川滇榿木4 種氣味活性物質對云南切梢小蠹胚后發育的影響
2023-06-20 12:27:28錢路兵張夢蝶劉金林李宗波
西南林業大學學報 2023年3期
錢路兵張夢蝶劉金林李宗波,3
(1. 西南林業大學生物多樣性保護院,云南省森林災害預警與控制重點實驗室,云南 昆明 650233;2. 西南大學植物保護學院,重慶 400715;3. 西南山地森林資源保育與利用教育部重點實驗室,云南 昆明 650233)
云南切梢小蠹(Tomicus yunnanensis)是一種以云南松(Pinus yunnanensis)為寄主的蛀干害蟲,該蠹蟲自20 世紀80 年代在云南中部大爆發以來,現已擴散至四川、貴州、廣西等云南松(Pinus yunnanensis)的原生分布區[1-2],嚴重威脅我國西南地區云南松松林生態系統的安全及生物多樣性。云南切梢小蠹1 年發生1 代,具有干轉梢、梢轉梢和梢轉干3 個獨特的生活時期。其中,在松梢中補充營養物質,完成生殖器官的成熟,并會導致云南松的生長勢衰弱;在樹干韌皮部進行交配、產卵,并完成胚后發育,最終致使樹木死亡[2]。盡管云南切梢小蠹會有一個短暫的轉移期,但其90%以上的生活史均在樹木內部度過,隱蔽的生活方式決定了云南切梢小蠹的防治難度大。目前,營造針闊混交林被認為是防治云南切梢小蠹最為有效的方式。在混交林生態系統中,非寄主林木與寄主林木之間常因時間和空間的持續變化,形成獨特的空間分布與配置的關系,一方面能夠增加物種的多樣性,增強物種之間的互作關系和制衡能力(如天敵、捕食者、寄生蜂),從而降低害蟲的危害程度和發生頻率[3];另一方面在于非寄主林木釋放的特征氣味譜,可直接影響或干擾害蟲的多種行為[4-5]。如綠葉揮發物能夠大幅度地降低云南切梢小蠹、南部松大小蠹(Dendroctonus frontalis)、中穴星坑小蠹(Pityogenes chalcographu)對聚集信息素和寄主氣味的趨向性,影響其寄主識別能力[5-7]。樹皮揮發物苯甲醇、苯甲醛、愈創木酚、鄰羥基苯甲醛以及7-甲基-1,6-二氧螺環[4-5]癸烷(又稱conophthorin)可干擾害蟲產卵地的選擇,延長求偶時間,縮短交配持續期,也會增加天敵的非致死效應[4,8-10]。并且,非寄主林木氣味也是混交林氣味的重要組成部分,且這些氣味物質無時不有,無處不在,很可能作為一種間接的環境脅迫因子[5,11]。相較于其他害蟲,云南切梢小蠹的主要生活史均在樹干的坑道內完成,生活環境也相對封閉,推測這些無時不在的非寄主氣味,特別是活性氣味物質,很可能作為一種非致死效應而直接影響云南切梢小蠹的發育和存活幾率。因此,采用前期篩選的非寄主林木—川滇榿木(Alnus ferdinandi-coburgii)的氣味活性物質,用生測法持續干擾云南切梢小蠹的胚后發育,以驗證非寄主林木氣味對害蟲繁殖機能的影響,并為深入理解混交林抑制害蟲種群數量提供參考。
1 材料與方法
1.1 云南切梢小蠹的胚后發育及形態特征
云南切梢小蠹的生殖器官發育成熟后,于當年10 月至翌年3 月開始蛀入樹干,待雌雄兩性在交配室內交配后,雌蟲即沿樹干縱向方向蛀食,并將卵產于母坑道左右兩側(圖1a)。卵呈橢圓形,長(1.02 ± 0.11)mm,寬(0.63 ± 0.05)mm,乳白色。卵經歷5~12 d 便會孵化為幼蟲,幼蟲沿著與母坑道垂直的方向進行取食,形成子坑道,每個幼蟲均在自己的坑道內發育;幼蟲體稍微彎曲,粗而多皺紋,有3 齡。其中,1 齡幼蟲呈淡黃褐色,頭殼寬0.2~0.6 mm;2 齡幼蟲為黃褐色,頭殼寬0.64~0.90 mm;3 齡幼蟲為橙黃色,頭殼寬0.92~1.28 mm[12]。1.5~2 個月后,幼蟲就會在坑道末端的蛹室內化蛹,新蛹呈乳白色,隨后復眼、口器和翅逐漸變成黑褐色。蛹經歷20~25 d 后,逐漸羽化為成蟲,新成蟲呈淡黃褐色,鞘翅末端2/3 處最先開始出現黑色斑塊,隨后整體呈黑褐色(圖1b)。成蟲在蛹室內待4~7 d 后,則會從蛹室上方咬開羽化孔爬出樹干,尋找合適的嫩梢補充營養,開始新的生命循環。
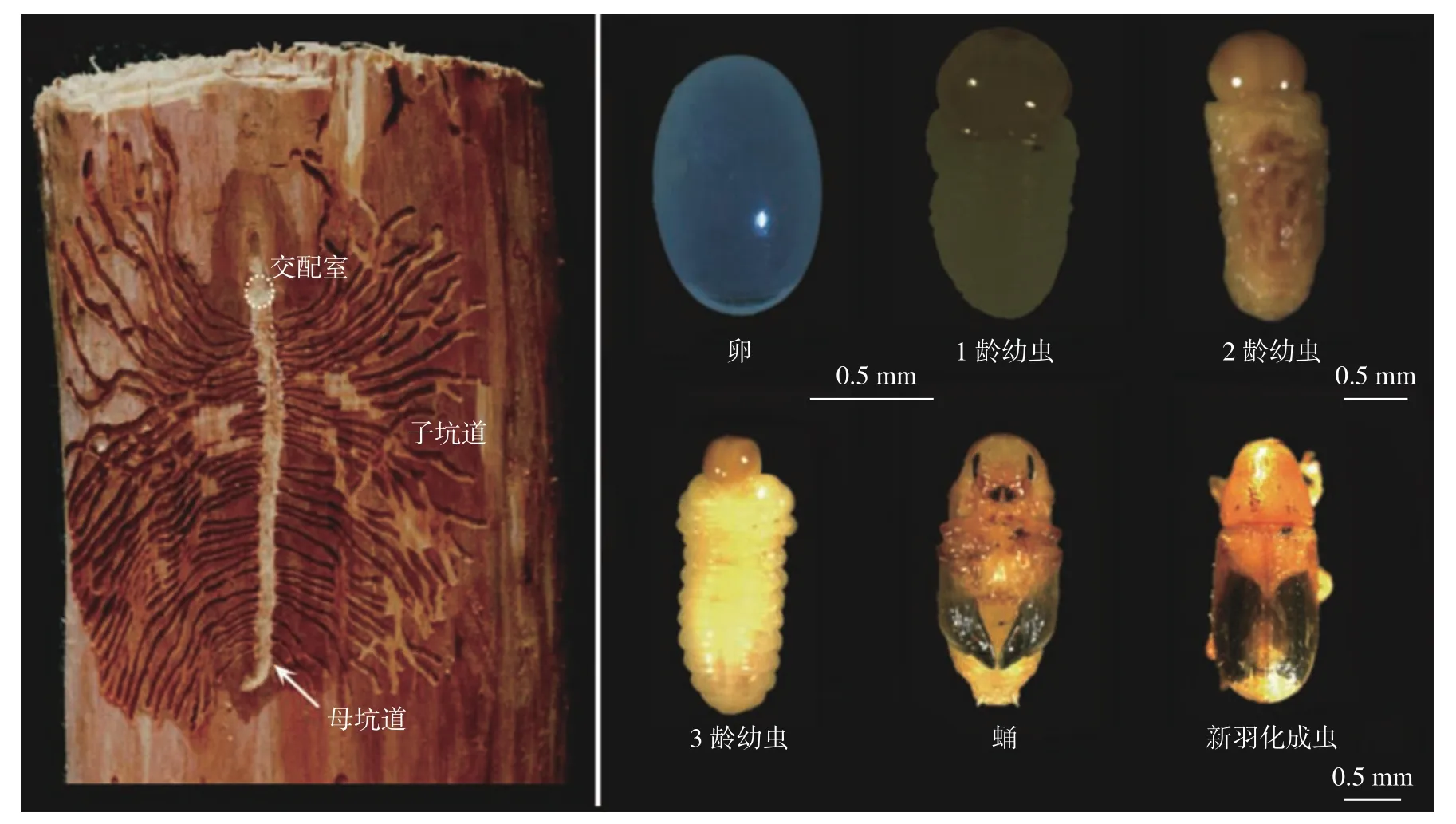
圖 1 云南切梢小蠹的坑道特征和胚后發育Fig. 1 Single maternal gallery and post-embryonic development of T. yunnanensis
1.2 氣味活性物質的篩選與來源
前期在抗云南切梢小蠹混交樹種的篩選過程中,已發現用川滇榿木營造的針闊混交林,能夠很好地削弱云南切梢小蠹的危害,隨后用昆蟲電生理和行為選擇實驗篩選出對乙基苯乙酮、反-2-己烯-1-醇、順-4-己烯-1-醇和4-烯丙基苯甲醚對云南切梢小蠹有趨避活性[13]。干擾實驗使用的4 個化合物以及對照試劑,均從商業公司購買的標準化合物,見表1。
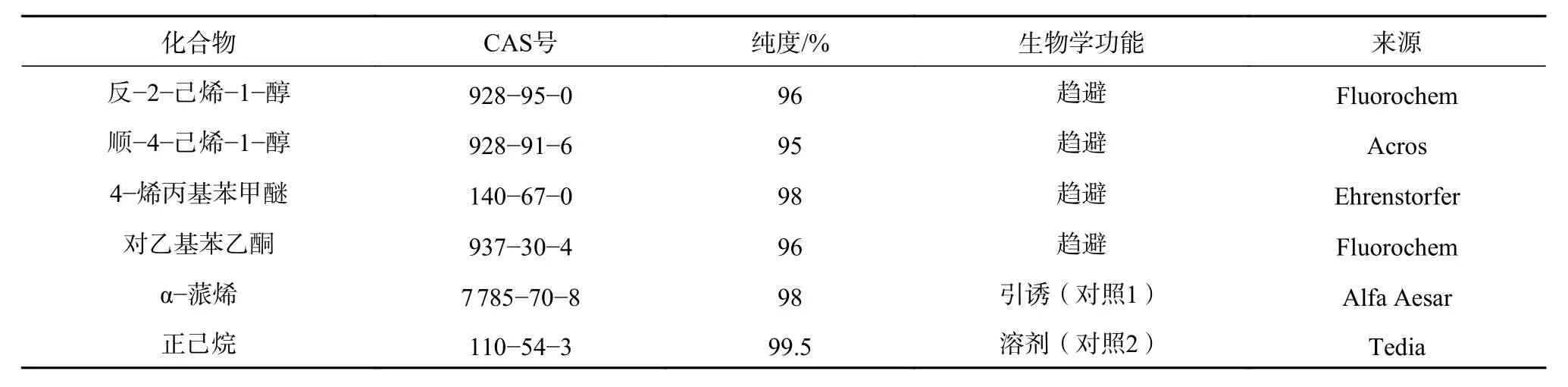
表 1 干擾用的活性氣味物質及來源Table 1 Description of odor-active compounds employed in stressing experiments for T. yunnanensis
1.3 氣味活性物質的干擾實驗
將從云南松松林采集的、生殖器官已發育成熟的云南切梢小蠹帶回實驗室,先根據腹部背板形態以及是否可見第8 節的特征區分雌雄,然后按照雌、雄各15 只的方式放在廣口玻璃瓶(高50 cm,直徑25 cm,體積20 L)中。瓶內先放置2 根新鮮的云南松木段,木段長30 cm,直徑10 cm,且兩端用液體石蠟封閉保濕,用紗布封住瓶口,以便于云南切梢小蠹的侵入和繁殖。待見到木段表面或瓶底有木屑后,意味著雌雄已完成了交配,雌蟲開始蛀坑產卵,隨即將內含活性氣味物質(0.45 ng/μL,溶劑為正己烷)的緩釋瓶置于廣口瓶瓶底,每隔15 d 更換1 次。每個化合物設置3 次重復,并以溶劑(正己烷)和對小蠹蟲具有較強引誘效果的寄主活性成分(α-蒎烯)為對照[14]。最后,將處理好的廣口瓶放置在養蟲室內進行飼養,每隔15 d 各取一木段進行解剖,收集云南切梢小蠹幼蟲(1~3 齡)、蛹及新羽化成蟲(圖1b),將其置于蔡氏體式鏡下拍照(Ste-REO Discovery V20, Geman),并用微量天平測量其體質量。
1.4 數據處理與分析
將體視鏡拍攝的照片導入ImageJ(Version 1.53c, http://imagej.nih.gov/ij)軟 件 中,分 別 對 云南切梢小蠹的幼蟲、蛹、成蟲、頭殼以及正常羽化木段內的母坑道和子坑道等指標進行量化,再根據卵室的數量判斷雌蟲的產卵量。隨后,采用單因素方差分析或者Kruskal-Wallis 檢驗分析氣味活性物質對云南切梢小蠹胚后發育的影響程度。所有的數據分析均在開源軟件Rstudio 中(R Version 4.0.5, https://cran.r-project.org/)完 成,作圖使用GraphPad Prism(Version 8.0.2)。
2 結果及分析
2.1 氣味脅迫對雌蟲產卵量與蛀食坑道的影響
與對照相比,云南切梢小蠹在4 種非寄主氣味脅迫下的產卵量沒有顯著的變化,產卵量均在50~60 粒(表2)。但是,雌成蟲在產卵過程中蛀食的母坑道有顯著的增長(P<0.05),尤其是對乙基苯乙酮、順-4-己烯-1-醇和4-烯丙基苯甲醚,母坑道的長度分別增加了60.47%、55.68%和36.84%,但卵與卵之間的間距沒有明顯的變化。當卵孵化后,幼蟲在子坑道內取食,4 種氣味活性物質均能顯著增加幼蟲蛀食的坑道長度(P<0.05)。其中,4-烯丙基苯甲醚增加的最多,達到47.91%,而順-4-己烯-1-醇增加比例最低,僅為13.07%(表2)。
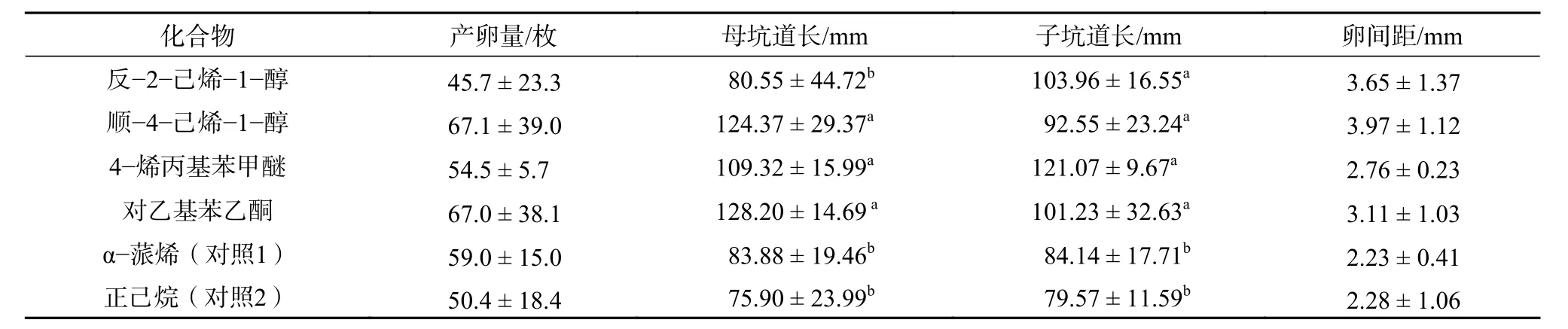
表 2 氣味活性物質對云南切梢小蠹產卵與坑道的影響Table 2 Responses of egg production and maternal gallery of T. yunnanensistressed by the odor-active compounds
2.2 氣味脅迫對幼蟲的影響
在幼蟲發育階段,4 種非寄主氣味活性物質均能夠顯著增加幼蟲各發育階段的體長(P<0.01),以反-2-己烯-1-醇的增加得最多,其次為對乙基苯乙酮,順-4-己烯-1-醇和4-烯丙基苯甲醚增加的最少(表3、圖2)。并且,反-2-己烯-1-醇、對乙基苯乙酮和4-烯丙基苯甲醚對幼蟲的影響效果一致,即從1 齡幼蟲至3 齡幼蟲呈先高后低的趨勢,而順-4-己烯-1-醇則為先低后高的趨勢(圖2)。與對照相比,4 種氣味活性物質均能夠增加幼蟲的體質量,但并沒有顯著差異(表3)。

表 3 氣味活性物質對云南切梢小蠹的幼蟲發育的影響Table 3 Responses of larval development of T. yunnanensisstressed by the odor-active compounds
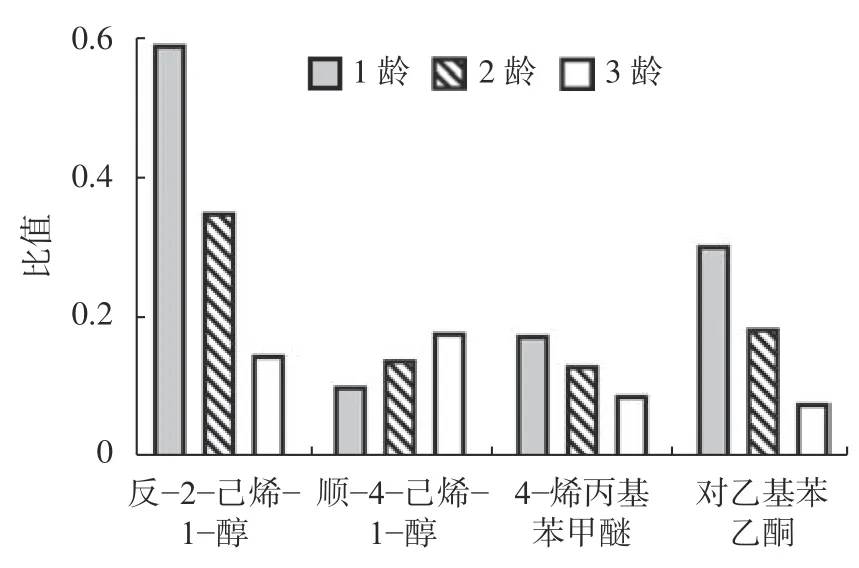
圖 2 氣味活性物質對云南切梢小蠹體長的相對增加比例的影響Fig. 2 Relative percentage of lengthened body length ofT. yunnanensislarvae stressed by the odor-active compounds
2.3 氣味脅迫對蛹和新羽化成蟲的影響
與對照相比(圖3),4 種氣味物質均能顯著降低云南切梢小蠹蛹(P<0.01)和新羽化成蟲的體長(P<0.01),以蛹的效果最好,順-4-己烯-1-醇、對乙基苯乙酮和4-烯丙基苯甲醚的降幅分別為16.63%、14.45%和13.68%,而反-2-己烯-1-醇則為6.95%。在體質量方面,4 種氣味活性物質干擾效果并不一致,反-2-己烯-1-醇、順-4-己烯-1-醇和4-烯丙基苯甲醚能夠顯著地降低蛹(P<0.01)和新羽化成蟲的體質量(P<0.01),以反-2-己烯-1-醇效果最佳,前者降幅為20.09%,后者降幅為15.78%;但對乙基苯乙酮則會小幅度增加蛹和成蟲的體質量,但效果并不明顯。
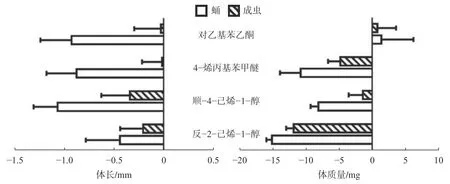
圖 3 氣味活性物質對云南切梢小蠹蛹與新羽化成蟲的影響Fig. 3 Responses of pupa and newly eclosed adults of T. yunnanensisstressed by the odor-active compounds
3 結論與討論
四種川滇榿木氣味活性物質均能明顯干擾云南切梢小蠹的胚后發育,特別是綠葉氣味成分—反-2-己烯-1-醇和順-4-己烯-1-醇能夠直接降低蛹和成蟲的個體大小和體質量,進而影響云南切梢小蠹后續的寄主識別過程及其存活能力,為深入理解針闊混交林降低害蟲種群數量提供了另一例證。
獲取食物是物種繁衍后代的本能需求。然而,其取食活動除了受到自身生理狀態影響外,還會受到各種外界因素的影響。如氣溫變暖引起中歐山松大小蠢(D. ponderosae)、黃杉大小蠹(D. rufipennis)的種群數量大爆發[15-16]、地球磁場改變導致褐飛虱(Nilaparvata lugens)新羽化雌、雄成蟲的體質量顯著下降(14.67%)和(13.17%)[17]、苯并二唑衍生物能夠直接抑制地中海實蠅(Ceratitis capitafa)保幼激素與受體的結合[18]、種間競爭聲信號能夠降低南部松大小蠹母坑道長度,并會增加成蟲羽化死亡率[19]。對小蠹蟲而言,尤其是針葉樹皮小蠹,在坑道內取食是其獲取營養物質和能量的唯一來源,其產卵量亦會隨著坑道長度的增加而增加[2]。在本研究中,4 種氣味活性物質脅迫能夠增加云南切梢小蠹的母坑道長度,但平均產卵量和卵間距沒有顯著的變化,這可能與其在生殖上所分配的能量是恒定的有關,生殖前增加或減少自身營養并不會影響其對生殖所分配的能量,如饑餓并不會影響中歐山松大小蠢的產卵量[20]。另一種可能就是氣味化合物抑制了雌成蟲的能量轉化效率,如合成卵黃原蛋白或卵母細胞受體[18,21],使其必須通過攝取更多的食物才能獲取與對照組等量的營養,幼蟲期既是一個明顯的例證,也是云南切梢小蠹對不良環境的一種生理響應,類似于生活在南北兩極的昆蟲為抵抗低溫而擁有更高的代謝速率、歐洲葡萄蛾(Lobesia botrana)幼蟲在溫度增加時會增加取食來補充自身的能量[22]。Huber 等[5]發現反-2-己烯-1-醇、順-4-己烯-1-醇、1-乙烯基己醇等綠葉氣味能夠降低中歐山松大小蠢、西部松大小蠹(D. brevicomis)、黃杉大小蠹幼蟲的取食量,這與本研究的結果并不一致,可能暗示著通用的綠葉氣味物質的干擾性質不同,正如反-2-己烯-1-醇干擾效果呈下降趨勢,而順-4-己烯-1-醇呈上升趨勢(圖2)。另一方面,大量的研究已經證明昆蟲激素存在低促高抑的現象,昆蟲對不同濃度的同種化合物同樣存在較大的差異[2,23]。在蛹期,受化合物干擾后的云南切梢小蠹的體長及體質量均出現不同程度的減少,這可能因它們幼蟲期擁有較快的代謝導致蛹期消耗更多的能量,從而導致了該階段以及新羽化成蟲的體長和體質量減少[20-21,23]。
混交林作為有害生物生態調控的重要措施之一,已被證明能夠有效抑制害蟲的種群數量,目前已有資源集中假說、寄主可見性假說、天敵假說等對其進行解釋[3-4],但本研究發現非寄主林木氣味能夠降低害蟲的繁殖機能,尤其是蛹和成蟲,將對其以后的存活、繁衍,甚至種間的互作和級聯關系等產生明顯的影響。如昆蟲的體長和體質量常作為衡量其繁殖力的一個重要特征。紅脂大小蠹在求偶的時候,雌蟲更偏愛體型更大的雄性,因為體型大意味著優良的后代;同樣的,雄蟲也偏愛選擇體型更大的雌蟲,因為體型越大意味著能夠產生更多的后代[24-25]。并且,在雄性間的競爭中,體型大小也是一個決定性因素,往往個體較大者容易在競爭中獲勝,容易實現自我繁殖適合度[26-27]。此外,個體小,體質量輕的個體,容易在擴散遷飛和寄主識別過程中死亡,或者被天敵或捕食者殺死[2,28-29],這可能是混交林抑制害蟲種群數量的重要因素之一。對于云南切梢小蠹來說,考慮到不同坑道間幼蟲發育程度,以及坑道相互交錯而致使部分幼蟲缺少食物而死亡(死亡率約為39.85%)[2],非寄主氣味干擾后幼蟲取食量增加可能會進一步加劇種內競爭。因此,在其種群密度較高時,未來是否可用非寄主氣味持續干擾來增加云南切梢小蠹的死亡率有待研究。
