
雞cGAS基因的克隆、表達特性分析及FAdV-4感染前后亞細胞定位變化
2023-06-08 23:29:47曹影麗劉琪柴震震楊侃侃梁月巧宋祥軍邵穎涂健祁克宗
江蘇農業學報 2023年2期
曹影麗 劉琪 柴震震 楊侃侃 梁月巧 宋祥軍 邵穎 涂健 祁克宗

摘要: cGAS作為一種新型的胞質DNA受體,在宿主抵抗DNA病毒而觸發的天然免疫中起著至關重要的作用。血清4型禽腺病毒(Fowl adenovirus serotype 4,FAdV-4)是無囊膜的一種雙鏈DNA病毒,可引起雞肝炎-心包積液綜合征。為明確雞cGAS(chcGAS)基因功能,探究在雞肝癌(LMH)細胞中過表達chcGAS以及FAdV-4感染前后細胞定位的變化情況。本研究針對chcGAS序列設計引物進行PCR擴增,并分析該基因序列與其他物種之間的同源性以及預測該基因的結構域,構建重組質粒pET-32a-chcGAS進行原核表達,通過SDS-PAGE和Western blot鑒定,利用激光共聚焦觀察FAdV-4感染LMH細胞前后chcGAS細胞定位變化。結果表明,本試驗克隆得到大小為1 317 bp的 chcGAS基因,與其他物種同源性為50.5%~84.4%,SDS-PAGE分析結果顯示,chcGAS蛋白主要以可溶性蛋白質的形式在上清液處表達,目的蛋白質相對分子質量為75 000,與預期大小相符。亞細胞定位結果顯示,FAdV-4感染可導致定位于細胞核膜上的chcGAS蛋白轉移到細胞質中。本研究結果為進一步研究FAdV-4與cGAS-STING信號通路的關聯調控機制提供了科學依據。
關鍵詞: cGAS;血清4型禽腺病毒(FAdV-4);序列分析;原核表達;亞細胞定位
中圖分類號: S831 文獻標識碼: A 文章編號: 1000-4440(2023)02-0453-08
Cloning and expression characteristics of chicken cGAS gene and changes in subcellular localization before and after FAdV-4 infection
CAO Ying-li, LIU Qi, CHAI Zhen-zhen, YANG Kan-kan, LIANG Yue-qiao, SONG Xiang-jun, SHAO Ying, TU Jian, QI Ke-zong
(Anhui Key Laboratory of Veterinary Pathobiology and Disease Control/Anhui Province Animal Food Quality and Biosafety Engineering Laboratory, Hefei 230036, China)
Abstract: cGAS plays a key role in the hosts innate immune response against DNA viruses as a receptor for DNA recognition within the cytoplasm. Fowl adenovirus serotype 4 (FAdV-4) is a double-stranded DNA virus without an envelope, and can cause hepatitis-pericardial effusion syndrome in chicken. In order to clarify the function of chicken cGAS (chcGAS) gene, the overexpression of chcGAS in chicken liver cancer (LMH) cells and the changes of cell location before and after FAdV-4 infection were investigated. In this study, the primers were designed according to the chcGAS sequence, and the chcGAS gene was amplified by PCR. The homology between the gene sequence and other species was analyzed, and the domain was predicted. The recombinant plasmid pET-32a-chcGAS was constructed for prokaryotic expression, identified by SDS-PAGE and Western blot, and the localization changes of chcGAS in LMH cells before and after FAdV-4 infection were observed by laser confocal microscopy. The results showed that the chcGAS gene with a size of 1 317 bp was successfully cloned, and the homology with other species was 50.5%-84.4%. The results of SDS-PAGE analysis indicated that chcGAS protein was mainly expressed in the supernatant in the form of soluble protein. The relative molecular mass of target protein was 75 000, which was consistent with the expected size. The results of subcellular localization showed that FAdV-4 infection could lead to transfer of chcGAS protein localized on the nuclear membrane into the cytoplasm. These results can provide a scientific basis for further studying the correlation regulatory mechanism between FAdV-4 and cGAS-STING signaling pathways.
Key words: cGAS;fowl adenovirus serotype 4(FAdV-4);sequence analysis;prokaryotic expression;subcellular localization
當外界的病原微生物或其產物進入機體時,細胞內的模式識別受體(PRRs)活化,進而產生I型干擾素(IFN)和一些促炎細胞因子[1-3]。在過去的幾年里,模式識別受體研究領域將胞質DNA感受器作為重要研究方向[4-6]。PRRs是一類天然免疫分子,是天然免疫受體識別的重要組成部分[7-8]。2013年Wu等[9]發現細胞中存在某種物質可以通過識別胞質內的DNA激活IFN-β信號通路,質譜檢測分析后發現是一種新的物質,這種物質為環磷酸鳥苷-腺苷(cGAMP),并且該物質可以通過與干擾素刺激基因(STING)結合從而激活IFN信號通路。也有研究結果表明,cGAS可以作為一種廣譜胞質DNA受體識別胞質內DNA并激活IFN信號通路[10]。常規情況下,細胞質內存在的DNA較少,故以cGAS為主的胞質DNA感受器處于未活化的狀態。但當細胞處于某些應激狀態時,比如DNA病毒感染,其胞質內的DNA含量上升,隨后可通過cGAS-STING通路發揮抗病毒作用。目前,普遍認為cGAS定位于細胞質中,而Barnett 等[11]指出,cGAS并非傳統意義上的胞質蛋白質,而是通過其N端磷酸肌醇結合域的作用從而出現膜定位的一種蛋白質,并提到在靜息狀態下,cGAS定位在細胞膜,而識別DNA病毒感染后可出現由細胞膜到細胞質轉位的現象。
目前,關于鳥類中cGAS的研究也有報道。楊潔[12]的研究結果表明,cGAS和STING信號軸對DNA病毒、RNA病毒和反轉錄病毒均有抗病毒作用,證明其廣泛的抗病毒功能和在雞先天免疫中的關鍵作用。Oliveira 等[13]的研究結果表明,在雞巨噬細胞中,cGAS/STING通路不但能產生I型干擾素以響應細胞內DNA刺激,而且對調節巨噬細胞效應器功能[包括組織相容性復合體(MHC-II)和共刺激分子的表達]至關重要。在禽痘病毒(一種禽DNA病毒)感染的情況下,發現cGAS/STING途徑與I型干擾素的產生及MHC-II轉錄有關。禽腺病毒4型屬于腺病毒科禽腺病毒屬,是無囊膜的雙鏈 DNA 病毒。血清4型禽腺病毒(FAdV-4)是肝炎-心包積液綜合征的主要病原體。作為一種DNA病毒,目前Wang等[14]通過在雞胚成纖維細胞(CEF)中瞬時轉染雞cGAS基因(chcGAS)來研究其亞細胞定位,結果表明,chcGAS主要定位在細胞質中。通過過表達chcGAS基因和RNA干擾chcGAS基因,證明了chcGAS對外源性雙鏈DNA(dsDNA)以及來自DNA損傷反應的自身dsDNA有反應,從而激活了STING/TBK1/IRF7介導的先天免疫。目前,對于 chcGAS的研究著重在其作為胞質 DNA感受器觸發一系列通路方面的探索,但關于雞cGAS在體外的蛋白質表達以及雞cGAS在DNA病毒感染后細胞內定位、轉位的研究較少。cGAS對胞質內DNA的識別是非特異性的,幾乎可以識別所有的雙鏈DNA [15-16] 。
本研究擬通過克隆chcGAS的完整開放閱讀框序列,構建原核表達載體,獲得chcGAS蛋白,此外,通過對比chcGAS被FAdV-4感染前后細胞定位的變化更進一步闡述chcGAS在觸發天然免疫激活中的作用,以期為進一步研究FAdV-4與cGAS-STING信號通路的關聯調控機制提供科學依據。
1 材料與方法
1.1 細胞和毒株
雞肝癌(LMH)細胞由中國農業科學院上海獸醫研究所劉光清研究員贈送。FAdV-4 AH-F19株 [17](登錄號:MN781666)由本實驗室分離并經過鑒定后保存于-80 ℃冰箱。
1.2 菌株和質粒
感受態細胞DH5α、Rosseta(DE3)購于南京擎科生物科技有限公司。pET-32a、pCAGGS-HA、pmCherry-C1質粒由本實驗室保存于-20 ℃冰箱。
1.3 主要試劑
SanPrep柱式DNA膠回收試劑盒、ECL發光顯色工作液、質粒小量提取試劑盒購自上海生工生物工程技術服務有限公司,2×Hiff Canace? Gold PCR Master Mix購自上海翌圣生物科技有限公司,去內毒素質粒提取試劑盒購自北京天漠科技開發有限公司,PAGE蛋白凝膠快速配制試劑盒、5×蛋白質上樣緩沖液、雅酶三色預染Marker購自上海雅酶生物醫藥科技有限公司,EcoR I限制性內切酶購自寶日醫生物技術(北京)有限公司。同源重組連接酶購自南京諾唯贊生物科技股份有限公司,鼠抗HA單克隆抗體、鼠抗His單克隆抗體購自艾比瑪特生物醫藥(上海)有限公司,FITC標記二抗羊抗鼠、山羊抗鼠IgG-HRP購自武漢博士德生物工程有限公司。
1.4 序列分析與結構域預測
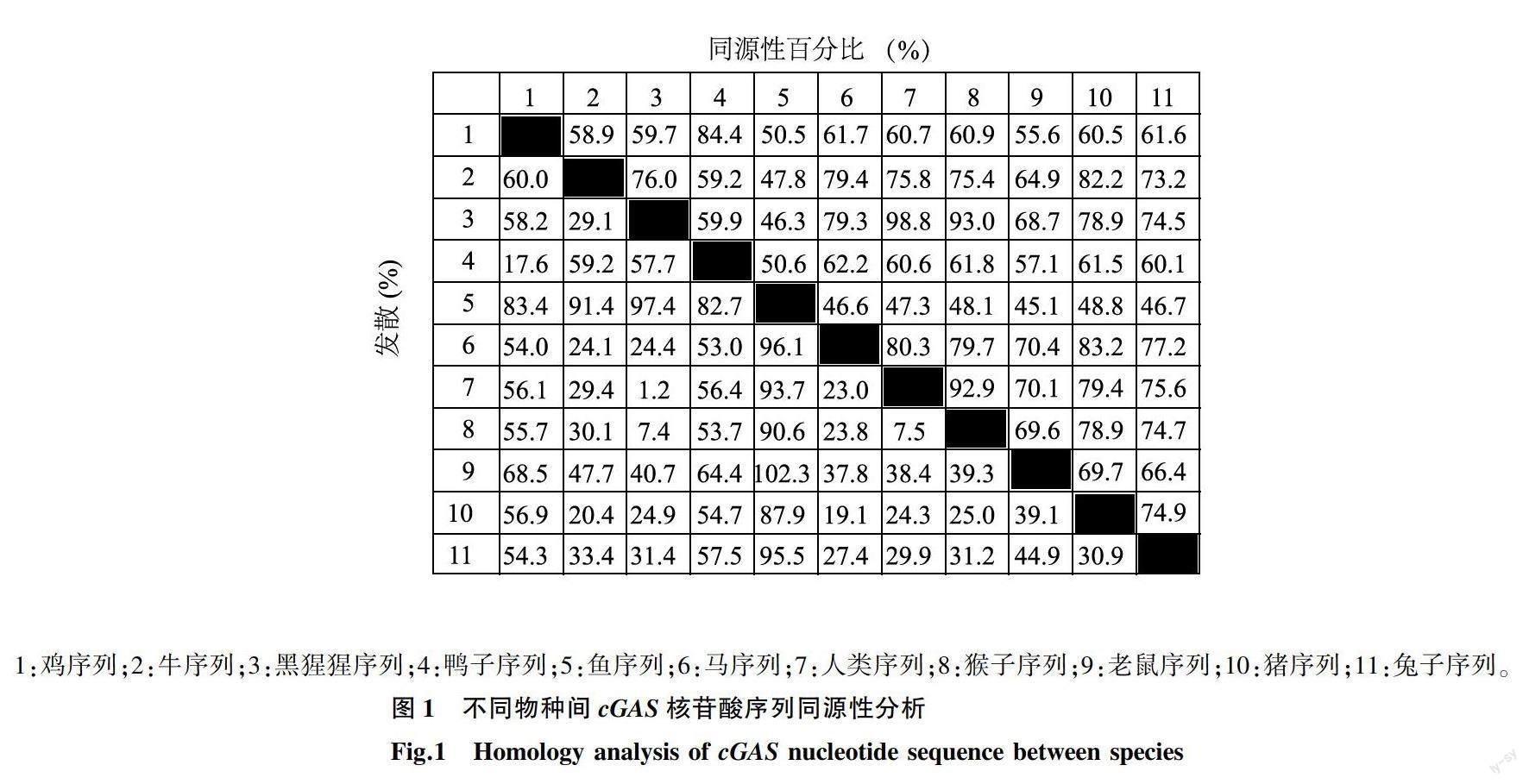
從NCBI中下載已發表的一些物種的cGAS序列,通過Megalign軟件進行同源性分析并利用MEGA5.0對各物種序列構建遺傳進化樹。用SMART在線網站(http://smart.embl-heidelberg.de/)預測chcGAS的結構域。
1.5 引物設計與基因擴增
根據GenBank中發表的chcGAS基因序列,使用Primer Premier 5軟件設計基因擴增的上下游引物,引物合成序列見表1。按照RNA提取試劑盒說明書提取LMH細胞中的總RNA并反轉錄為cDNA,以此為模板擴增chcGAS特異性引物。
1.6 重組質粒chcGAS載體的構建
根據chcGAS基因序列,利用CE DesignV1.04軟件依次設計pET-32a-chcGAS、pCAGGS-HA-chcGAS、pmCherry-C1-chcGAS同源臂引物,引物合成序列見表1。
PCR反應體系(50 μl):2×Hiff Canace?Gold PCR Master Mix 25 μl、ddH2O 19 μl、 DNA模板2 μl、上下游引物各2 μl 。PCR反應條件:98 ℃預變性3 min;98 ℃變性10 s,58 ℃ 退火20 s,72 ℃延伸3 min,共35個循環;72 ℃再延伸5 min。利用PCR擴增含有酶切位點的chcGAS基因,PCR產物分別切膠回收,將其克隆到經EcoR I酶切的pET-32a、pCAGGS-HA、pmCherry-C1空載體中。重組質粒經PCR鑒定正確后移交到測序公司進行測序。重組質粒經鑒定后分別命名為pET-32a-chcGAS、pCAGGS-HA-chcGAS、pmCherry-C1-chcGAS。
1.7 pET-32a-chcGAS誘導表達與可溶性分析
將pET-32a-chcGAS與pET-32a空載體質粒分別轉化Rosetta(DE3)感受態細胞,涂布到氨芐抗性的固體培養板,次日分別挑取單菌落至溶菌肉湯(LB)液體培養基中。用搖床37 ℃、180 r/min搖4~5 h后轉接到大容積的LB液體培養基中,再次用搖床37 ℃、180 r/min培養,當OD600值為0.6~0.8時,添加0.5 mmol/L的異丙基硫代半乳糖苷(IPTG)于15 ℃、120 r/min誘導表達,取誘導20 h后的菌體,用磷酸鹽緩沖液(PBS)進行洗菌,隨后超聲裂解至液體清亮,離心后分別吸取上清液和沉淀進行SDS-PAGE電泳,而后經考馬斯亮藍染色、脫色后進行觀察和分析。通過分析確定重組蛋白質表達可溶性。
1.8 Western-blot檢測
將誘導后處理好的蛋白質樣品,在 SDS-PAGE凝膠電泳后轉至聚偏二氟乙烯(PVDF)膜上,恒流100 mA轉膜1.0 h。將轉膜后的PVDF膜放入配制好的5%脫脂奶粉封閉液中,隨后移入4 ℃冰箱過夜封閉,用磷酸緩沖液(PBST)洗滌2~3次,按照1∶5 000(體積比)加入鼠抗His單克隆抗體作為一抗,常溫下孵育1.5 h,在用PBST洗滌2~3次,同樣按照1∶5 000(體積比)加入山羊抗鼠lgG-HRP作為二抗,常溫孵育1.0 h,用PBST洗滌2~3次,隨后用辣根過氧化物酶化學發光劑(HRP-ECL)底物發光顯色工作液進行顯色。
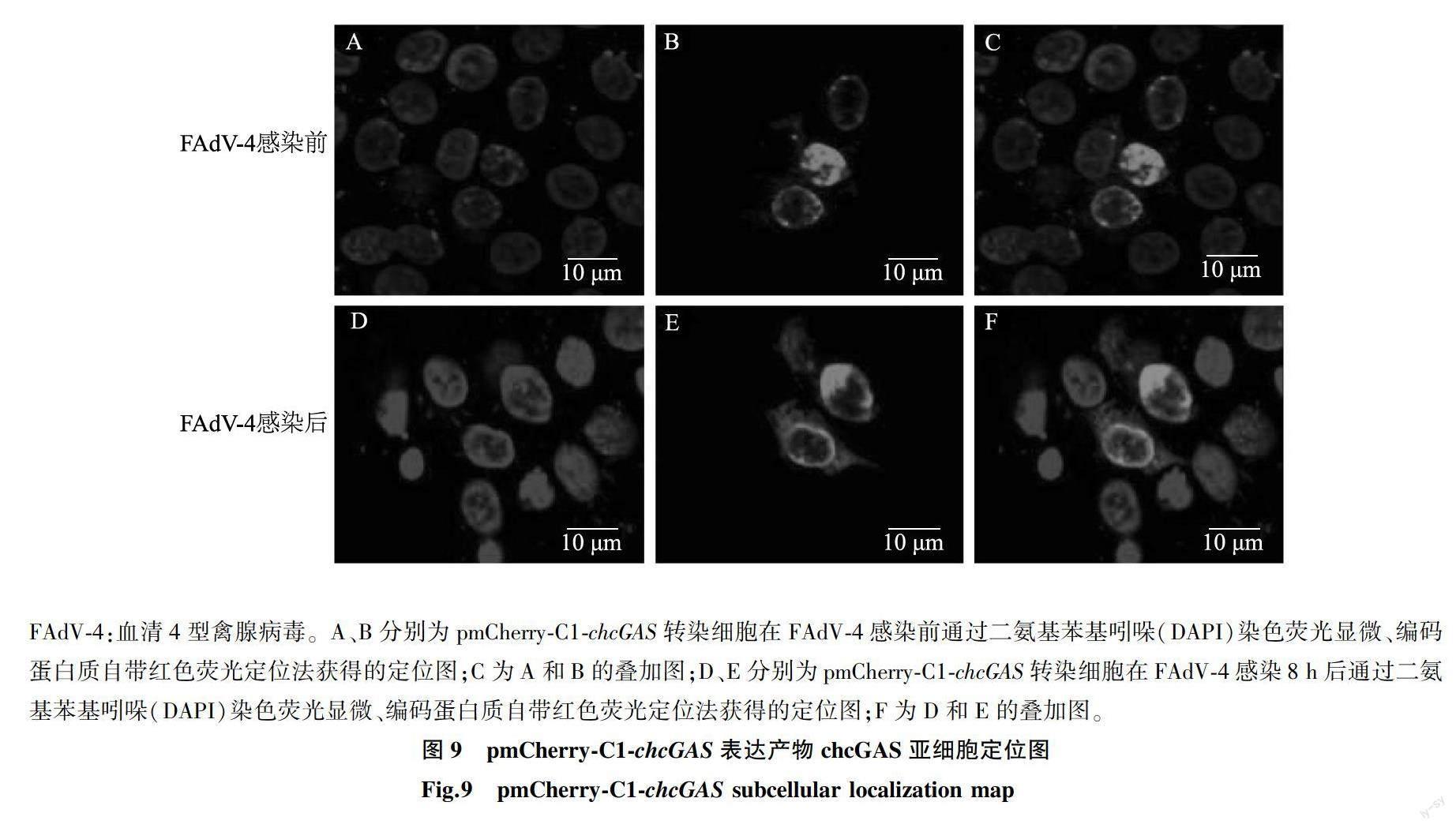
1.9 chcGAS感染前后亞細胞定位
為了觀察chcGAS在FAdV-4感染前后亞細胞定位變化,設置2組試驗,分為感染組和對照組(未感染),分別將2組長滿的LMH細胞接種于鋪上細胞爬片的12孔板中,觀察細胞,細胞密度達到80%左右將pCAGGS-HA-chcGAS重組質粒轉染進細胞,對照組24 h后棄培養液,換成新鮮的維持培養液,感染組在質粒轉染24 h后接種FAdV-4 AH-F19株,37 ℃培養1 h,棄去培養液,用PBS清洗細胞2次,感染8 h后分別收取感染組和對照組樣品,通過間接免疫熒光步驟,分別使用HA標簽鼠抗多克隆抗體作為一抗,FITC綠色熒光抗體作為二抗,而后利用激光共聚焦觀察分析chcGAS在LMH細胞中的定位情況。
為了進一步證實chcGAS的定位,同樣將pmCherry-C1-chcGAS重組質粒轉染到LMH細胞,以相同的方法對轉染后的細胞進行收樣,利用pmCherry-C1載體自帶紅色熒光在激光共聚焦顯微鏡下觀察chcGAS定位情況,觀察是否與pCAGGS-HA-chcGAS重組質粒定位情況一致。
2 結果與分析
2.1 序列分析與結構域預測
通過Megalign比對分析物種間cGAS核苷酸同源性,結果顯示,chcGAS與其他物種cGAS的同源性為50.5%~84.4%(圖1)。將chcGAS與其他物種cGAS核苷酸序列比對并對系統發育樹進行分析,結果顯示,chcGAS與鴨子cGAS位于同一分支上,親緣關系較近;但與魚類cGAS和哺乳類cGAS不在同一分支上,表明親緣關系稍遠(圖2)。利用SMART在線網站預測chcGAS的結構域,結果顯示,編碼區包括2個低復雜區域(Low-Complexity region, LCR),分別為第3至第39氨基酸與第68至第81氨基酸,還包括1個高度保守的 Mab-21結構域 (第129至第429氨基酸)(圖3)。
2.2 chcGAS基因擴增結果
經PCR擴增chcGAS基因得到目的條帶(圖4),目的片段大小為約1 317 bp,顯示與預期條帶大小一致,經測序比對后與目的基因序列完全一致。
2.3 重組質粒的酶切鑒定
將pET-32a-chcGAS、pCAGGS-HA-chcGAS、pmCherry-C1-chcGAS重組質粒經EcoR I酶切鑒定后在核酸凝膠電泳上顯示的條帶大小與預期的載體大小和目的基因大小一致(圖5),并且經測序比對分析后與目的基因序列完全一致,堿基未見缺失或者突變,說明重組質粒構建成功。
2.4 pET-32a-chcGAS表達產物的SDS-PAGE分析
將pET-32a-chcGAS重組質粒轉化到Rosetta(DE3)感受態細胞中后,成功表達出相對分子質量為75 000的目的蛋白質(圖6),分析結果顯示,蛋白質在包涵體和上清液都有表達且主要集中在上清液表達。
2.5 重組蛋白質的Western-blot鑒定
利用Western-blot對蛋白質進一步鑒定分析,結果(圖7)顯示,在PVDF膜上有一條明顯條帶且位置大小與預期目的蛋白質大小一致。說明重組蛋白質可以被抗His標簽的一抗識別。
2.6 chcGAS感染前后亞細胞定位分析
通過在LMH細胞中轉染pCAGGS-HA-chcGAS和pmCherry-Cl-chcGAS來研究chcGAS的細胞定位。結果顯示,未感染FAdV-4時chcGAS主要分布在細胞核的核膜上,感染FAdV-4后,可以觀察到chcGAS從細胞核的核膜向細胞質里轉移(圖8、圖9)。
3 討論
cGAS作為近幾年來發現的一種新型胞質DNA受體,其功能為識別胞質內的DNA。然而雞cGAS基因的相關研究較少,通過Megalign比對分析物種間cGAS核苷酸同源性,結果顯示,chcGAS與其他物種cGAS的同源性為50.5%~84.4%,且具有2個在第3至第39氨基酸和第68至第81氨基酸的低復雜區域和1個高度保守的具有第129至第429氨基酸的Mab-21結構域,表明chcGAS與其他物種具有差異。本研究對chcGAS基因進行了克隆。首先,通過構建pET-32a-chcGAS原核表達載體,其次,經過多種對比分析后從大腸桿菌表達菌株中挑選了Rosetta(DE3)表達菌株,最后以15 ℃、異丙基硫代半乳糖苷(IPTG) 0.5 mmol/L、誘導20 h為最適誘導條件,成功表達了chcGAS蛋白。SDS-PAGE分析結果顯示,重組蛋白質chcGAS主要以可溶性蛋白質的形式在上清液處表達,利用His標簽抗體經Western-blot鑒定,在PVDF膜上顯示有單一的條帶且與預期目的蛋白質條帶大小相符。
在正常情況下DNA只存在于細胞核和線粒體中,但在病毒感染后會游離在細胞質中,隨后被位于細胞質中的cGAS識別,繼而激活cGAS-STING通路發揮抗病毒作用[10]。目前為止報道人和小鼠cGAS調控較多[18],以往的研究中,普遍認為人cGAS僅僅分布在細胞質中[19],并行使DNA識別受體的功能[20-21]。但近期有試驗結果表明,人cGAS定位在細胞膜上,并指出有外源DNA感染細胞時cGAS發生移位即可從細胞膜上轉移到細胞質中從而識別一些DNA而后激活天然免疫。雞cGAS還沒有得到足夠的研究,尤其雞cGAS的細胞定位、功能及精確調控機制還未完全研究清楚。先前Wang等[14]通過在雞胚成纖維細胞中瞬時轉染Flag標記的cGAS來研究雞cGAS的亞細胞定位,定位分析結果表明,chcGAS分布在細胞質中。本研究使用pCAGGS-HA-chcGAS和pmCherry-C1-chcGAS 2個重組質粒在LMH細胞中對chcGAS進行定位,結果顯示,chcGAS是定位在細胞核的核膜上,此外還通過FAdV-4感染chcGAS對細胞定位進行觀察,結果表明,在FAdV-4感染后chcGAS定位可從細胞核的核膜上轉移到細胞質中,后續將通過構建穩定表達chcGAS的LMH細胞系對這一結果進行更加深入地研究驗證。通過chcGAS定位了解其在天然免疫中發揮的作用,同時也為后續研究chcGAS的功能提供了理論依據。
參考文獻:
[1] TAKEUCHI O, AKIRA S. Pattern recognition receptors and inflammation[J]. Cell, 2010, 140(6): 805-820.
[2] KATO K, ISHII R, GOTO E, et al. Structural and functional analyses of DNA-sensing and immune activation by human cGAS[J]. PLoS One, 2013, 8(10): e76983.
[3] MORI A, OLESZYCKA E, SHARP F A, et al. The vaccine adjuvant alum inhibits IL-12 by promoting PI3 kinase signaling while chitosan does not inhibit IL-12 and enhances Th1 and Th17 responses[J]. European Journal of Immunology, 2012, 42(10): 2709-2719.
[4] STEMPEL M, CHAN B, BRINKMANN M M. Coevolution pays off: herpesviruses have the license to escape the DNA sensing pathway[J]. Medical Microbiology and Immunology, 2019,208(3): 495-512.
[5] DAMBUZA I M, BROWN G D. C-type lectins in immunity: recent developments[J]. Current Opinion in Immunology, 2015, 32:21-27.
[6] STEIN S C, LAM E, FALCK-PEDERSEN E. Cell-specific regulation of nucleic acid sensor cascades: a controlling interest in the antiviral response[J]. Journal of Virology, 2012, 86(24): 13303-13312.
[7] ZHOU H, CHEN S, WANG M S, et al. Interferons and their receptors in birds: a comparison of gene structure, phylogenetic analysis, and cross modulation[J]. International Journal of Molecular Sciences, 2014,15(11):21045-21068.
[8] KEATING S E, BARAN M, BOWIE A G. Cytosolic DNA sensors regulating type I interferon induction[J]. Trends in Immunology, 2011,32(12):574-581.
[9] WU J X, SUN L J, CHEN X, et al. Cyclic GMP-AMP is an endogenous second messenger in innate immune signaling by Cytosolic DNA[J]. Science,2013,339(6121):826-830.
[10]SUN L J, WU J X, DU F H, et al. Cyclic GMP-AMP synthase is a cytosolic DNA sensor that activates the type I interferon pathway[J]. Science, 2013,339(6121):786-791.
[11]BARNETT K C, CORONAS-SERNA J M, ZHOU W, et al. Phosphoinositide interactions position cGAS at the plasma membrane to ensure efficient distinction between self- and viral DNA[J]. Cell, 2019,176(6):1432-1446.
[12]楊 潔. 雞源天然免疫DNA感受器cGAS-STING信號軸功能及其抗病毒作用研究[D].揚州:揚州大學,2020.
[13]OLIVEIRA M, RODRIGUES D R, GUILLORY V, et al. Chicken cGAS senses fowlpox virus infection and regulates macrophage effector functions[J]. Front in Immunology, 2021,11. DOI:10.3389/fimmu.2020.613079.
[14]WANG J, BA G, HAN Y Q, et al. Cyclic GMP-AMP synthase is essential for cytosolic double-stranded DNA and fowl adenovirus serotype 4 triggered innate immune responses in chickens[J]. International Journal of Biological Macromolecules, 2020,146:497-507.
[15]ZHANG H W, JIN W J, DING K, et al. Genetic characterization of fowl adenovirus strains isolated from poultry in China[J]. Avian Diseases, 2017,61(3):341-346.
[16]CAVLAR T, DEIMLING T, ABLASSER A, et al. Species-specific detection of the antiviral small-molecule compound CMA by STING[J]. The EMBO Journal, 2013,32(10):1440-1450.
[17]YIN D D, XUE M, YANG K K, et al. Molecular characterization and pathogenicity of highly pathogenic fowl adenovirus serotype 4 isolated from laying flock with hydropericardium-hepatitis syndrome[J]. Microbial Pathogenesis, 2020,147. DOI:10.1016/j.micpath.2020.104381.
[18]DAI J, HUANG Y J, HE X H, et al. Acetylation blocks cGAS activity and inhibits self-DNA-induced autoimmunity[J]. Cell,2019,176(6):1447-1460.
[19]ZHOU Y, HE C, WANG L, et al. Post-translation regulation of antiviral innate signaling[J]. European Journal of Immunology, 2017,47(9):1414-1426.
[20]CIVRIL F, DEIMLING T, DE OLIVEIRA MANN C C, et al. Structural mechanism of cytosolic DNA sensing by cGAS[J]. Nature, 2013,498(7454):332-337.
[21]LI X D, WU J X, GAO D X. et al. Pivotal roles of core-cGAS-cGAMP signaling in antiviral defense and immune adjuvant effects[J]. Science, 2013,341(6152):1390-1394.
(責任編輯:陳海霞)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年18期)2018-11-14 01:48:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
山東工業技術(2016年15期)2016-12-01 05:31:22
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
