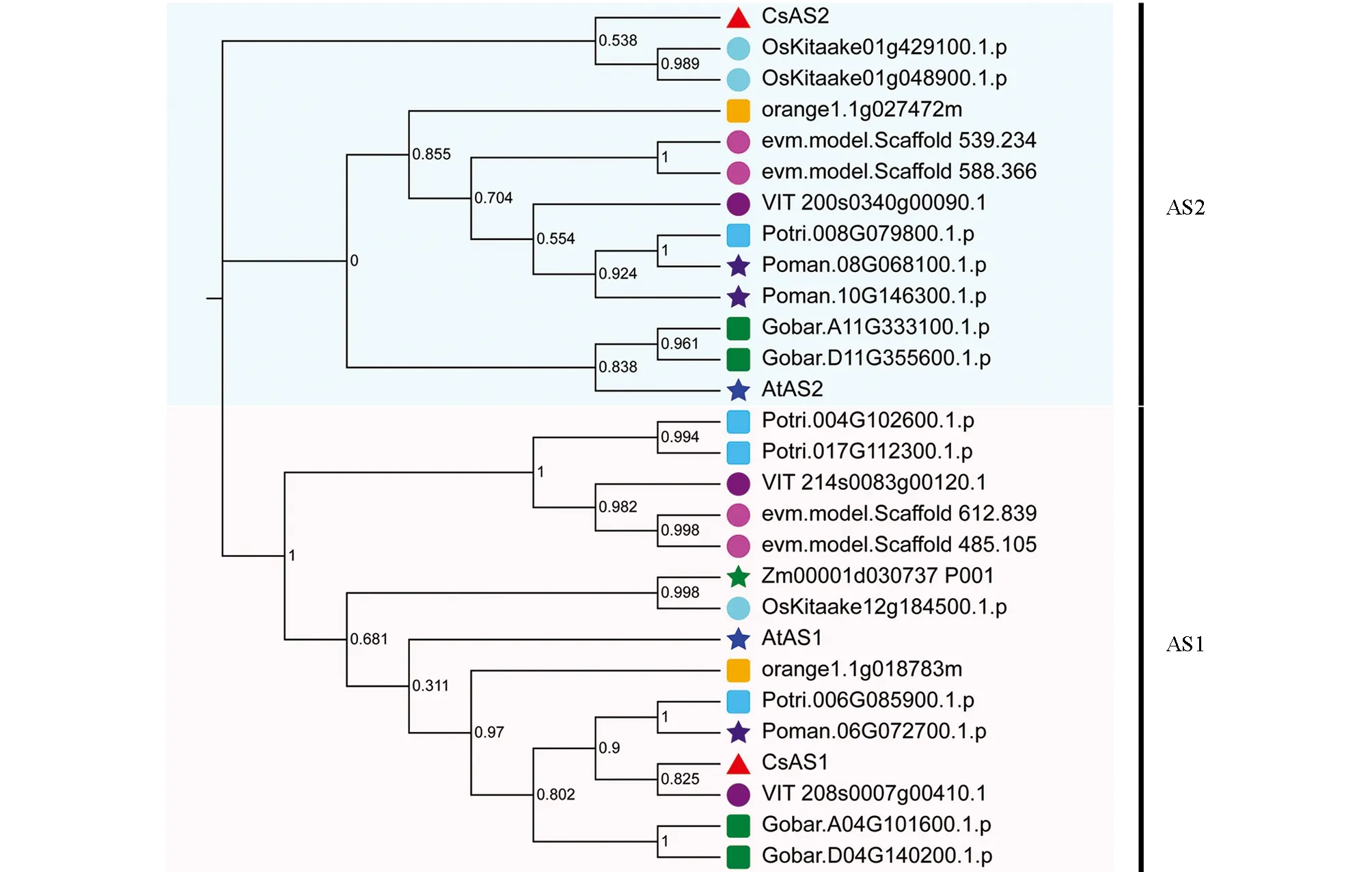
At為擬南芥, Os為水稻, Zm為玉米, VIT為葡萄, Potri為楊樹, Poman為小黑樹, Gobar為陸地棉, orange為柑橘, evm為咖啡, CsAS為茶樹.圖3 植物中AS蛋白的進化樹分析
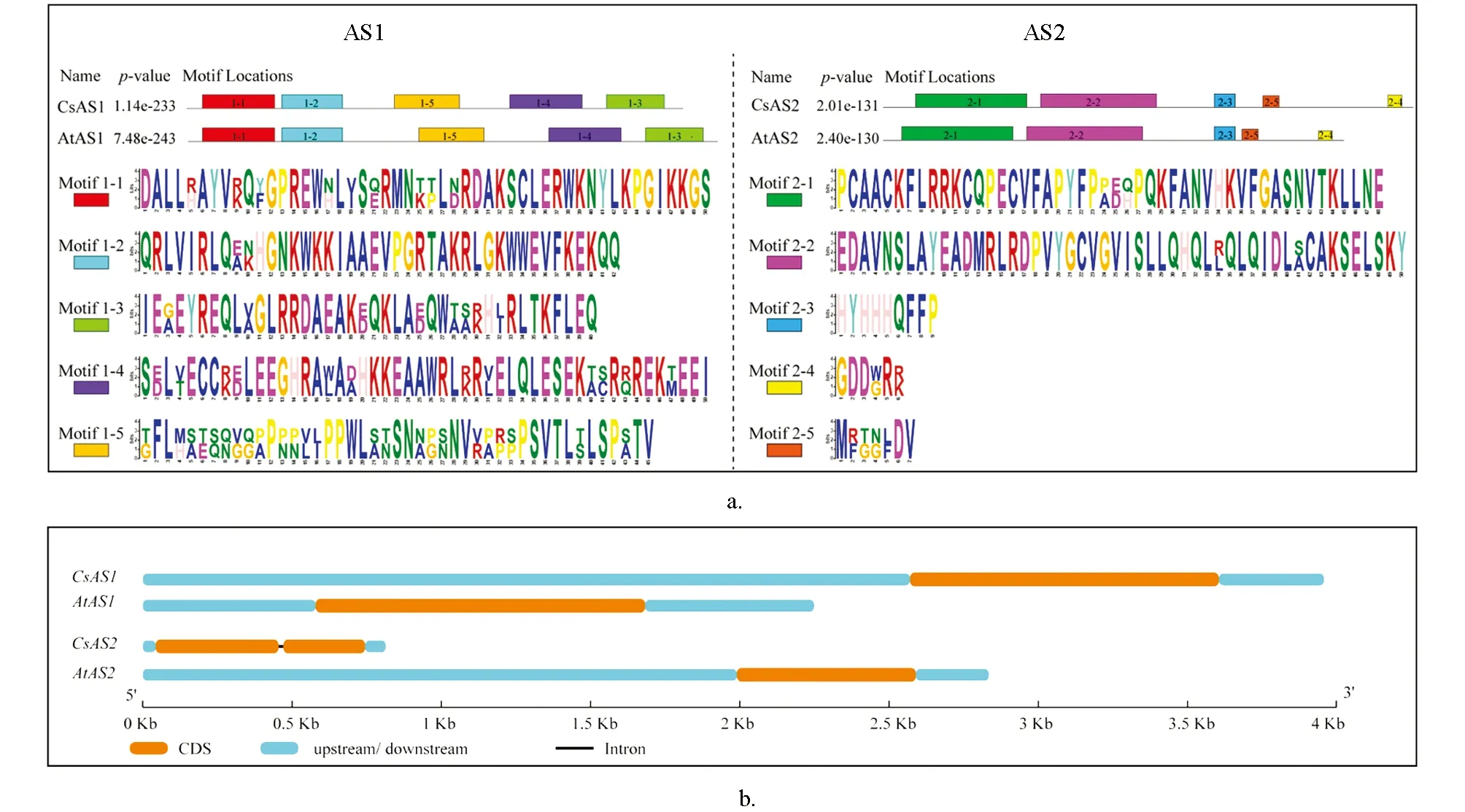
(a) AS蛋白保守基序分析, 不同顏色的方塊和數字代表不同的保守基序, 不同顏色的字母代表保守基序的氨基酸序列信息. (b) AS基因結構分析. AtAS為擬南芥AS蛋白, CsAS為茶樹AS蛋白.圖4 植物中AS蛋白家族的保守基序分析及CsAS基因結構分析

At為擬南芥, Os為水稻, VIT為葡萄, Cs為茶樹.圖5 茶樹CsAS與其它植物AS氨基酸序列比對
2.3 茶樹CsAS1和CsAS2蛋白的二、 三級結構分析
蛋白空間結構分析顯示(圖6a), 茶樹CsAS1蛋白中α-螺旋所占的比例最大, 為56.69%, 其次是自由卷曲, 為28.78%, 延長鏈和β-轉角所占的比例小; CsAS2蛋白中α-螺旋和自由卷曲所占的比例均較大, 分別為36.68%和36.24%, 延長鏈和β-轉角所占的比例較小. 分別以已知蛋白MYB21和LOB蛋白為模板, 用SWISS-MODEL軟件構建的蛋白三級結構顯示, CsAS1和CsAS2蛋白中均含有大量的α-螺旋和自由卷曲, 但三級空間結構不同, 差異較大, 均與同源的擬南芥AS蛋白結構相似. 由此可推測CsAS蛋白與模式植物中的AS蛋白具有相同的生物學功能.
2.4 茶樹CsAS1和CsAS2基因的時空表達特異性分析
為明確CsAS1和CsAS2蛋白在茶樹葉片和其它器官生長發育中的調控功能, 本研究利用Real-time PCR的方法分析了CsAS1和CsAS2基因在茶樹根、 莖、 葉、 花和芽等不同組織部位的表達特異性.
如圖7a,CsAS1基因在茶樹不同組織中均有表達, 但在芽中的表達量遠遠高于其它組織; 而CsAS2基因在茶樹不同組織中均有表達, 但在根中的表達量遠遠高于其它組織. 如圖7b,CsAS1和CsAS2基因在芽的不同發育時期中隨著成熟度的增加均有先增加再減少的趨勢. 如圖7c,CsAS1基因在葉片不同發育時期中隨著成熟度的增加而減少, 在幼嫩芽葉中的表達高于第一、 二和三葉;CsAS2基因在葉片不同時期發育先降低, 再恢復到在芽中的表達水平, 在幼嫩芽葉和第三葉中的表達高于第一、 二和三葉. 由此推測茶樹CsAS1和CsAS2基因在茶樹不同組織和葉片不同發育階段發揮著不同的功能.
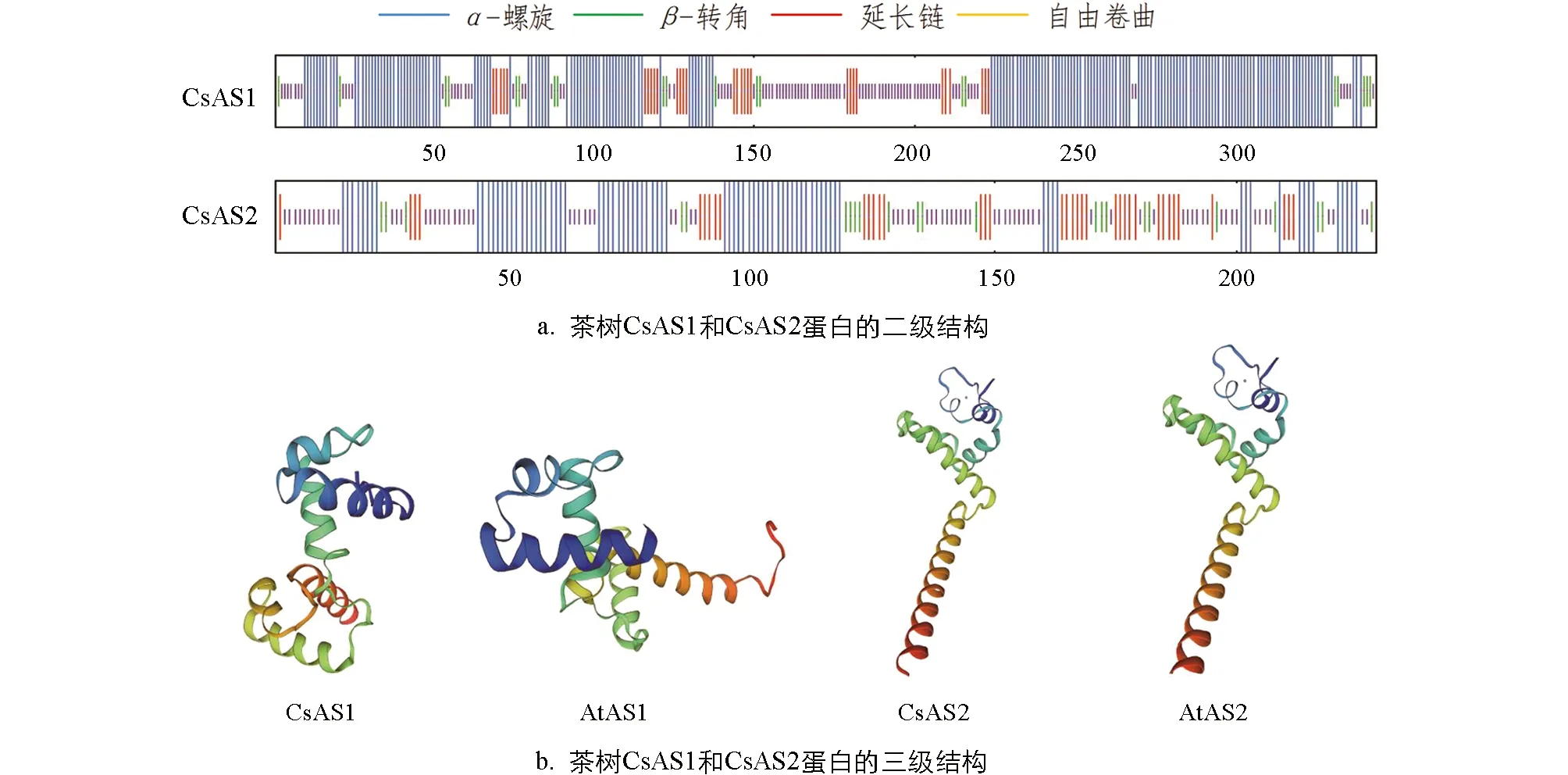
圖6 茶樹CsAS1和CsAS2蛋白結構分析
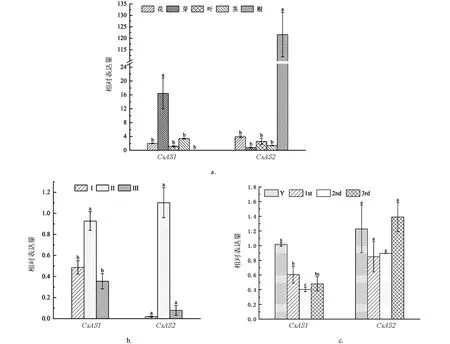
(a) CsAS1和CsAS2基因在茶樹不同組織的表達分析. (b) CsAS1和CsAS2基因在茶樹芽不同發育時期的表達分析. (c) CsAS1和CsAS2基因在茶樹不同葉位的表達分析. 小寫字母不同表示p<0.05, 差異具有統計學意義.圖7 CsAS1和CsAS2基因的表達分析
2.5 茶樹CsAS1和CsAS2基因啟動子片段順式作用元件分析
為分析茶樹CsAS1和CsAS2基因的潛在功能, 通過Plant Care軟件分析了CsAS1和CsAS2基因的啟動子序列. 結果如圖8所示,CsAS1和CsAS2基因啟動子序列中均含有多個順反子元件, 主要包括脅迫響應元件(MYB、 MYC)、 光響應元件、 植物激素響應元件和發育調控元件. 其中脅迫響應元件占比比較大, 植物激素響應元件也有較多存在. 由以推測CsAS1和CsAS2基因可參與茶樹對非生物逆境和植物激素的響應過程. 為了驗證這兩個基因能夠參與植物激素的調控過程, 本研究分析了CsAS1和CsAS2基因在外源激素脅迫條件下的表達特性, 如圖8d所示, 在50 mg/L IAA處理24 h和48 h后,CsAS1基因受到誘導, 有較大程度的表達量增加,CsAS2基因在24 h時減少, 48 h時增加; 在100 mg/L GA3處理24 h和48 h后,CsAS1和CsAS2基因均受到誘導, 但CsAS1增加的表達量相對比較少,CsAS2基因的表達被明顯誘導, 增加量接近50倍.
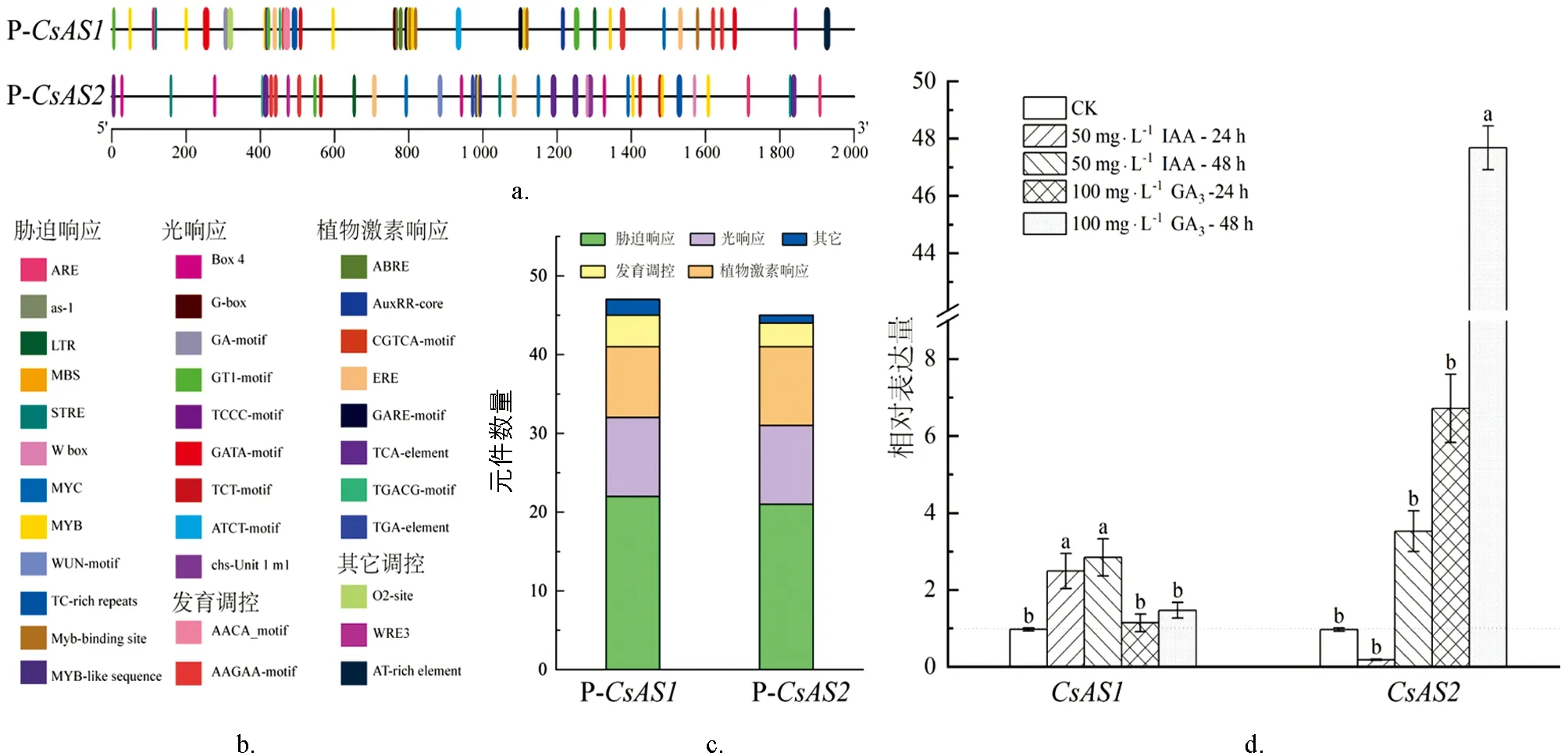
(a-c) 啟動子元件分析. (d) CsAS1和CsAS2基因在不同外源GA3、 IAA激素處理脅迫下的表達分析. 小寫字母不同表示p<0.05, 差異有統計學意義.圖8 茶樹CsAS1和CsAS2基因啟動子片段順式作用元件分析
2.6 茶樹CsAS1和CsAS2基因響應非生物逆境脅迫的表達特異性分析
本研究利用TPIA已經公布的轉錄組數據分析了CsAS1和CsAS2基因在NaCl、 干旱、 MeJA和冷等不同非生物逆境脅迫條件下的表達分析. 如圖9所示,CsAS1基因可以被NaCl、 干旱、 MeJA和冷脅迫誘導表達, 其中被冷馴化后表達上調, 而在其余3種脅迫后表達下調;CsAS2基因僅在干旱處理72 h和NaCl處理24 h后被誘導, 且表達上調. 由以上結果可以推測, 茶樹CsAS1和CsAS2基因可能參與茶樹應對環境中鹽、 干旱和冷等非生物逆境脅迫的響應過程.
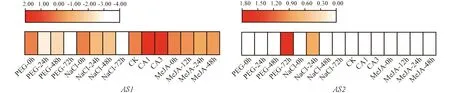
利用PEG模擬干旱脅迫; 圖例中正數表示上調表達, 負數表示下調表達; 圖下方為不同脅迫處理及其對照.圖9 茶樹CsAS1和CsAS2基因在不同逆境脅迫下的表達分析
2.7 茶樹CsAS1和CsAS2的蛋白關聯蛋白分析
利用擬南芥同源的AtAS蛋白進行了CsAS蛋白的關聯蛋白預測分析, 如圖10所示, 茶樹CsAS1和CsAS2的蛋白與多種葉片形態建成相關的蛋白間均存在關聯性, 其中包含KNAT1(knotted1-like homeobox protein), KNAT2(knotted2-like homeobox protein), KAN(Homeodomain-like superfamily protein), LOB(Lateral organ boundaries domain family protein), TCP14(TEOSINTE BRANCHED、 cycloidea and PCF 14), TCP15(TEOSINTE BRANCHED、 cycloidea and PCF 15), AFO(Plant-specific transcription factor YABBY family protein)和HDA6(Histone deacetylase 6)等蛋白. 由此可推測, CsAS1和CsAS2蛋白是可以參與茶樹葉片形態建成過程的.
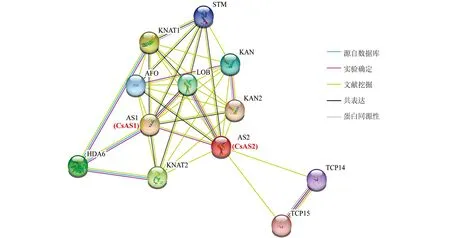
圖10 茶樹CsAS1和CsAS2相互作用蛋白網絡分析
3 討論
茶樹是葉用經濟作物, 其葉片的形態建成也存在形態的多樣性, 例如, 葉片的大小、 厚度和顏色等. 其中茶樹葉片的內卷和外卷等與葉片極性建立相關的現象同樣也經常被觀察到. 但目前茶樹分子生物學研究的重點依舊在葉片的次生代謝和顏色調控上, 對于葉片發育的關注度還不夠. 眾多轉錄因子參與了葉片發育和形態建成過程[18, 19]. AS蛋白已經在多種植物中被報道參與葉片的近-遠軸極性建立過程,AS基因的變異或缺失會導致葉片的卷曲、 外翻或者內卷等葉片去極性特征[8, 20-21]. 本研究從“福鼎大白茶”茶樹中克隆獲得了2個與葉片發育有著緊密相關性的基因:CsAS1和CsAS2, 其分子量、 等電點和編碼區長度與已經報道的AS蛋白具有高度的同源性, 且均定位于細胞核(表2). 茶樹CsAS1和CsAS2的氨基酸序列與水稻和葡萄等植物的該類蛋白高度相似, 分別含有植物ARP蛋白的保守結構域(DNA結合結構域Ⅰ和Ⅱ)和LOB家族蛋白的保守結構域(半胱氨酸結構域CX2CX6CX3C即ZF基序, GAS, ICG和LZL區域, 負責蛋白質二聚體的類亮氨酸拉鏈的螺旋基序LX6LX3LX6L)等特殊功能結構域. 其中, ZF基序是AS2蛋白結合DNA活性所必需的, ICG和LZL區域是AS2核定位的必要區域[22-24], 說明茶樹CsAS1和CsAS2蛋白具備發揮該類蛋白功能的基礎序列元件.CsAS1和CsAS2基因在葉片不同發育時期中隨著成熟度的增加而減少, 在幼嫩芽葉中的表達量高于第一、 二和三葉;CsAS1和CsAS2基因在芽的不同發育時期中隨著成熟度的增加均有先增加后減少的趨勢. 以上結果可說明CsAS1和CsAS2蛋白具備其它植物中報道的AS1和AS2蛋白的相似功能, 可參與葉片發育中的近-遠軸極性建立的調控.
AS轉錄因子蛋白家族還被報道廣泛參與調控植物除葉片外的其它組織器官的發育和非生物逆境脅迫響應等調控過程, 例如擬南芥的AS1蛋白可以負調控ABA代謝途徑[25, 26], 馬鈴薯AS2蛋白有利于提高其應對高鹽、 干旱、 高溫、 低溫等非生物脅迫的能力[16]. 啟動子順式作用元件和表達分析發現, 茶樹CsAS1和CsAS2基因的啟動子序列中均含有多個逆境脅迫響應元件, 茶樹CsAS1基因的表達會被NaCl、 MeJA和干旱脅迫抑制, 可能存在負調控關系; 在冷脅迫條件下,CsAS1基因的表達被上調, 是正調控的關系;CsAS2基因的表達可被干旱脅迫和NaCl處理誘導. 茶樹CsAS2基因在根中的表達量遠高于其它組織, 與玉米LOB家族蛋白RTCS和RTCL基因類似, 可能能夠參與側根及卷須根的形成調控過程[27].
AS蛋白發揮生長發育調控功能是通過與其它蛋白形成復雜的網絡而完成的. 茶樹CsAS1和CsAS2的蛋白可與多種葉片形態建成相關的蛋白間存在關聯性, 通過與KNAT1、 KNAT2和KAN蛋白的相互作用參與葉片的起始過程[28], 和LOB蛋白家族中的其它成員相互作用, 參與葉片的極性建立過程, 參與TCP14、 TCP15、 AFO和HDA6等共同調控葉片及其它器官的發育過程[29-31]. 后續本研究團隊將以CsAS1和CsAS2蛋白為中心, 通過蛋白互作網絡的研究, 解析茶樹葉片等重要組織器官的發育過程, 探究CsAS1和CsAS2蛋白在茶樹生命周期不同發育階段的詳細功能.
4 結論
本論文從“福鼎大白茶”茶樹莖和根中鑒定和克隆獲得茶樹CsAS1和CsAS2基因. 生物信息學分析顯示, 茶樹CsAS1和CsAS2蛋白具有高度的保守性;CsAS1和CsAS2基因的表達在茶樹不同組織和不同發育階段存在時空特異性, 且受到外源植物激素IAA和GA3及其它非生物逆境的影響. 根據本研究結果推斷CsAS1和CsAS2基因可廣泛參與茶樹葉片的形態建成和非生物逆境的脅迫響應過程, 為深入分析茶樹CsAS1和CsAS2蛋白的功能提供了一定參考.
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
