器官信號“串話”在代謝綜合征分子機制及潛在治療應用中的研究進展
2023-05-30 22:15:07鄭美華楊明艷孟祥婷齊棟范華英
中國現代醫生 2023年2期
鄭美華 楊明艷 孟祥婷 齊棟 范華英
[摘要] 外周代謝器官和中樞神經系統間的多向性相互作用在維持全身代謝平衡、感知營養物質水平以及應對自然環境挑戰具有重要作用。為了響應不同的代謝需求,人類在長期進化過程中形成一套器官間“相互交流”的生物通信系統。通過該系統,一些組織可以影響遠處組織的代謝功能。然而,當器官間“相互交流”的信號通路失調,可能導致代謝性疾病的發生,如肥胖、糖尿病、肝臟疾病等。肝臟、脂肪組織和胃腸道-內分泌腺是該系統中主要的內分泌器官,除了經典的營養和能量存儲/利用作用外,這些器官還分泌大量生物活性分子,參與機體代謝調節。本文就機體各器官間信號通路“串話”作用進行綜述,重點討論肝臟、脂肪組織以及胃腸道-內分泌腺分泌因子協調全身代謝平衡以及器官間信號網絡,改善代謝綜合征的作用。
[關鍵詞] 代謝綜合征;肝因子;脂肪因子;胃腸道-內分泌腺因子;串話機制
[中圖分類號] R589? ? ? [文獻標識碼] A? ? ? [DOI] 10.3969/j.issn.1673-9701.2023.02.033
人類的新陳代謝涉及一系列復雜的過程,機體各器官間的交流可以維持內環境穩態。為維持全身動態平衡,響應不同的代謝需求,機體的不同器官/組織之間可通過多種信號分子相互“串話”協調代謝穩態,若協調失敗可導致肥胖、糖尿病等代謝性疾病的發生。
1? 概述
代謝綜合征(metabolic syndrome, MetS)是一種廣泛存在的慢性疾病,嚴重影響機體健康和生命安全。肥胖是代謝綜合征的核心[1]。MetS由一組復雜的代謝紊亂疾病群組成,包括肥胖、高血壓、血脂異常、胰島素抵抗等,表現為代謝組織能量攝取和利用失衡。據估計,全球約有超過10億人患有MetS[2]。在我國,60歲以上人群MetS患病率達到50%以上。我國人口老齡化程度的持續加深,使得MetS人群基數越來越大[3]。目前MetS發病機制仍未完全清楚,尚無相關治療藥物獲批,現有的藥物干預手段僅針對MetS單個疾病組成部分,雖能緩解MetS相關癥狀,但治療方案復雜且效果有限,長期聯合應用不良反應較大,且這些藥物均沒有針對MetS的病因進行治療[4]。因此,臨床仍急需開發新的治療性藥物,可以同時針對MetS的所有關鍵特征。
2? 器官信號通路“串話”
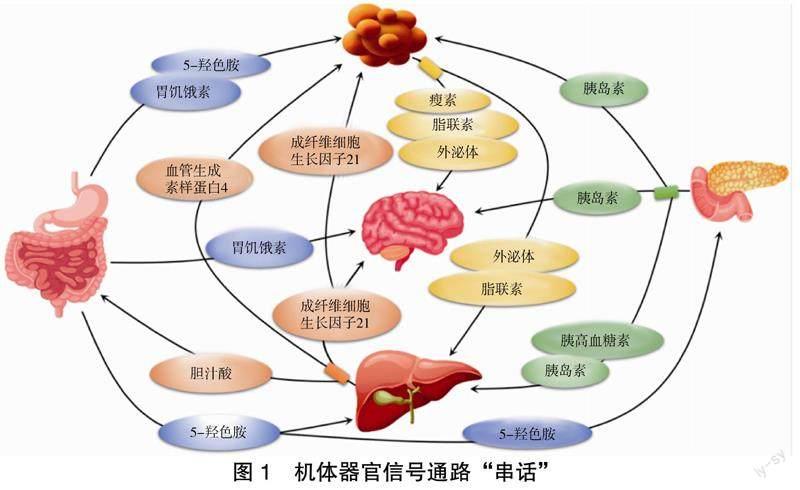
機體各器官/組織,如肝臟、脂肪、腸道等,在調節糖脂代謝方面具有重要的作用。這些器官通過分泌組織因子,如肝臟分泌血管生成素樣蛋白4(angiopoietin-like protein 4,Angptl4)、成纖維細胞生長因子21(fibroblast growth factor 21, FGF21)、膽汁酸(bile acids, BA)等,脂肪組織分泌瘦素、脂聯素、外泌體等,胃腸道-胰腺分泌胰島素、胃饑餓素、5-羥色胺等,與遠處器官進行交流,影響其生理功能,從而協調全身代謝[5],見圖1。
2.1? 肝臟分泌因子
肝臟是一個中樞代謝器官,在維持全身能量穩態方面發揮重要作用。肝臟分泌因子進入血液循環,可對肝臟和外周組織甚至中樞神經系統等靶器官產生強大的影響。肝臟即通過這些分泌因子與其他器官系統進行“串話”。
2.1.1? Angptl4? 主要由肝臟和脂肪組織分泌,與胰島素抵抗、血脂異常、肥胖等代謝性疾病相關。研究發現,Angptl4作用于脂肪組織,過表達會抑制脂肪組織中脂蛋白脂肪酶(lipoprotein lipase,LPL)的催化活性,減少循環脂蛋白中的三酰甘油(triglycerides,TG)水解為游離脂肪酸(free fatty acids,FFA),血漿TG水平升高,導致患心血管代謝疾病的風險升高[6]。據報道,Angptl4過表達可導致葡萄糖耐量受損[7],這與激活胰島素相關信號通路細胞外信號調節蛋白激酶1/2(extracellular signal-regulated protein kinase1/2,ERK1/2)及磷脂酰肌醇-3-激酶(phosphatidylinositol-3-kinase,PI3K)/絲氨酸蘇氨酸蛋白激酶(protein kinase B,AKT)信號通路有關;相反,肝臟Angptl4缺乏可以改善葡萄糖穩態并增強代謝組織中的胰島素敏感性[8]。因此,Angptl4可能是用于改善人類代謝性疾病有希望的靶標。
2.1.2? FGF21? 由肝臟分泌,通過成纖維細胞生長因子受體(fibroblast growth factor receptor,FGFRs)發揮作用,但是FGF21與FGFRs的結合需輔助受體β-klotho(KLB)的介導,在調節糖脂代謝穩態方面具有重要作用[9]。在肝臟中,FGF21通過抑制編碼甾醇調節元件結合蛋白1c(sterol regulatory element-binding transcription factor 1c,SREBP1c)和脂肪酸合酶的脂肪生成基因,抑制肝臟中脂肪的從頭生成[10],可通過結合肝臟中的FGFR2/KLB復合物,調節肝臟膽固醇代謝。除了影響脂代謝外,FGF21也可抑制肝葡萄糖輸出并增加肝糖原的儲存[11],調節葡萄糖代謝。脂肪組織是FGF21的主要靶標,白色脂肪組織(white adipose tissue,WAT)內具有高表達的FGFR1c和KLB,FGF21與其結合,促進脂肪分解;此外,在WAT中,FGF21調節PPARγ的活性并刺激產熱反應,從而促進能量的消耗[12]。Ge等[13]研究顯示葡萄糖轉運蛋白1(glucose transporter type 1,GLUT1)基因通過ERK1/2、血清反應因子和Ets樣蛋白1的轉錄激活,致使FGF21刺激脂肪細胞中的葡萄糖攝,FGF21還可以促進WAT褐變而增加葡萄糖攝取。在大腦中,FGF21通過調節下丘腦中視交叉上核(suprachiasmatic nucleus,SCN)和后腦背迷走神經復合體(dorsal vagal complex,DVC)中的KLB表達,增加代謝率以及改善胰島素敏感性[14]。
2.1.3? BA? 通過法尼醇X受體(farnesoid X receptor,FXR)發揮作用,控制BA、糖脂代謝、能量消耗等,主要作用于肝腸道組織。在肝臟中,FXR可通過FXR-小異二聚體伴侶(orphan nuclear receptor small heterodimer partner,SHP)通路抑制肝臟SREBP1c的表達來減少脂肪生成,還可通過增加LPL激活劑載脂蛋白C-Ⅱ(apolipoprotein C-Ⅱ,ApoCⅡ)的表達而降低LPL抑制劑ApoCⅢ的表達來增強LPL的活性[15],刺激富含TG的脂蛋白的血管內脂解作用。此外,FXR還可下調葡萄糖合成途徑中的磷酸烯醇丙酮酸羧激酶(phosphoenolpyruvate carboxylase,PEPCK)和葡萄糖-6-磷酸酶(glucose- 6-phosphatase,G6Pase),減少糖異生,影響葡萄糖代謝[16]。在胃腸道組織中,腸道FXR的激活可以促進全身脂肪細胞的褐變,增強脂肪分解,防止因飲食引起的肥胖,但是FXR在維持葡萄糖穩態中的作用具有爭議,激活和抑制FXR都可能提高葡萄糖耐量和胰島素敏感性,降低肝糖異生[17]。目前,FXR特異性激動劑奧貝膽酸正在研究中。在一項隨機對照試驗中,患者接受奧貝膽酸治療72周,其肝臟功能顯著改善,體質量減輕約2kg,提高了胰島素敏感性[18]。
2.1.4? 其他分泌因子? ?肝臟的其他分泌因子,如胎球蛋白A/B,其表達增加可導致胰島素抵抗;硒蛋白可以激活AMP激活的蛋白激酶(AMP-activated protein kinase,AMPK),其水平過高時亦可導致胰島素抵抗[19]。
總之,肝臟在控制機體代謝動態平衡方面起核心作用。肝臟通過分泌肝臟因子,以各種方式影響遠處器官的新陳代謝。了解肝臟代謝失調如何影響其他器官系統,可能會為MetS帶來新的治療策略。
2.2? 脂肪分泌因子
脂肪組織作為能量儲存和釋放的主要場所,在控制全身新陳代謝方面具有重要的調節作用。脂肪分泌因子對全身新陳代謝有重要影響,瘦素和脂聯素是經典的脂肪因子,在肥胖和代謝并發癥的發病機制中具有重要關系。
2.2.1? 瘦素? 主要作用于大腦,由皮下WAT分泌,通過與其受體結合發揮負向調節食物攝入量、糖脂代謝穩態以及增加能量消耗的作用[20]。在大腦中,瘦素通過激活阿黑皮質素原(pro-opiomelanocortin,POMC)表達神經元、抑制下丘腦刺鼠相關蛋白(agouti-related neuropeptide,AgRP)和食欲神經肽Y(neuropeptide Y,NPY)的表達,降低機體食欲;瘦素與其受體結合后,可激活多種細胞內信號通路,包括信號轉導和轉錄激活因子3(signal transducer and activator of transcription 3,STAT3)信號通路[21],還可激活絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)和PI3K信號通路,協調食物的攝入和能量的調節來維持機體平衡。
2.2.2? 脂聯素? ?主要作用于肝臟和大腦,由成熟脂肪細胞分泌,具有增強胰島素敏感性、改善肥胖等作用,在肝臟中,通過激活AMPK和PPARα信號通路改善胰島素抵抗[22-23];可通過減少糖異生底物的水平,抑制葡萄糖的產生,從而影響葡萄糖代謝。脂聯素還可影響脂質代謝,通過促進ApoAI和ATP結合和轉運蛋白A1(ATP binding cassette transporter A1,ABCA1)的產生,使高密度脂蛋白膽固醇(high density lipoprotein cholesterol,HDL-C)水平相較與對照組增加30%左右,脂聯素可能與HDL-C水平呈獨立且正相關,通過激活LPL、改善胰島素抵抗,誘導HDL-C上調,TG水平可顯著降低[24],脂聯素還可通過誘導血清ApoC Ⅲ降低,使血清TG水平降低[23]。在大腦中,脂聯素在能量穩態中起重要作用。下丘腦中的脂聯素信號會影響飽腹感及能量穩態[25],還可增強瘦素的影響,增加下丘腦促腎上腺皮質激素釋放激素(corticotropin releasing hormone,CRH)的表達,增加產熱,減輕體重,降低血糖血脂水平;脂聯素在腦室內遞送,可激活下丘腦胰島素信號通路,通過減弱胰島素抵抗改善葡萄糖穩態[26]。
2.2.3? 其他分泌因子? 近年來研究發現,脂肪組織可分泌外泌體,調控肝臟脂質代謝,促進炎癥因子分泌,導致胰島素抵抗的發生[27]。外泌體可作用于下丘腦,調節機體的食欲[28]。抵抗素是肥胖相關胰島素抵抗的介質[29],在葡萄糖代謝中,抵抗素與胰島素信號傳導抑制劑的激活有關。
總之,脂肪組織在調節機體新陳代謝方面發揮核心作用,而脂肪組織的失調會引起其他器官系統的新陳代謝失調。因此,研究脂肪組織在代謝性疾病中的作用,將為MetS的治療提供進一步的見解。
2.3? 胃腸道-內分泌腺分泌因子
胃腸道-內分泌腺分泌的激素包括胃饑餓素、胰島素、5-羥色胺等,作用于中樞神經系統,可改變食物的攝入量,對肝臟、脂肪組織等靶器官也有影響。
2.3.1? 胰島素? 由胰島β細胞釋放,可作用于肝臟、脂肪組織和大腦,抑制肝臟葡萄糖生成,調節脂質代謝。在肝臟中,胰島素直接作用于肝細胞,通過改變肝臟糖異生基因的表達以及抑制PEPCK和G6Pase的活性,減少糖異生;同時還可與其肝臟受體結合,抑制細胞內信號,導致糖原分解迅速減少;通過對PI3K/Akt信號通路的調節,控制肝臟中的脂質代謝與葡萄糖代謝[30]。在脂肪組織中,胰島素通過脂解作用抑制肝葡萄糖的產生[31],作用機制主要包括:①FFA氧化減少使乙酰輔酶A的含量降低,從而減少了丙酮酸羧化酶的變構活化,導致糖異生減少[32];②FFA損害胰島素受體信號傳導,導致胰島素介導的糖原合成減少,G6Pase的活性受到抑制,使糖異生減少;③FFA減少增強葡萄糖介導的對糖原分解的抑制[31]。此外,胰島素還可以通過觸發GLUT4葡萄糖轉運體移位到細胞膜,促進葡萄糖的吸收并迅速降低血糖[33]。
2.3.2? 胃饑餓素? ?由胃黏膜釋放,主要作用于大腦和脂肪組織,具有促食欲特性,在調節體重方面具有重要作用,可影響食物攝入及減少能量消耗。在大腦中,胃饑餓素通過激活下丘腦AMPK信號通路發揮其促食欲作用[34],胃饑餓素還可以降低BAT交感神經活動[35],調節脂肪細胞代謝,通過促進葡萄糖和TG攝取、增加脂肪生成和抑制白色脂肪細胞中的脂質氧化,促進脂肪儲存。在脂肪組織中,胃饑餓素可增加WAT的儲存,降低BAT中與產熱相關的解偶聯蛋白的表達[36]。
2.3.3? 其他分泌因子? 5-羥色胺由腸道分泌,對胰腺、肝臟和脂肪組織中的葡萄糖代謝具有重要影響[37];胰高血糖素由胰島α細胞分泌,可促進糖原分解和糖異生,提示胰高血糖素是治療MetS的一種途徑[38]。胃腸道-內分泌腺分泌因子在治療MetS方面具有潛在的作用。胃饑餓素和胰島素的調節作用目前正在研究中,作為一種治療方法,已顯示出在治療MetS方面的希望。
2.4? “串話”和代謝影響
除了單獨了解每種肝因子、脂肪因子和胃腸道-內分泌腺因子的作用之外,重要的是這些組織因子在機體各器官間“串話”的共同作用,在不同的組織中形成復雜的作用網絡,改善機體葡萄糖及脂質代謝紊亂,從而對肥胖、2型糖尿病、代謝綜合征等疾病產生影響。了解這種“串話”機制對于制定預防和治療措施以更好地管理其所涉及的并發癥是有效的。
3? 總結與展望
肝臟、脂肪組織及胃腸道-內分泌腺分泌因子,通過器官間“串話”進行交流,可作為體內平衡紊亂的生物標志物,在控制全身新陳代謝的過程中具有重要作用。從多肽激素到小分子再到脂質,這些分泌因子協調靶器官的生理過程,作為潛在的治療靶點發揮作用。本文討論了與經典代謝相關器官的分泌因子,這些器官系統間可相互作用,影響機體新陳代謝。因此,了解肝臟分泌因子、脂肪組織分泌因子和胃腸道-內分泌腺分泌因子的釋放和相互作用,或可成為改善MetS新的治療選擇。
[參考文獻]
[1]WANG H H, LEE D K, LIU M, et al. Novel insights into the pathogenesis and management of the metabolic syndrome[J]. Pediatr Gastroenterol, 2020, 23(3): 189–230.
[2]SAKLAYEN M G. The global epidemic of the metabolic syndrome[J]. Curr Hypertens Rep, 2018, 20(2): 12.
[3]LI R, LI W, LUN Z, et al. Prevalence of metabolic syndrome in Mainland China: a meta-analysis of published studies[J]. BMC Public Health, 2016, 16(1): 1–10.
[4]PRIEST C, TONTONOZ P. Inter-organ cross-talk in metabolic syndrome[J]. Nat Metab, 2019, 1(12): 1177–1188.
[5]de OLIVEIRA DOS SANTOS A R, DE OLIVEIRA ZANUSO B, MIOLA V F B, et al. Adipokines, myokines, and hepatokines: crosstalk and metabolic repercussions[J]. Int J Mol Sci, 2021, 22(5): 2639.
[6]SINGH A K, CHAUBE B, ZHANG X, et al. Hepatocyte-specific suppression of ANGPTL4 improves obesity-associated diabetes and mitigates atherosclerosis in mice[J]. J Clin Invest, 2021, 131(17): e140989.
[7]HARADA M, YAMAKAWA T, KASHIWAGI R, et al. Association between ANGPTL3, 4, and 8 and lipid and glucose metabolism markers in patients with diabetes[J]. PLoS One, 2021, 16(7): e0255147.
[8]GUSAROVA V, O'DUSHLAINE C, TESLOVICH T M, et al. Genetic inactivation of ANGPTL4 improves glucose homeostasis and is associated with reduced risk of diabetes[J]. Nat Commun, 2018, 9(1): 2252.
[9]GENG L, LAM K S L, XU A. The therapeutic potential of FGF21 in metabolic diseases: from bench to clinic[J]. Nat Rev Endocrinol, 2020, 16(11): 654–667.
[10]ZHANG Y, LEI T, HUANG J F, et al. The link between fibroblast growth factor 21 and sterol regulatory element binding protein 1c during lipogenesis in hepatocytes[J]. Mol Cell Endocrinol, 2011, 342(1–2): 41–47.
[11]LIU J, YANG K, YANG J, et al. Liver-derived fibroblast growth factor 21 mediates effects of glucagon-like peptide-1 in attenuating hepatic glucose output[J]. EBioMedicine, 2019, 41: 73–84.
[12]HAN M S, PERRY R J, CAMPOREZ J P, et al. A feed-forward regulatory loop in adipose tissue promotes signaling by the hepatokine FGF21[J]. Genes Dev, 2021, 35(1-2): 133–146.
[13]GE X, CHEN C, HUI X, et al. Fibroblast growth factor 21 induces glucose transporter-1 expression through activation of the serum response factor/Ets-like protein-1 in adipocytes[J]. J Biol Chem, 2011, 286(40): 34533–34541.
[14]BONDURANT L D, AMEKA M, NABER M C, et al. FGF21 regulates metabolism through adipose- dependent and independent mechanisms[J]. Cell Metab, 2017, 25(4): 935–944.e4.
[15]CLAUDEL T, INOUE Y, BARBIER O, et al. Farnesoid X receptor agonists suppress hepatic apolipoprotein CⅢ expression[J]. Gastroenterology, 2003, 125(2): 544–555.
[16]SHAPIRO H, KOLODZIEJCZYK A A, HALSTUCH D, et al. Bile acids in glucose metabolism in health and disease[J]. J Exp Med, 2018, 215(2): 383–396.
[17]TICHO A L, MALHOTRA P, DUDEJA P K, et al. Intestinal absorption of bile acids in health and disease[J]. Compr Physiol, 2019, 10(1): 21–56.
[18]MCGLONE E R, BLOOM S R. Bile acids and the metabolic syndrome[J]. Ann Clin Biochem, 2019, 56(3): 326–337.
[19]MISU H, TAKAYAMA H, SAITO Y, et al. Deficiency of the hepatokine selenoprotein P increases responsiveness to exercise in mice through upregulation of reactive oxygen species and AMP-activated protein kinase in muscle[J]. Nat Med, 2017, 23(4): 508–516.
[20]ABELLA V, SCOTECE M, CONDE J, et al. Leptin in the interplay of inflammation, metabolism and immune system disorders[J]. Nat Rev Rheumatol, 2017, 13(2): 100–109.
[21]CHEN Y, WU R, CHEN H Z, et al. Enhancement of hypothalamic STAT3 acetylation by nuclear receptor Nur77 dictates leptin sensitivity[J]. Diabetes, 2015, 64(6): 2069–2081.
[22]MATSUDA M, SHIMOMURA I. Roles of adiponectin and oxidative stress in obesity-associated metabolic and cardiovascular diseases[J]. Rev Endocr Metab Disord, 2014, 15(1): 1–10.
[23]OKU H, MATSUURA F, KOSEKI M, et al. Adiponectin deficiency suppresses ABCA1 expression and ApoA-I synthesis in the liver[J]. FEBS Lett, 2007, 581(26): 5029–5033.
[24]YANAI H, YOSHIDA H. Beneficial effects of adiponectin on glucose and lipid metabolism and atherosclerotic progression: mechanisms and perspectives[J]. Int J Mol Sci, 2019, 20(5): 1190.
[25]BLOEMER J, PINKY P D, GOVINDARAJULU M, et al. Role of adiponectin in central nervous system disorders[J]. Neural Plast, 2018, 2018: 4593530.
[26]PARK S, KIM DS , KWON D Y, et al. Long-term central infusion of adiponectin improves energy and glucose homeostasis by decreasing fat storage and suppressing hepatic gluconeogenesis without changing food intake[J]. J Neuroendocrinol, 2011, 23(8): 687–698.
[27]XIE Z, WANG X, LIU X, et al. Adipose-derived exosomes exert proatherogenic effects by regulating macrophage foam cell formation and polarization[J]. J Am Heart Assoc, 2018, 7(5): e007442.
[28]GAO J, LI X, WANG Y, et al. Adipocyte-derived extracellular vesicles modulate appetite and weight through mTOR signalling in the hypothalamus[J]. Acta Physiol (Oxf), 2020, 228(2): e13339.
[29]MUSOVIC S, SHRESTHA M M, KOMAI A M, et al. Resistin is co-secreted with adiponectin in white mouse adipocytes[J]. Biochem Biophys Res Commun, 2021, 534: 707–713.
[30]TITCHENELL P M, LAZAR M A, BIRNBAUM M J. Unraveling the regulation of hepatic metabolism by insulin[J]. Trends Endocrinol Metab, 2017, 28(7): 497–505.
[31]LEWIS G F, CARPENTIER A C, PEREIRA S, et al. Direct and indirect control of hepatic glucose production by insulin[J]. Cell Metab, 2021, 33(4): 709–720.
[32]PETERSEN M C, SHULMAN G I. Mechanisms of insulin action and insulin resistance[J]. Physiol Rev, 2018, 98(4): 2133–2223.
[33]KLIP A, MCGRAW T E, JAMES D E. Thirty sweet years of GLUT4[J]. J Biol Chem, 2019, 294(30): 11369–11381.
[34]LV Y, LIANG T, WANG G, et al. Ghrelin, a gastrointestinal hormone, regulates energy balance and lipid metabolism[J]. Biosci Rep, 2018, 38(5): BSR20181061.
[35]SOVETKINA A, NADIR R, FUNG J N M, et al. The physiological role of ghrelin in the regulation of energy and glucose homeostasis[J]. Cureus, 2020, 12(5): e7941.
[36]THEANDER-CARRILLO C, WIEDMER P, CETTOUR-ROSE P, et al. Ghrelin action in the brain controls adipocyte metabolism[J]. J Clin Invest, 2006, 116(7): 1983–1993.
[37]MARAZZITI D, BETTI L, BARONI S, et al. The complex interactions among serotonin, insulin, leptin, and glycolipid metabolic parameters in human obesity[J]. CNS Spectr, 2022, 27(1): 99–108.
[38]AL-MASSADI O, FERN? J, DI?GUEZ C, et al. Glucagon control on food intake and energy balance[J]. Int J Mol Sci, 2019, 20(16): 3905.
(收稿日期:2022–07–22)
(修回日期:2022–12–07)
