張廣才嶺嚙齒動物分散儲食選擇3種林木種子特征研究
2023-06-13 16:46:38李殿偉常詩麒張承志高明曹鈺煒徐夢昊王陽
牡丹江師范學院學報(自然科學版) 2023年2期
李殿偉 常詩麒 張承志 高明 曹鈺煒 徐夢昊 王陽

摘 ?要:為了解東北地區寒溫帶森林嚙齒動物對不同種子特征的選擇規律,2018年和2019年夏季,在張廣才嶺地區采用標簽標記法追蹤調查釋放的紅松(Pinuskoraiensis)、毛榛(Corylus mandshurica)、蒙古櫟(Quercus mongolica)3種林木種子,測定嚙齒動物取食或擴散后的種子特征.調查結果表明:紅松、毛榛、蒙古櫟三種林木種子形態學特征和營養成分具有明顯的差異,這些種子特征的差異可能會影響取食者的偏好和決策.種子大小、種子質量、種仁質量及比值、種皮厚度與硬度、種子的營養物質與其他成分等影響嚙齒動物取食或者儲食的重要因素,嚙齒動物對種子做出的行為,是基于種子特征而做出的綜合評估的結果.
關鍵詞:嚙齒動物;食物選擇;種子特征;種子命運
[ ? 中圖分類號 ? ?]Q958[ ? ?文獻標志碼 ? ] ?A
Selection of 3 Forest Seeds Characteristics of Rodents for
Scattered Storagein Zhang Guangcai Mountains
LI Dianwei*,CHANG Shiqi,ZHANG Chengzhi,GAO Ming,CAO Yuwei,
XU Menghao,WANG yang
(College of Life Science and Technology,Mudanjiang Normal University,Mudanjiang 157011,China)
Abstract:Tounderstand the selection pattern of rodents on different seed In order to understand the selection of different seed characteristics by rodents in cold temperate forests in Northeast China, in the summers of 2018 and 2019, three forest seeds of Pinus koraiensis, Corylus mandshurica, and Quercus mongolica were tracked and investigated by label tagging in Zhang Guangcai mountains and the seed characteristics of rodents after feeding or spreading were determined.The results showed that there were obvious differences in seed morphological characteristics and nutrient composition of red pine, woolly hazel and Mongolian oak seeds, which will have an impact on the decisions of the predators. Rodent's selection of seeds is based on a comprehensive evaluation of various seeds'properties including size, kernel quality, and the specific value between seed and kernel quality, and thickness, hardness, nutrients.
Key words:rodent;food select;seed characteristic;seed fate
在寒溫帶森林中,紅松(P.koraiensis)、毛榛(C.mandshurica)、蒙古櫟(Q.mongolica)植被分布廣泛,均產大型種子,是嚙齒動物主要的食物來源.了解這些常見種子的主要特征以及嚙齒動物對各類種子的選擇特征,是深入開展嚙齒動物儲食生態學研究的基礎,對于探索動、植物之間的互惠關系具有非常重要的意義.
嚙齒動物取食植物的種子和果實,對森林植被造成危害[1] ,但嚙齒動物對種子的儲藏行為,又使其成為植物擴散的有效傳播者.在種子成熟季節,食物資源豐富,許多嚙齒動物會將食物儲藏起來,以應對資源短缺(如冬季).動物再利用[2]會消耗大部分儲藏物,也有很多儲藏種子被遺忘或忽略.當條件適宜時,部分種子會萌發并長成幼苗,成為森林天然更新的重要驅動力之一.[3]嚙齒動物的取食和儲食行為是一個復雜的過程,這是一種進化過程中形成的動植物間的互惠作用,植物通過進化一些特征吸引動物擴散、儲藏其果實或種子.嚙齒動物對種子的選擇或決策受到多種因素的影響.[2,4]種子大小和質量、種皮特征、水分含量、營養物質以及次生代謝化合物等任何單一的種子性狀都不能解釋種子的擴散模式.單粒種子的收益是影響種子命運和擴散距離的關鍵因素[5],因為很難區分種子個體性狀對嚙齒動物取食、儲食行為的影響和它們之間的相互作用[6],種子性狀組合在嚙齒動物取食和儲食行為研究中非常重要.[2]本研究測定實驗種子的基礎特征,標記并投放,了解嚙齒動物對主要林木種子的選擇規律,為嚙齒動物危害防治和森林生態系統保護積累資料.
1 研究地點與方法
1.1 研究地點概況
黑龍江省牡丹江市三道林場林區(44°40? ~ 44°45? N,129°24? ~ 129°32? E,海拔在380 ~ 550 m)位于張廣才嶺主脊東部余脈,地形復雜,多為低山和丘陵,屬于寒溫帶大陸性季風氣候.冬季嚴寒干燥,晝短夜長,年平均氣溫3 ℃,最高氣溫33.6 ℃,最低氣溫-39 ℃,平均積溫2 500 ℃,無霜期130天左右.森林主要以闊葉林、針闊混交林兩種生境為主.牡丹江市三道林場動植物資源豐富,生產大型種子的林木主要有蒙古櫟(Q.mongolica)、紅松(P.koraiensis)、毛榛(C.mandshurica)、山杏(Armeniaca sibirica (L.) Lam)、胡桃楸(Juglans mandshurica)、云杉(Picea asperata Mast.)、冷杉(Abies fabri)等.[7-9]
1.2 研究方法
嚙齒動物調查 采用鋏日法用 2 號鐵板捕鼠鋏(15 cm×8 cm)進行嚙齒動物群落組成調查.每塊樣地按3條樣線布鋏,樣線間隔20 m,鋏距5 m,按照地形地勢特點,每條樣線上布置100個鐵鋏.用炒熟的白瓜子為誘餌,次日(24 h后)查看捕獲情況,補充誘餌,記錄捕獲動物種類、數量.
種子采集與測量 采集成熟的紅松、毛榛、蒙古櫟種子,在常溫下自然風干后儲存、備用.隨機選取種子,測定種仁中蛋白質、脂類、碳水化合物含量(g /100 g).用電子游標卡尺測量種子大小和種皮厚度(±0.01 mm).用電子天平測量種子和種仁質量(±0.01 g),計算種仁/種子質量比值.用數顯維氏硬度儀(HVS502,HRC模式,施加力19.8 N)測量種皮硬度(±0.01 HV).
種子標記與種子命運調查 采用標簽標記法標記種子類別、樣點和種子編號.對投放的種子命運進行調查.種子的命運定義為以下幾種:原地完好(未被取食和搬運)、原地取食(在投放點被取食)、搬運后取食(種子被搬離后取食)、搬運后完好(被搬離后棄在地表)、搬運后埋藏(被搬離后埋藏土壤下)、搬運后丟失(搬離后無法找到)、消耗(除原地完好的種子)、取食(原地取食+搬運后取食)、擴散(搬運后完好+搬運后埋藏+搬運后丟失).
1.3 數據統計與分析
應用Excel對種子的大小(長、短徑)、種皮厚度、種子質量、種仁質量、種皮硬度、種仁/種子質量比值和主要營養物質含量進行數據統計處理與檢驗分析.用SPSS 22.0軟件進行數據分析,計算數據是否有異常值,檢驗數據正態性和方差齊性.符合正態性和方差齊性的數據用參數檢驗,不符合的用非參數檢驗.根據不同研究內容的需要,分別利用T檢驗、Person相關分析、多樣本檢驗、獨立樣本檢驗進行數據檢驗.所有數據統計值用平均值±標準差表示,顯著性水平為α=0.05,極顯著水平為α=0.01.
2 結果與分析
2.1 嚙齒動物群落結構
嚙齒動物群落組成調查共布設1 500鋏日,捕獲嚙齒動物共計156只,捕獲率為10.40%.捕獲的嚙齒動物隸屬于2科3屬4種.鼠科(Muridae)姬鼠屬(Apodemus)大林姬鼠(Apodemus peninsulae)占46.15%,黑線姬鼠(Apodemus agrarius)占39.74%,倉鼠科(Circetidae)?屬(Craseomys)棕背?(Craseomys rufocanus)占13.46%,大倉鼠屬(Tscherskia)大倉鼠(Tscherskia triton)占0.65%.黑線姬鼠與大林姬鼠為研究地點的優勢種.[10]
2.2 種子的基礎特征
2.2.1 種子的形態特征
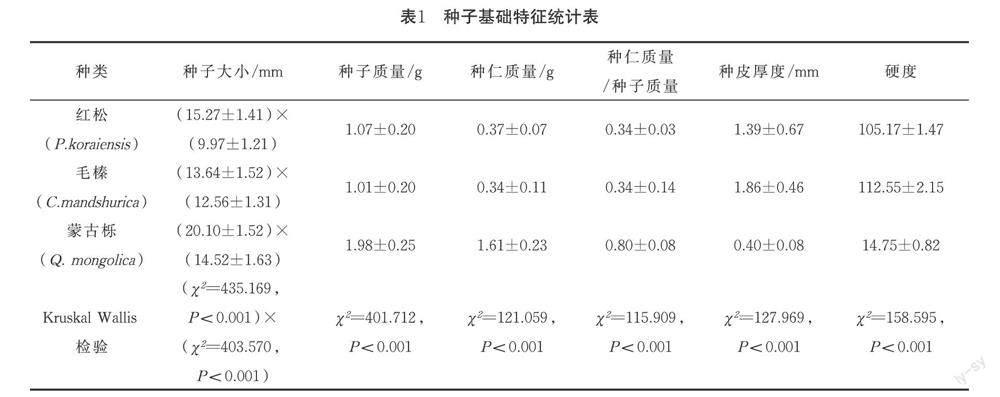
3種林木種子的形態特征差異明顯.蒙古櫟種子大小與質量均最大,其次是毛榛,紅松的最小;蒙古櫟種仁質量最大,其次是紅松,毛榛的最小;蒙古櫟的種仁質量/種子質量比值最大,其次是毛榛,紅松的最小;種皮厚度和硬度均有相同規律,毛榛最大,其次是紅松,蒙古櫟最小.詳見表1.
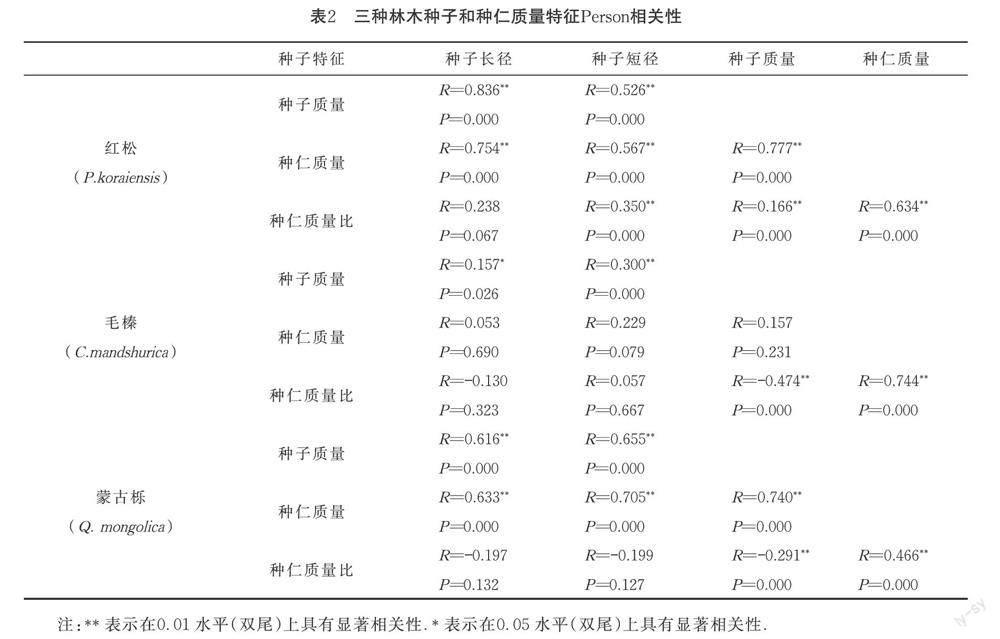
Person相關性檢驗顯示,紅松、毛榛和蒙古櫟種子質量均與種子長、短徑呈不同程度正相關(P<0.001).種仁質量相關性檢驗顯示,僅毛榛的種仁質量與種子長、短徑以及種子質量無顯著相關性,紅松和蒙古櫟的種仁質量均與其種子長徑、短徑以及種子質量表現出正相關(P<0.001).種仁質量比值相關性檢驗表明,三種種子的種仁質量比值與種子質量均表現出顯著的負相關(P<0.001),與種仁質量呈現顯著的正相關(P<0.001).詳見表2.
2.2.2 種子的營養成分
100 g種仁中蛋白質含量:毛榛(20.18%)>紅松(17.33%)>蒙古櫟(6.79%);脂類物質含量:紅松(63.81%)>毛榛(53.89%)>蒙古櫟(40.14%);碳水化合物含量:蒙古櫟(53.07%)>毛榛(25.93%)>紅松(18.86%).
2.3 不同命運的種子特征
2.3.1 消耗種子的形態特征
經過嚙齒動物選擇,不同命運的種子特征差異較大.統計發現,不同種子的形態特征對嚙齒動物選擇種子具有影響.分別對嚙齒動物取食(原地取食、搬運取食)和擴散(搬運后完好、搬運后埋藏)的3種林木種子特征與種子的基礎值進行比較,結果見表3.
紅松:(1)原地取食種子的長徑、短徑和基礎值無差異,質量無差異.(2)搬運取食種子的短徑小于基礎值,長徑、質量與基礎值之間無差異.(3)擴散的紅松種子質量、長短徑與基礎值之間無差異.(4)原地取食和搬運后取食的種子比較,長短徑和質量均無差異.(5)取食種子的長短徑、質量和擴散種子之間無差異.(6)擴散距離小于搬運后取食的距離.
毛榛:(1)原地取食的種子長、短徑大于基礎值,質量無差異.(2)搬運取食種子長短徑大于基礎值,質量無差異.(3)擴散的毛榛種子質量、長短徑與基礎值之間無差異.(4)原地取食和搬運后取食的種子比較,長短徑和質量均無差異.(5)取食種子長短徑大于擴散種子,質量無差異.(6)擴散距離與搬運后取食距離有差異.
蒙古櫟:(1)原地取食的種子短徑和質量大于基礎值,長徑無差異.(2)搬運后取食的種子長短徑和質量與基礎值均無差異.(3)擴散的種子長短徑和質量均小于基礎值.(4)原地取食種子的長短徑和質量與搬運取食均無差異.(5)擴散種子長短徑和質量均小于原地取食,搬運取食種子的長短徑和質量均大于擴散種子.(6)擴散距離與搬運后取食距離有差異.
2.3.2 擴散種子之間的比較
搬運后埋藏與搬運后未埋藏的種子特征比較顯示,3種林木種子的長、短徑和質量均無顯著差異.相關性檢驗結果表明,種子擴散距離與種子長、短徑和種子質量均無相關性.詳見表4
3討 論
種子性狀是影響動物取食、儲食的重要因素,并決定種子的命運,植物種子的性狀能夠吸引動物取食并影響動物儲藏行為[11-12],有利于植物的擴散和更新[12-13].紅松、毛榛、蒙古櫟是研究區域常見的植被,產出的大型種子是嚙齒動物偏好的食物[14-15],這些種子形態特征、營養成分的差異是進化中形成的適應特征,尋找嚙齒動物儲食這些種子的特征規律,有助于理解動植物互惠進化關系.[15-16]
種子大小通常被認為是影響取食的最重要因素,對嚙齒動物取食或搬運種子的行為具有顯著影響.[1-3]本研究中,原地取食種子的長短徑和質量普遍大于搬運取食種子,說明嚙齒動物對種子大小具有的選擇性.也有研究認為,種仁質量比值相對于種子大小會起到更重要的作用.[2-7]種仁質量比決定了種子的單粒收益,種仁質量比值越大表明單粒收益越大.蒙古櫟種仁質量比達到0.84左右,比值最高;紅松種子質量最小,但種仁質量比與毛榛接近,達到0.33左右.
從種皮特征的角度,種皮較厚、較硬的種子處理的時間長,被捕食風險高.[17-20]通常嚙齒動物取食蒙古櫟最多,可能也與其種皮較薄容易處理有關,但是較薄的種皮不利于長期儲藏,保存期間容易發霉.與紅松和蒙古櫟相比,毛榛的種皮厚度更厚、硬度更大,會增加嚙齒動物處理種皮的時間,所以,取食的比例偏低,但被擴散儲藏的比例相對更高.[14-15,21-22]
種子的營養成分是影響動物取食和儲食的關鍵因素.[1-3]種子的脂類和淀粉含量較高時,動物獲得能量收益也會增加.[23-25]紅松、蒙古櫟、毛榛種子之所以成為嚙齒動物偏好的食物,是因為紅松種子的脂類物質含量超過60%,蒙古櫟種子的碳水化合物超過50%、脂類物質超過40%,毛榛種子脂類物質含量超過50%,蛋白質超過20%.嚙齒動物優先取食淀粉和脂肪含量更高的蒙古櫟和紅松,能夠獲得最大的能量收益.[26]
對于同種的種子,種子大小的差異可能是影響動物做出決策的主要因素,但對于多種種子同時進行選擇時,單純的大種子并不是動物的首選,種子的綜合特征在選擇過程中發揮著更大作用.
4 結論
紅松、毛榛、蒙古櫟三種林木種子形態學指標和營養成分具有顯著差異,種子大小和質量、種皮的厚度和硬度、種子的營養物質含量等特征,會影響嚙齒動物對食物選擇的決策,嚙齒動物選擇種子是對多種特征綜合評估的結果,不能僅考慮單一因素的顯著作用.
參考文獻
[1]Vander Wall S B. How plants manipulate the scatter-hoarding behavior of seed-dispersing animals[J]. Philosophical Transactions of The Royal Society B Biological Sciences, 2010, 365(1542):989-997
[2]Lichti N I, Steele M A, Swihart R K. Seed fate and decision-making processes in scatter-hoarding rodents[J]. Biological Reviews, 2015,92(1):474-504.
[3]肖治術, 張知彬. 嚙齒動物的貯藏行為與植物種子的擴散[J]. 獸類學報, 2004, 24(1):61-70.
[4]李殿偉,孟繁星,劉佳慧,等.張廣才嶺東部兩種姬鼠兩性異形比較研究[J].湖北農業科學,2022,61(20):105-110+187.
[5]Luna C A, Loayza A P, Squeo F A, et al. Fruit Size Determines the Role of Three Scatter-Hoarding Rodents as Dispersers or Seed Predators of a Fleshy-Fruited Atacama Desert Shrub[J]. PLOS ONE, 2016, 11(11):e0166824.
[6]Wang B, Chen J. Seed size, more than nutrient or tannin content, affects seed caching behavior of a common genus of Old World rodents.[J]. Ecology, 2009, 90(11):3023-3032.
[7]金志民.牡丹江三道林場嚙齒動物區系[J].天津農業科學,2015,21(03):79-81.
[8]李殿偉,姚旭,金志民,等.張廣才嶺東部山地嚙齒動物群落多樣性調查[J].中國森林病蟲,2019,38(05):28-33+37.
[8]Moore, J. E., McEuen, A. B., Swihart, R. K., Contreras, T. A., & Steele, M. A. Determinants of seed removal distance by scatter-hoarding rodents in deciduous forests. Ecology,2007, 88(10), 2529–2540.
[9] 張 巍,牟長城,屈紅軍.東北林區落葉松人工林群落植物多樣性研究[J].牡丹江師范學院學報:自然科學版,2008(3):1-4
[10] 紀春艷,李強,楊春文.大林姬鼠晝夜活動節律的研究[J].牡丹江師范學院學報:自然科學版,2005(04):1-3.
[11]苗迎權. 基于種子特征的六盤山區嚙齒動物對種子的取食和擴散行為[D]. 銀川:北方民族大學,2016
[12]Vander Wall S B, Jenkins S H. Reciprocal pilferage and the evolution of food-hoarding behavior[J]. Behavioral Ecology,2003,14(5):656-667.
[13]Steele M A. Evolutionary interactions between tree squirrels and trees: a review and synthesis[J]. Current Science,2008,95: 871-876.
[14]李殿偉,姚旭,陳蒙愛,等.黑線姬鼠對林木種子的選擇性[J].中國森林病蟲,2019,38(06):1-5.
[15]Li, D.W., Jin, Z.M., Yang, C.Y., et al. (2018) Scatter-hoarding the seeds of sympatric forest trees by Apodemus peninsulae in a temperate forest in northeast China[J]. Pol. J. Ecol,2018, 66, 382-394.
[16]Dally J M, Clayton N S, Emery N J. The behaviour and evolution of cache protection and pilferage[J]. Animal Behaviour,2006,72(1):13-23.
[17]Stapanian M A, Smith C C. A model for seed scatter-hoarding: Coevolution of fox squirrels and black walnuts [J] .Ecology,1978,59:884-898
[18]Chang G, Xiao Z, Zhang Z. Effects of burrow condition and seed handling time on hoarding strategies of Edward's long-tailed rat (Leopoldamys edwardsi)[J]. Behavioural Processes, 2010,85(2):163-166.
[19]Xiao Z, Zhang Z. Nut predation and dispersal of Harland Tanoak lithocarpus harlandii, by scatter-hoarding rodents[J]. Acta Oecologica, 2006, 29(2):205-213.
[20]Xiao Z, Wang Y, Harris M, et al. Spatial and temporal variation of seed predation and removal of sympatric large-seeded species in relation to innate seed traits in a subtropical forest, Southwest China[J]. Forest Ecology and Management, 2006, 222(1-3):46-54.
[21]Xiao Z, Chang G, Zhang Z. Testing the high-tannin hypothesis with scatter-hoarding rodents: experimental and field evidence[J]. Animal Behaviour, 2008, 75(4):1235-1241.
[22]Li, D.W., Hao, J.W., Yao, X., et al. Observations of the foraging behavior and activity patterns of the Korean wood mouse, Apodemus peninsulae, in China, using infra-red cameras[J]. ZooKeys,2020,992:139-155.
[23]Li, D.W., Liu, Y., Shan, H.J., et al. Effects of season and food on the scatter-hoarding behavior of rodents in temperate forests of Northeast China[J]. ZooKeys,2021,1025,73-89.
[24]Vander Wall S B. The evolutionary ecology of nut dispersal[J]. The Botanical Review,2001,67(1):74-117.
[25]Wang B,Ye C X,Cannon C H,et al. Dissecting the decision making process of scatter-hoarding rodents[J].Oikos,2013,122(7):1027-1034.
[26]Wang B.,Chen J. 2012-Effects of fat and protein levels on foraging preferences of tannin in scatter-hoarding rodents[J]. PLoS ONE2012,7: e40640.
編輯:琳莉
