管氏腫腿蜂密度對其寄生及繁衍能力的影響
2023-05-27 14:00:56牛國霜
環境昆蟲學報 2023年2期
牛國霜,李 莉*,唐 明,2
(1. 貴州師范大學生命科學學院,貴陽 550025;2. 貴州師范大學西南喀斯特山地生物多樣性保護國家林草局重點實驗室,貴陽 550025)
管氏腫腿蜂SclerodermusguaniXiaoetWu隸屬于膜翅目Hymenoptera腫腿蜂科Bethylidae硬皮腫腿蜂屬Sclerodermus,因其寄主廣泛、生活史短、繁殖速度快、生態適應性強和搜索寄主能力強等習性,在我國廣泛用于林木蛀干害蟲的生物防治(陳君和程惠珍, 2000; 來燕學等, 2012; 黃維亞, 2017; 楊忠岐, 2018)。寄生蜂不僅要提高寄主搜索效率,盡快找到合適的寄主產卵,還要應對天敵或微生物的威脅,才能在復雜的棲息環境中生存(代平禮, 2005; 黃維亞, 2017; 郭涵等, 2020; 楊春平等, 2021)。作為一種亞社會性昆蟲,其親代雌成蜂為降低單獨處理寄主帶來的死亡風險,提高其寄生成功率及繁殖力,會采取協同覓食、共同克服寄主抗性和抵御天敵侵襲等策略以面對不利生存環境(Tallamy, 2000, 2001; 尚玉昌, 2008; Miller and Zink, 2012; 黃維亞, 2017; Liuetal., 2021)。
另外,管氏腫腿蜂也是典型的卵育型(Synovigenic)抑性寄生蜂(Idiobiont),雌蜂羽化時雖達到性成熟,但體內無成熟卵粒,直到克服寄主、獲得充足營養后,卵巢才會進一步發育(胡尊瑞等, 2013; 蔣玉潔, 2021)。很大程度上,這種繁殖前提取決于雌蜂處理或制服寄主達到可寄生狀態的效率(即通過雌蜂向寄主體內注入毒素將其麻痹致死,永久性削弱寄主活動力,并抑制其生長發育)(周冰穎, 2017; 楊忠岐, 2018)。當寄主被制服后,雌蜂則在較低寄主抗性風險下取食寄主血淋巴,腹部逐漸膨大,卵巢內形成成熟卵,隨后產出(周冰穎, 2017; 蔣玉潔, 2021)。其中,寄主抗性與其體型大小有關,個體越大對寄生蜂釋放毒素的耐受性就越強(辛蓓等, 2016; 任雪敏等, 2018),需要多次注入毒素或多頭協作蜇刺方可增加寄生成功率,才能保障子代的存活和發育(Tangetal., 2014)。例如,川硬皮腫腿蜂Sclerodermussichuanensis等腫腿蜂屬昆蟲往往會以多頭雌蜂共同進攻、克服單一寄主并合作撫育子代(趙琪等, 2020),提高寄生率和增加子代數量(王雪菲等, 2020)。有研究表明,多頭雌蜂協作寄生的情況會縮短其產卵前期(林芳芳等, 2015)和子代發育歷期(唐艷龍等, 2020)。因此,寄生過程中寄生蜂的數量或密度與其寄主利用效率密切相關。在同一寄主上,雌蜂密度大小與其繁殖利益之間可能會存在矛盾或沖突。比如,在相同大小寄主上,多頭雌蜂所產生平均后代數量少于單頭雌蜂寄生的情況(唐秀云, 2015)。那么,寄生蜂在應對有限寄主資源時,如何解決寄生密度與繁殖最大化之間的矛盾呢?
為了明確不同雌蜂密度對寄主的寄生效能(Parasitism efficency),以及對隨后子代存活和發育的影響,本研究以“管氏腫腿蜂-松墨天牛”為研究對象,引入Hassell-Varley數據模型分析尋找效應(E)與天敵自身密度(P)的相互干擾關系,以探究雌蜂密度對管氏腫腿蜂寄主利用的適應性策略,主要包括:8種不同雌蜂密度(1~8頭/寄主)下,雌蜂克服寄主的行為和效能,以及雌蜂生殖力和子代適合度變化,擬為亞社會性寄生蜂多頭協作寄生和撫育子代的生殖策略提供重要科學依據。
1 材料與方法
1.1 供試蟲源
選取健康的松墨天牛M.alternatus3~4齡(0.350~0.450 g)幼蟲作為寄生蜂室內繁育寄主。該供試寄主購于貴州省黔東南自治州凱里市(東經107.981°,北緯26.566°),以單頭、單管分裝在放有木屑的試管中,并冷藏、保存于4~5℃冰箱中待用。
管氏腫腿蜂來源于貴州師范大學生命科學學院“有害生物綜合防控及資源化利用”實驗室人工飼養。以上述寄主幼蟲續代飼養(>40代),獲得穩定的實驗種群。光照培養箱飼養條件為溫度26℃±0.5℃,相對濕度70%±5%,光周期L∶D=10∶14。
1.2 儀器和試劑
光照培養箱(RXZ智能型,寧波江南儀器廠);冰箱(BCD-290W;青島海爾股份有限公司),滅菌鍋(BXM-30R;上海博迅實業有限公司醫療設備廠),鼓風干燥箱(JC101;上海成順儀器儀表有限公司),體式解剖鏡(OLYMPUS SZX7),電子天平(HZT-A500,美國康州HZ電子有限公司),玻璃指型管,毛筆,鑷子,醫用脫脂棉,計數器,蒸餾水,75%酒精(貴州利健消毒制品有限公司)。
1.3 比較不同雌蜂密度下管氏腫腿蜂的寄生行為
選取3~4齡健康松墨天牛幼蟲(體重為0.350~0.450 g)供試。首先對寄主體表進行消毒處理,稱重并記錄單頭寄主體重后裝入玻璃管(?:12 mm,L:75 mm)中。選取初羽化5~6 d已交配的雌成蜂,分別按照1~8頭/寄主接蜂,每雌蜂密度20個重復,在體視顯微鏡下觀察雌成蜂克服寄主的過程。從接入寄生蜂開始,實時觀察到雌成蜂首次蟄刺寄主后,每天早、晚8點各觀察1次寄主麻痹情況及雌蜂產卵情況,此后每天早上8點觀察一次卵的孵化情況。依據卓志航等(2013),寄主完全麻痹的判斷標準為在體視顯微鏡下觀察寄主對刺激(用軟毛刷輕輕刷動寄主頭部)完全沒有反應,完全僵化,沒有任何動作。觀察記錄生物學指標包括:蟄刺前歷期(h)、寄主完全麻痹時長(h)和產卵前期(d)。其中,管氏腫腿蜂蟄刺前歷期(h),指從接入雌蜂開始到雌蜂首次蟄刺寄主為止的時間(Gaoetal., 2016);寄主麻痹時長(h),指從雌蜂第一次蟄刺寄主到寄主完全不動為止(h);產卵前期(d)指從接入雌蜂到觀察到寄主上有第一粒卵為止。其中有腫腿蜂被天牛咬死的重復去除不用。
1.4 不同雌蜂密度下寄主致死作用的尋找效應
尋找效應指天敵在寄生或捕食過程中對于獵物攻擊的行為效應(張世澤等, 2005)。管氏腫腿蜂對寄主的尋找效應參考捕食性天敵尋找效應的計算方法,按照以下公式計算(王小藝等, 2010):
(1)
式中,E為尋找效應,Na為被致死的寄主數(指已麻痹且被產卵寄主數)(展茂魁, 2014),N為寄主密度,P為天敵密度。
在一定空間內天敵自身密度的干擾作用會降低其天敵尋找效應(陳福壽等, 2006)。尋找效應(E)與天敵自身密度(P)的相互干擾關系可用Hassell-Varley模型來估計,即:
E=Q×P-m
(2)
其中,Q、m分別為搜索參數(即在無競爭條件下天敵尋找效應)和相互干擾參數(天敵種內競爭的估計)。要估計Q和m參數,可參照陳福壽等(2006)將其線性化為下面方程:
LnE=LnQ+(-m)×LnP
令Y=LnE,X=LnP,A=LnQ,B=-m;上式轉化為Y=A+BX,通過線性最小二乘法估算A、B的值,通過試驗在一定寄主密度下,管氏腫腿蜂對松墨天牛幼蟲的寄生量,從而得出參數Q和m。由此得出管氏腫腿蜂自身密度對寄主致死尋找效應的影響。
1.5 雌蜂密度對雌蜂生殖力的影響
根據1.3的試驗處理,寄主被麻痹產卵后,觀察記錄雌蜂生殖力指標:總產卵量按照寄主體上管氏腫腿蜂子代總數不再增加時的數量(粒)進行統計;單雌平均產卵量(粒,產卵總數/母蜂數)、單雌產雌量(頭,羽化雌蜂數/母蜂數)。
1.6 不同雌蜂密度下子代適合度的比較
根據1.3的試驗處理,比較分析的子代適合度指標包括:子代平均發育歷期、子代存活率、子代雄性比及單雌體重。為更好地觀察子代發育歷期,根據伍紹龍等(2013)對管氏腫腿蜂子代不成熟時期的劃分方法,將其分為6個時期,即卵期(Egg stage)、低齡幼蟲期(Early instar larva)、高齡幼蟲期(Late instar larva)、老熟幼蟲期(Mature larva)、吐絲幼蟲期(Spinning mature larva)和繭蛹期(Pupa cocoon)。卵歷期指從觀察到管氏腫腿蜂雌蜂開始產第一粒卵到第一頭低齡幼蟲出現的時間(d);幼蟲歷期指從第一頭子代進入幼蟲階段到化蛹的時間(d);雌、雄蜂蛹歷期分別指從子代進入蛹期到雌、雄蜂羽化的時間(d);卵存活率(%)=(存活卵數量/總的產卵數量)×100;幼蟲存活率(%)=(存活幼蟲數量/總的產卵數量)×100(黃維亞, 2017);子代蛹存活率(%)=(羽化子代數量/總的產卵數量)×100;性比(%)=(子代雄蜂數/子代成蜂總數)×100;單雌體重(mg)=子代雌成蜂重量/子代雌成蜂的總數量。
1.7 數據分析
運用Excel 2021.0整理數據,使用SPSS 25.0(SPSS Inc.‘Chicago’IL’USA)進行統計分析。百分數數據(子代存活率、性比)先進行反正弦平方根轉換,然后利用K-S方法檢驗數據是否符合正態分布;對符合正態分布的數據采用單因素方差分析(ANOVA)進行統計分析,運用(least significant difference,LSD)最小顯著差法比較不同雌蜂密度下雌蜂蟄刺前歷期、寄主麻痹時長、產卵前期、雌蜂產卵量、單雌產卵量、單雌產雌量、卵、幼蟲、蛹的平均發育歷期和存活率以及子代性比和單雌體重之間的差異;對不符合正態分布或方差齊性的數據采用非參數分析,并用Kruskal-WalliH進行檢驗。對不同雌蜂密度下的蟄刺前歷期、寄主麻痹時長、產卵前期、雌蜂產卵量、單雌產卵量和單雌產雌量進行斯皮爾曼(Spearman)相關性分析。致死效應數學模型用DPS軟件進行參數估計和方程的顯著性檢驗。使用Origin 2021軟件進行繪圖。
2 結果與分析
2.1 比較不同雌蜂密度下管氏腫腿蜂的寄生行為
不同雌蜂密度下管氏腫腿蜂的蟄刺前歷期不同(n=159,H=66.646,P<0.01)(圖1)。當雌蜂密度僅為1頭/寄主時,蟄刺前歷期最長,約為14.964±2.830 h(mean±SE,下同),雌蜂密度增加到7頭/寄主時,其蟄刺前歷期最短,平均只有0.213±0.070 h,僅為雌蜂密度為1頭/寄主時的1/70。隨著寄生蜂密度增加,蟄刺前歷期逐漸縮短,當雌蜂密度為2~4頭/寄主時沒有顯著差異(n=59,H=3.530,P=0.171);且當雌蜂密度增加到6頭/寄主以上時,蟄刺前歷期不再有顯著差異(n=60,H=2.208,P=0.332)。
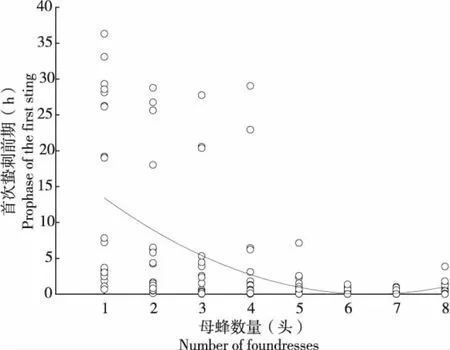
圖1 雌蜂密度對蟄刺前歷期的影響Fig.1 Effect of duration before the first stinging of Sclerodermus guani in different parasitoid density
管氏腫腿蜂完全麻痹寄主的時間隨雌蜂密度的增加而降低(n=140,H=47.317,P<0.01)(圖2)。通常,即使雌蜂密度最低時也能使寄主完全麻痹,但寄主麻痹時間長高達64.888±5.434 h,而當雌蜂密度為8頭/寄主時,寄主麻痹時長最短,僅需要20.649±3.60 h。當雌蜂密度增加到5頭/寄主時,寄主麻痹時長沒有顯著差異(n=70,H=5.558,P=0.135)。
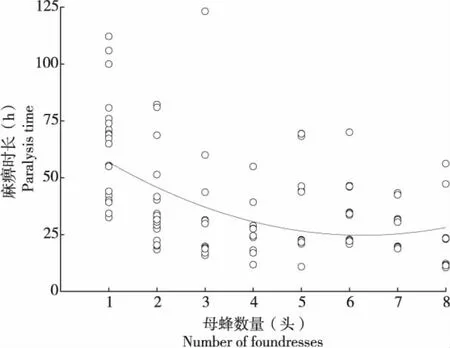
圖2 不同雌蜂密度下的寄主麻痹時長Fig.2 Host paralysis duration of Sclerodermus guani in different parasitoid density
雌蜂密度對管氏腫腿蜂的產卵前期具有極顯著的影響(n=134,H=100.969,P<0.01)(圖3)。當雌蜂密度為1頭/寄主時,雌蜂產卵前期最長,平均為6.264±0.192 d,雌蜂密度為7頭/寄主時,雌蜂產卵前期最短,僅為3.261±0.237 d。隨著雌蜂密度增加,管氏腫腿蜂的產卵前期縮短,當雌蜂密度為2~6頭/寄主時,雌蜂產卵前期沒有顯著差異。
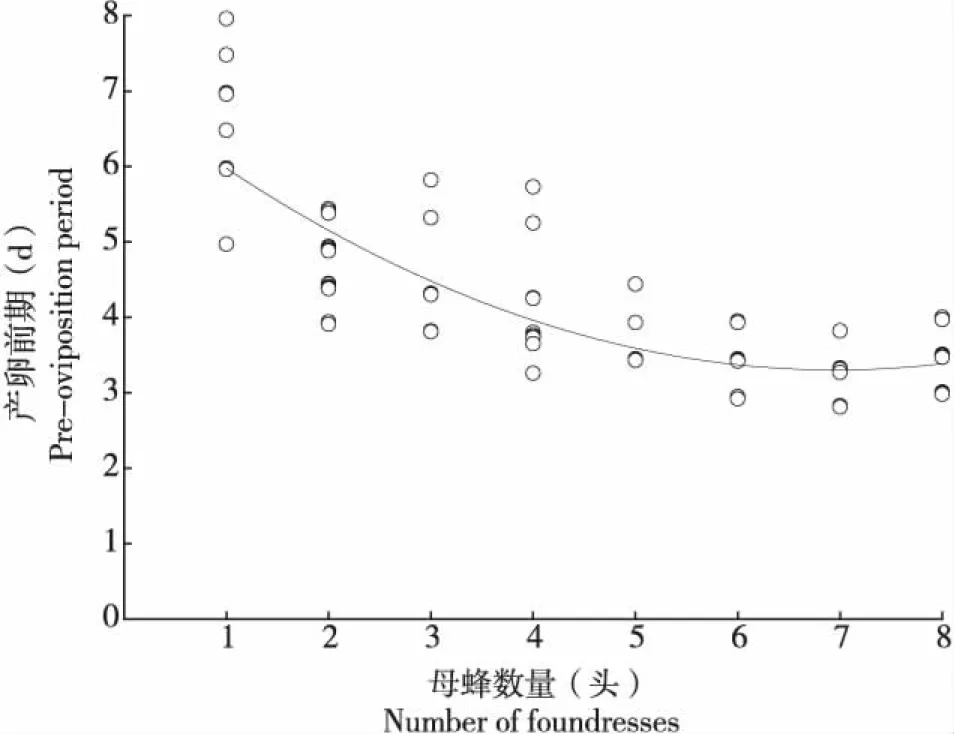
圖3 不同雌蜂密度下管氏腫腿蜂的產卵前期Fig.3 Pre-oviposition duration of Sclerodermus guani in different parasitoid density
不同雌蜂密度下管氏腫腿蜂寄生效率Spearman相關性分析結果表明,雌蜂密度與蟄刺前歷期、寄主麻痹時長及產卵前期存在顯著負相關關系(P<0.01),其相關系數分別為-0.595、-0.381、-0.835,說明隨著雌蜂密度增加,蟄刺前歷期、寄主麻痹時長及產卵前期顯著縮短,尤其是雌蜂密度與產卵前期,相關系數達到0.8以上。
2.2 不同雌蜂密度下寄主致死作用的尋找效應
當寄主密度固定為1頭/寄主不變時,隨著寄生蜂密度的增加寄主致死數量顯著上升(n=160,H=21.268,P<0.05)(表1),當雌蜂密度增加到2頭/寄主時,寄主的致死率達到100%,當雌蜂密度繼續上升時,其對寄主的致死率始終保持在100%。
不同雌蜂密度下管氏腫腿蜂對松墨天牛幼蟲致死作用的尋找效應利用(1)式求得尋找效應E值,然后根據不同雌蜂密度下的腫腿蜂對松墨天牛幼蟲致死作用按照(2)式模型擬合,式中P為腫腿蜂密度1~8頭/寄主。(2)式兩邊取對數為LnE=LnQ-m×LnP經線性回歸可求得m和Q。隨著管氏腫腿蜂密度的增加,雌蜂對松墨天牛幼蟲致死作用的尋找效應逐漸降低(圖4),尋找效應方程為:E=0.9023×P-0.9378(df=1,6,F=1882.4610,P<0.001,r=0.9984)。
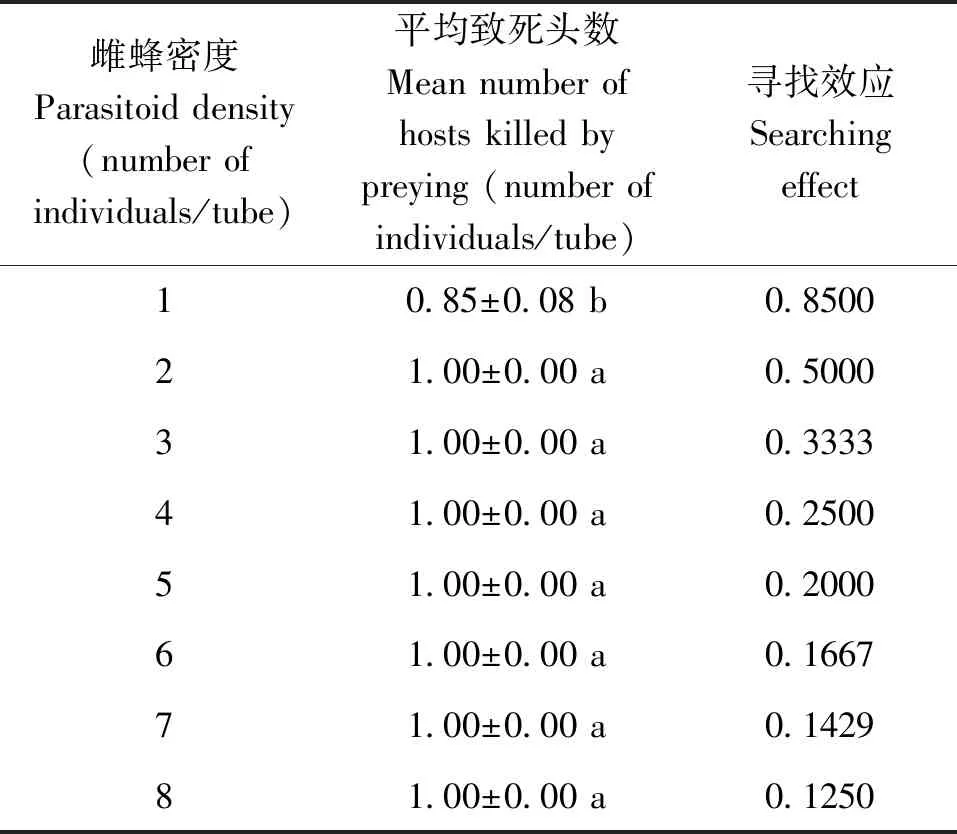
表1 管氏腫腿蜂寄生松墨天牛時自身密度的干擾作用
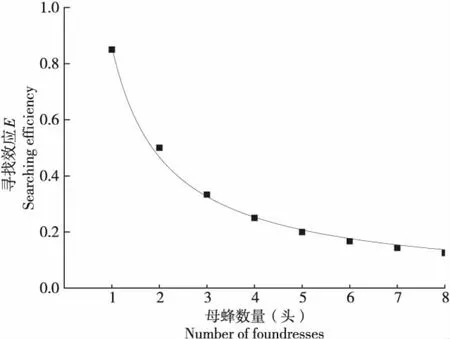
圖4 寄生蜂自身密度對松墨天牛幼蟲致死作用尋找效應的影響Fig.4 Effect of parasitoid densities on the searching efficiency for the killing action on Monochamus alternatus larve
2.3 雌蜂密度對親代生殖力的影響
雌蜂密度顯著影響寄主體上總產卵量及單雌產卵量(總產卵量:n=131,H=82.332,P<0.01;單雌產卵量:n=131,H=102.423,P<0.01)(圖5)。寄主體上總產卵量隨雌蜂密度增加不斷增加,就單頭雌蜂平均產卵量來講,母蜂平均產卵量隨著雌蜂密度增加而顯著減少,雌蜂密度為1頭/寄主時,寄主體上總產卵量及單雌產卵量均為74.769±4.415粒,當雌蜂密度為8頭/寄主時,寄主體上總產卵量高達232.529±8.429粒,但單雌產卵量僅為30.685±0.977粒。
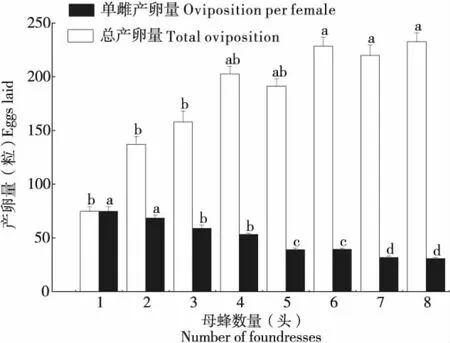
圖5 比較不同雌蜂密度下雌蜂的產卵量Fig.5 Comparison of the oviposition of Sclerodermus guani in different parasitoid density
雌蜂密度與總產卵量及單雌產卵量經Spearman相關性分析結果表明,雌蜂密度與總產卵量存在極顯著正相關關系(P<0.01),與單雌產卵量存在極顯著負相關關系(P<0.01),說明雌蜂密度越大總產卵量越多,但單雌產卵量越少。
雌蜂密度對管氏腫腿蜂的單雌產雌量也有顯著影響(n=131,H=79.484,P<0.01),兩者存在顯著負相關關系(r=-0.765,P<0.001)(圖6)。隨著雌蜂密度增加,寄生蜂的單雌產雌量顯著降低,當雌蜂密度為1頭/寄主時,單雌產雌量為41.077±4.932頭,當雌蜂密度為8頭/寄主時,單雌產雌量下降到15.404±1.134頭。
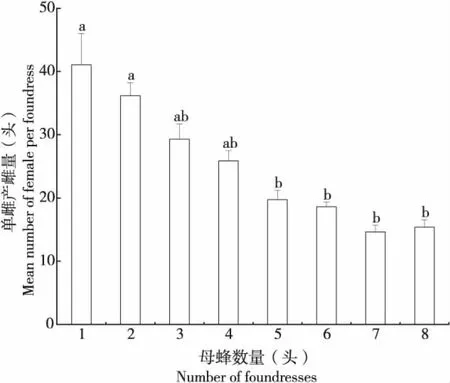
圖6 比較不同雌蜂密度對管氏腫腿蜂單雌產雌量的影響Fig.6 Comparison of the effect of the female offspring per foundress of Sclerodermus guani in different parasitoid density
2.4 不同雌蜂密度下子代適合度的比較
2.4.1子代平均發育歷期和存活率
隨著雌蜂密度增加,管氏腫腿蜂子代卵期縮短(n=154,H=84.641,P<0.01)(表2),雌蜂密度為1頭/寄主時,子代卵期為5.24±0.11 d,雌蜂密度為6頭/寄主時,子代卵期僅為3.55±0.08 d。子代處于幼蟲期時,老熟幼蟲平均發育歷期也存在相同趨勢(n=146,H=55.168,P<0.01),當雌蜂密度達到4頭/寄主時,子代老熟幼蟲發育歷期沒有顯著差異;子代雄蜂繭蛹期平均為13.15~14.24 d,而雌蜂繭蛹期平均為14.05~15.11 d,一般來說雌蜂繭蛹期比雄蜂繭蛹期長 1~2 d。
雌蜂密度對管氏腫腿蜂子代各時期存活率影響明顯(表3)。隨著雌蜂密度增加,低齡幼蟲存活率顯著降低(F7,144=3.196,P<0.01),雌蜂密度增加到6頭/寄主時低齡幼蟲存活率不再明顯下降,雌蜂密度為8頭/寄主時,低齡幼蟲存活率僅為77.68%±2.28%;雌蜂密度對高齡幼蟲和老熟幼蟲存活率影響無明顯差異(高齡幼蟲:F7,144=1.728,P>0.05;老熟幼蟲:F7,140=1.621,P>0.05);雌蜂密度對卵、吐絲幼蟲及成蟲存活率的影響不規律,但隨雌蜂密度增加,存活率呈下降趨勢。
2.4.2子代性比和單雌體重
通過比較分析雌蜂密度對子代性比和單雌體重的影響,結果表明:管氏腫腿蜂子代性比隨雌蜂密度增加呈下降趨勢,不同處理間差異顯著(n=127,H=21.065,P<0.01)(圖7)。當雌蜂密度增加到4頭/寄主以上時,子代性比沒有顯著差異。隨著雌蜂密度增加,子代單雌體重顯著降低(n=136,H=73.011,P<0.01)(圖8),雌蜂密度為1頭/寄主時,單雌平均體重為1.438±0.057 mg/頭,雌蜂密度為2頭/寄主時,單雌平均體重下降到0.897±0.064 mg/頭,當雌蜂密度增加到4頭/寄主時,單雌平均體重不再顯著下降。
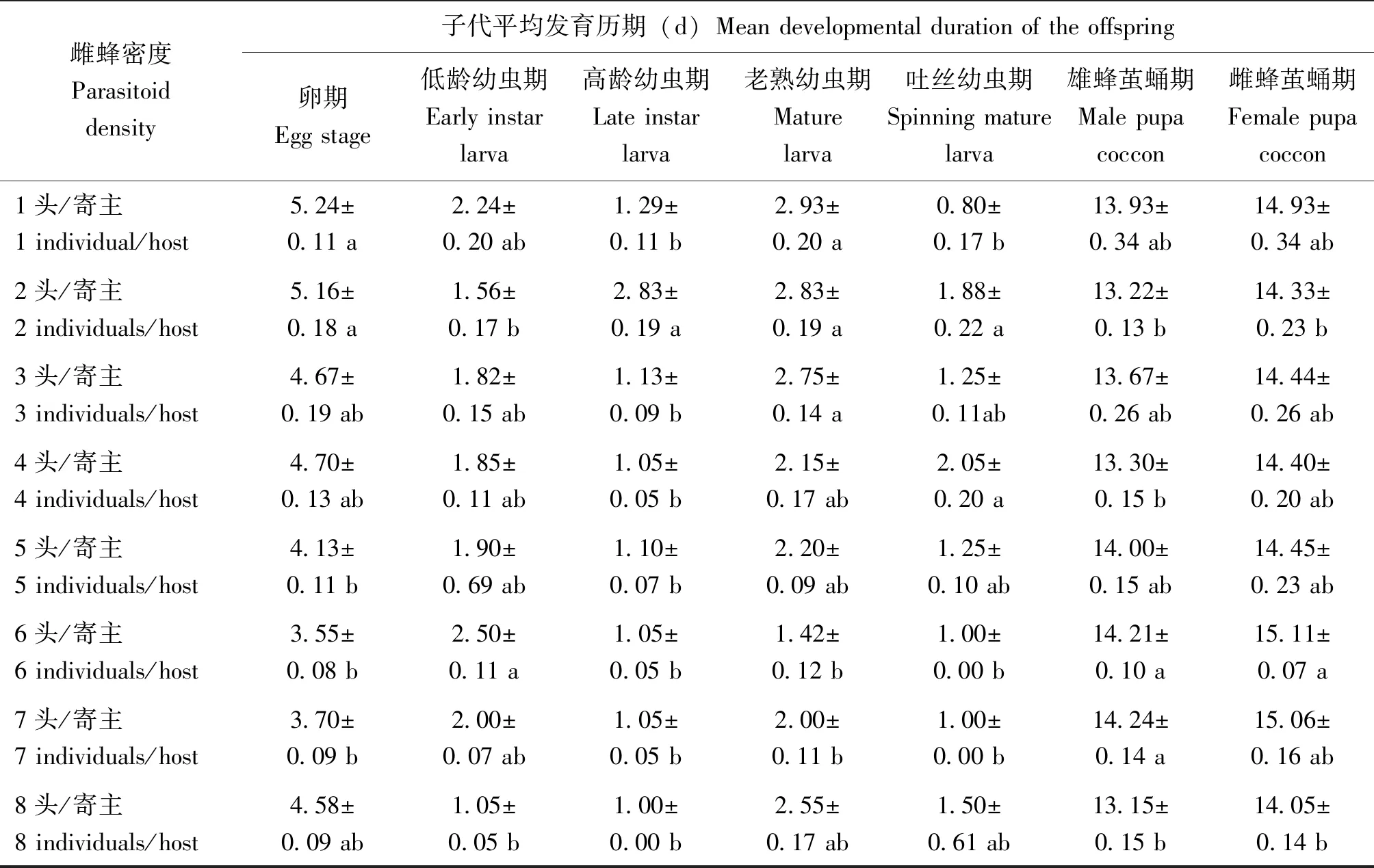
表2 不同雌蜂密度下管氏腫腿蜂子代蜂的平均發育歷期
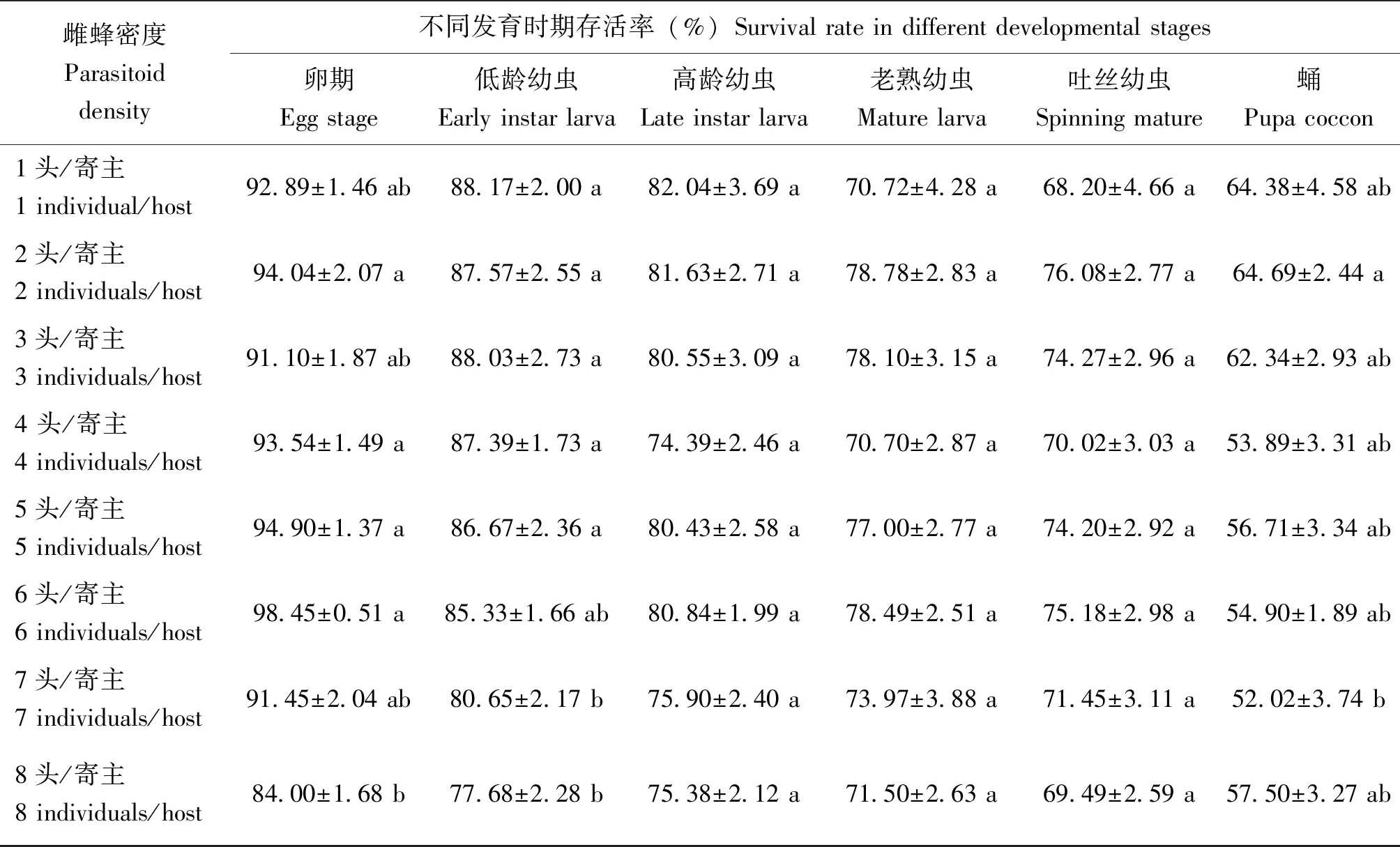
表3 不同雌蜂密度下管氏腫腿蜂子代的存活率Table 3 Offspring survival rate of Sclerodermus guani in different parasitoid density
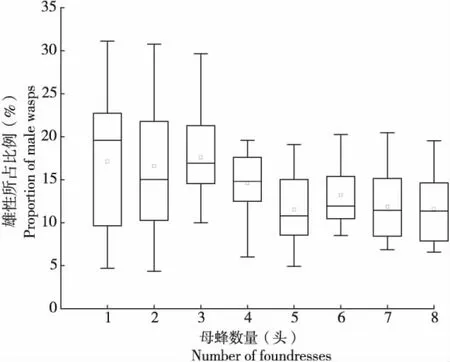
圖7 不同雌蜂密度下管氏腫腿蜂子代的性比Fig.7 Sex ratios of offspring of Sclerodermus guani in different parasitoid density
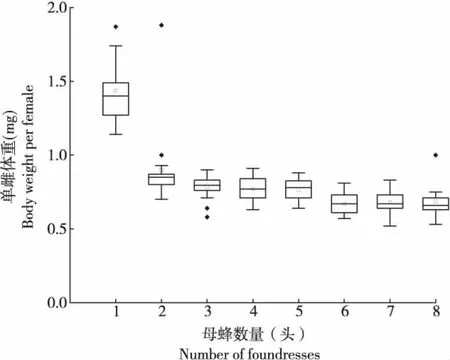
圖8 不同雌蜂密度下子代雌成蟲的體重Fig.8 Female adult weight of offspring of Sclerodermus guani in different parasitoid density
3 結論與討論
在復雜的生態環境中,寄生蜂的生存與繁衍不僅僅受到捕食性和其他寄生性天敵昆蟲的影響,還會遭遇棲息環境中的昆蟲病原微生物的侵染(楊清碰和李莉, 2017; 郭涵等, 2020)。而這些選擇壓力也驅使寄生蜂種群衍生出多種多樣的適應性行為策略,如:雌蜂通過學習行為提高搜索和利用寄主的效率(Lietal., 2009; 唐艷龍等, 2015; Lietal., 2015; 馬琴等, 2019);親代適時撫育以保障子代存活與發育(伍紹龍, 2013; 唐秀云, 2015; 黃維亞, 2017)。作為行為可塑性較強的體外寄生蜂,管氏腫腿蜂兼具這些習性特點(來燕學等, 2012; 楊忠岐, 2018; 胡帥, 2019)。實驗表明,親代雌蜂的寄生效率與雌蜂密度密切相關,當多個雌蜂攻擊單個寄主時能縮短麻痹、制服寄主所消耗的時間,加快從雌蜂取食補充營養到產出子代的歷程,這在一定程度上,無疑提高了管氏腫腿蜂利用寄主繁殖后代的效率,也體現了雌蜂合作的重要性和必要性,尤其是應對個體較大的寄主時,雌蜂自身密度將會給繁殖成功帶來一定收益。此結果與Gao等(2016)報道的白蠟吉丁腫腿蜂Sclerodermuspupariae密度對其寄生效能的結果相一致。然而,實驗也發現,當雌蜂密度過高時,也會面臨生殖力和子代適合度下降的風險。例如:高密度(8頭/寄主)下,單雌平均產卵量及產雌量分別為30.689±0.977粒和15.404±1.134頭,與中等密度和低密度相比明顯降低;低齡幼蟲存活率和單雌體重均有所下降;且在寄主資源有限的條件下,單個幼蟲可獲得的資源變少,其子代發育歷期縮短。此外,腫腿蜂子代發育歷期還與繁育溫度有關,楊希等(2007)在室內繁育腫腿蜂時發現子代發育歷期隨溫度升高而縮短,且在25℃和28℃時蟲蜂比為3∶ 1的情況下,子代發育歷期分別為30 d和24 d,低于20℃時雌蜂不產卵。本研究雌蜂密度為3頭/寄主時,子代發育歷期為25.29 d與其研究相符。由此可見,在實際應用中,既要考慮繁蜂的溫度,還需要權衡寄生蜂密度下“寄生效能-繁殖收益”的矛盾。當雌蜂密度達到4頭/寄主時,能獲得較大的子代數量和質量,且隨雌蜂密度增加子代數量與質量不再明顯增加和下降,因此,雌蜂密度為4頭/寄主才有可能實現繁殖利益最大化。
就管氏腫腿蜂的寄主大多屬于隱蔽性鉆蛀性害蟲的幼蟲或蛹,藏匿在植物的樹干內或韌皮部之下的蛹室或蛀道中,為寄生蜂提供了一個天然的保護屏障(Lietal., 2009)。但這樣的寄主資源呈現斑塊化,因受限于植物種類及栽植密度,較為分散和不均勻,為不善于飛行的寄生蜂搜索適宜寄主帶來難度。所以,一旦雌成蜂尋覓到合適的寄主,如何有效克服寄主抗性、將寄主完全麻痹就是寄生前的最重要環節之一,是親代產卵和子代食物來源的保障,也是降低親代死亡風險的最有效途徑(Liuetal., 2021)。因為單頭雌蜂攻擊中型-大型寄主時,其死亡率高達50%(Liuetal., 2021),寄主在未完全麻痹前均會通過在狹小蛹室中扭動身體或口器反咬的方式來抵御寄生蜂寄生(李元喜和劉樹生, 1999; 唐秀云, 2015);寄主個體越大,寄生蜂蟄刺麻痹寄主的難度越大(辛蓓等, 2016)。本研究發現,當2頭及以上雌蜂攻擊寄主時,雌蜂對寄主的致死率達到了100%,且寄主麻痹時長和產卵前期明顯縮短。這與Tang等(2014)提出的“互助利用寄主假說”相一致,寄生蜂自身密度與寄主的可利用性相適應,就會實現多個寄生蜂個體之間的互惠互利合作。
在寄主資源有限的條件下,最佳寄生者密度對于種間相互協調、子代發育時間和空間以及實現“理想的自由分布”至關重要(Tregenzaetal., 1996)。有研究表明,最佳寄生者密度可以防止過量產卵或繁殖(張燁等, 2015; 羅淑萍等, 2016)。當被引入單頭寄主按石蠅Anastrephafraterculus時,前裂長管繭蜂Diachasmimorphalongicaudata通過分泌物理或者化學標記物來防止超寄生或者過寄生,并減少種內競爭(Franciscoetal., 2020),大多數寄生者使用寄主標記信息素來確定每個寄主能容納的最佳產卵量(Nufio and Papaj, 2001)。具有通訊功能的化學物質可以幫助昆蟲避免過度競爭,適應種群密度,并提高后代的適應性(Klomp, 1964; Addessoetal., 2021)。
此外,管氏腫腿蜂的親代合作不僅表現在聯合攻擊寄主時,更重要的是隨后親代對子代的撫育。親代撫育行為可以提高子代適合度(黃維亞, 2017),且與親緣選擇無關,親生母蜂與非親生母蜂共同撫育時的子代存活率分別顯著高于單獨撫育時的子代存活率(唐秀云, 2015; 伍紹龍等, 2017)。后代不能正常發育(輻照處理)的雌蜂會對其他健康蜂所產的卵進行撫育,行使“幫手蜂”的功能,間接證明單頭雌蜂撫育能力有限,合作撫育能提高子代整體的發育適合度(陳雅竹, 2016)。管氏腫腿蜂的撫育行為不僅受子代體表關鍵化學信息物質的影響(黃維亞, 2017),還與雌蜂體內保幼激素(JH Ⅲ)和蛻皮激素(20 E)的含量變化有關(蔣玉潔, 2021),該蜂的合作撫育受哪些基因的調控有待進一步研究。
據此,可從腫腿蜂科昆蟲行為生物學研究基礎出發,探究昆蟲在自然環境下面對克服寄主時的合作需要與繁殖最大化之間的利益和沖突,明確腫腿蜂科昆蟲的適應性生殖行為策略,將有利于揭示寄生蜂亞社會性到社會性的進化歷程,并為有害生物綜合防治提供重要理論依據。
