GADD45A 通過端粒替代延長途徑調控骨肉瘤細胞增殖
2023-06-30 09:26:08韓鑫宇李婷芳王峰
天津醫科大學學報 2023年3期
關鍵詞:實驗
韓鑫宇,李婷芳,王峰
(天津醫科大學基礎醫學院醫學遺傳學系,天津 300070)
骨肉瘤是一種起源于類骨質組織,常見于兒童和青少年的惡性骨腫瘤,具有極強的侵襲性并且常常在早期發生轉移,主要涉及股骨、脛骨和肱骨[1-2]。端粒延長是腫瘤生長所必需的關鍵過程,不同于絕大多數腫瘤通過激活端粒酶來維持端粒,以骨肉瘤為代表的10%~15%的人類腫瘤通過端粒延長替代途徑(alternative lengthening of telomeres pathway,ALT)來維持端粒。使用ALT 延長端粒的腫瘤細胞具有獨特的ALT 相關表型,例如含有端粒DNA 的大型早幼粒細胞白血病體(promyelocyte leukemia,PML),稱為ALT 相關PML 小體(ALT-related PML bodies,APBs)以及染色體外端粒DNA 環(C-circle)等[3-4]。
生長停滯和DNA 損傷誘導蛋白45a(GADD45A)定位于細胞核,其水平受周期調控,在G1 期最高而S期顯著降低。GADD45A 在細胞應激反應中起重要作用[5-6]。此外還有研究表明,GADD45A 參與了小鼠干細胞端粒DNA 損傷反應[7]。GADD45A 在細胞周期進展和停滯、DNA 修復和表觀遺傳修飾等多方面的作用與ALT 陽性腫瘤頻繁的DNA 損傷等特點高度相關[7-8]。因此本文旨在探討GADD45A 對骨肉瘤細胞生長、端粒功能和端粒延長替代途徑的影響。
1 材料和方法
1.1 實驗材料 人骨肉瘤細胞U2OS 為本實驗室保存,DMEM 培養基(美國Corning 公司),Opti-MEM培養基、胰蛋白酶(美國Gibco 公司),胎牛血清(烏拉圭Lonsera 公司),ChamQ Mix、逆轉錄試劑盒、dNTP Mix(南京諾維贊生物科技有限公司),DNA 提取試劑盒(Biomiga 倍沃醫學科技有限公司),RNA提取試劑盒(美國Promega 公司),無水乙醇、無水甲醇、異丙醇、冰乙酸(天津康科德生物化工公司),4%多聚甲醛、1 mol/L Tris-HCl(pH7.2)、牛血清白蛋白(BSA)、6×DNA loading buffer(北京索萊寶生物技術公司),國產甲酰胺、秋水仙素(上海生工生物技術股份有限公司),進口甲酰胺、TritonX-100、TWEEN-20、Gelatin from cold water fish(美國Sigma 公司),Blocking Reagent、DIG Easy Hyb Granules、CD-Star Ready-to-use solution、Anti-DIG-AP 抗體(瑞士Roche公司),TelC-FITC PNA 探針488/CY3(韓國Panagene公司),DAPI、Anti-γH2AX 抗體(德國Millipore 公司),Anti-PML 抗體(C-5)(Santa Cruz),Alexa 555 goat anti-Mouse 抗體、Lipofectamine RNAi-Max(美國Thermo 公司),siRNA(蘇州吉瑪基因有限公司),Phi 29 DNAPolymerase(美國NEB 公司),DNase/RNase free water(天根生化科技有限公司),細胞增殖-毒性檢測試劑盒CCK-8(蘭杰柯科技有限公司)。
1.2 實驗方法
1.2.1 細胞培養 將U2OS 細胞接種于10 cm 培養皿中,以DMEM 培養基(含10%胎牛血清)培養于37℃,5%CO2的培養箱中,待細胞密度達70%~80%時進行傳代。
1.2.2 siRNA 轉染 將U2OS 細胞接種于6 孔板中,待細胞生長至密度為40%時進行轉染。取兩支離心管加入Opti-MEM 后分別加入siRNA 和Lipofectamine RNAi-Max,震蕩混勻后將兩管混合,室溫靜置5 min 后逐滴加入對應細胞中,反應24 h 后棄去轉染液每孔加入2 mL 培養基,48 h 后收取細胞用于后續實驗。對應siRNA 序列見表1。
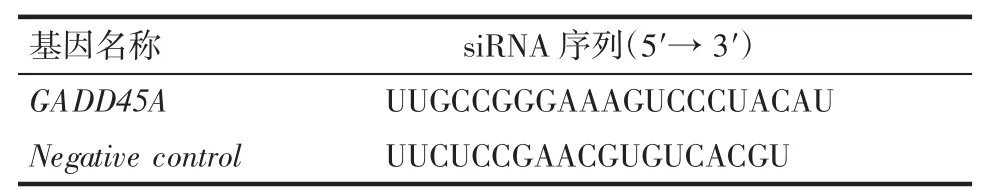
表1 目的基因siRNA 序列Tab 1 siRNA sequences of the target gene
1.2.3 穩定敲低細胞系構建 使用pLKO.1 質粒構建shGADD45A 敲低細胞系。shRNA 序列見表2。用293T 細胞對質粒進行包被,吸取過濾病毒上清感染U2OS 細胞構建穩定敲低細胞系。
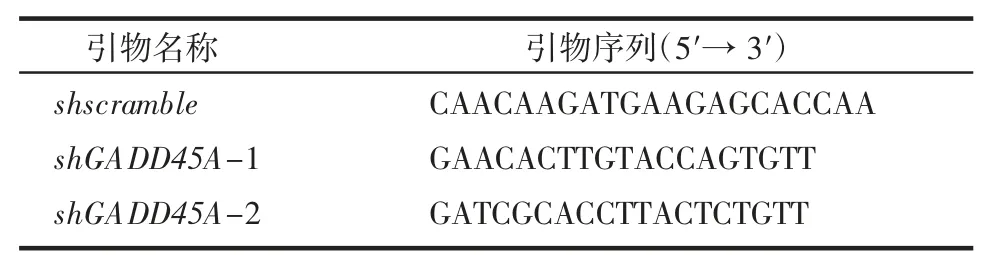
表2 shRNA 序列Tab 2 Sequence of shRNA
1.2.4 實時熒光定量PCR 收取細胞,試劑盒提取RNA。測量RNA 濃度后根據逆轉錄試劑盒將1 μg RNA 逆轉合成cDNA,稀釋5 倍后用于后續實時熒光定量PCR。按照primer F 0.4 μL、primer R 0.4 μL、ChamQ Mix 10 μL、無核酸酶水7.2 μL 的體系按照程序進行擴增(表3)。根據熒光曲線的ct 值用2-ΔΔCt計算siRNA 敲低效率。
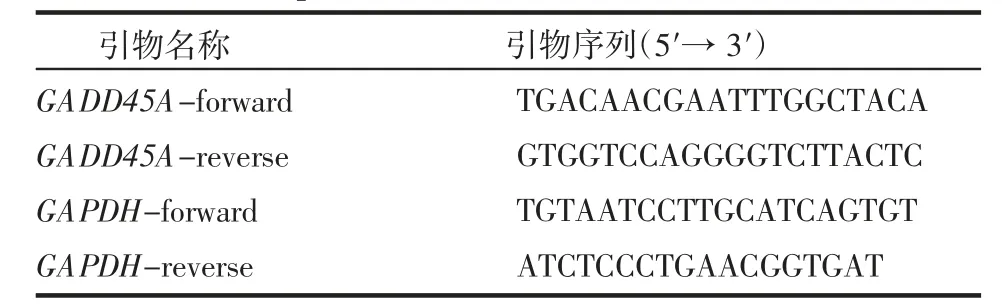
表3 PCR 引物序列Tab 3 Primer sequence for PCR
1.2.5 CCK-8 增殖實驗 牛鮑汁計數板計數U2OS細胞,96 孔板每孔接種1 000 個細胞,每個處理組設置6 組重復。分別測量轉染后第1、3、5、7 天的450 nm 處吸光度值。
1.2.6 克隆形成實驗 牛鮑汁計數板計數U2OS 細胞,6 cm 皿接種5 000 個細胞,每3 天換1 次液,第7 天收取培養皿。吸棄培養基并用超純水潤洗,然后加4%組織固定液固定20 min。洗凈固定液后加入0.5%結晶紫染液,染色20 min。洗凈多余染液,在吸水紙上倒扣瀝干水分,拍照保存用于后續分析。
1.2.7 細胞中期分裂相-熒光原位雜交(Metaphase-FISH) 直接接種穩定敲低細胞或將U2OS 接種于6 cm 培養皿,進行siRNA 轉染,收取細胞前10 h更換含秋水仙素的DMEM 培養基(秋水仙素∶培養基=1∶100)。細胞沉淀加入PBS 重懸,邊震蕩邊加入10 mL 37℃預熱的0.075 mol/L Kcl,37℃水浴孵育30 min,然后加入5 滴預冷固定液(甲醇∶冰乙酸=3∶1),室溫靜置5 min,900 r/min 離心8 min。吸棄上清液,邊震蕩邊加入10 mL 預冷固定液,室溫靜置30 min,900 r/min 離心8 min。吸棄上清液,加1 mL 固定液重懸并取50 μL 滴片,靜置1 晚。第2 天用4%多聚甲醛固定細胞,脫水后用TelC-FITC PNA 探針CY3 雜交端粒3 h,洗滌封片,-20℃保存用于后續拍攝統計。
1.2.8 免疫熒光-熒光原位雜交(IF-FISH) 將無菌蓋玻片放入六孔板中,直接接種穩定敲低細胞或接種U2OS 細胞并進行siRNA 轉染。收取蓋玻片洗滌固定,脫水后用TelC-FITC PNA 探針488 雜交端粒3 h,PBG 封閉1 h(PBS∶BSA∶Gelatin=1∶1∶100),加入一抗室溫孵育1 h(PML∶PBG=1∶1 000,γH2AX∶PBG=1∶1 000),洗滌后二抗室溫孵育30 min(Alexa 555 goat anti-Mouse∶PBG=1∶1 000),DAPI 封片,熒光顯微鏡拍攝并進行統計。
1.2.9 C-circle 實驗 收取細胞提取DNA,測濃度后稀釋至30 ng/μL 并按DNA template 1 μL、BSA 0.4 μL、dNTP Mix 2 μL、Phi 29 DNA Polymerase 0.5 μL、Phi 29 buffer 0.4 μL、無核酸酶水14.1 μL的體系進行PCR 反應擴增,反應程序為30℃,8 h;65℃,20 min。將擴增產物進行斑點雜交,紫外交聯固定后雜交過夜。第2 天回收雜交液并洗膜、封閉,孵育DIG 抗體8~10 h。孵育結束后洗膜最后用CDStar 顯色液曝光。
1.3 統計學處理 采用GraphPad Prism6 和Image J 軟件進行統計學分析和作圖,所有實驗數據為重復3 次實驗結果。統計結果采用t 檢驗,P<0.05 為差異有統計學意義。
2 結果
2.1 GADD45A 對骨肉瘤細胞生長增殖的影響 實時熒光定量PCR 檢測siRNA 和shRNA 對GADD45A的敲低效率。與對照組相比,siRNA 敲低GADD45A后mRNA 水平顯著降低(t=25.96,P<0.000 1);與shscramble 組相比,shGADD45A-1 和shGADD45A-2兩細胞系GADD45A 的mRNA 水平下降(t=21.12、18.37,均P<0.000 1)(圖1)。CCK-8 增殖實驗結果表明,在siRNA 敲低GADD45A 后或穩定敲低細胞系shGADD45A-1 和shGADD45A-2 中,與對照組相比,骨肉瘤細胞存活率明顯下降(t=5.051、6.192、3.775、14.86、22.93、3.013、14.61、20.93,均P<0.05)(圖2A、2B)。克隆形成實驗結果表明,在siRNA 敲低GADD45A 后以及shGADD45A-1 和shGADD45A-2兩組穩定敲低細胞系中,骨肉瘤細胞增殖能力受到明顯抑制,克隆形成減少(t=46.68、23.73、24.98,均P<0.000 1),見圖2C~2D。
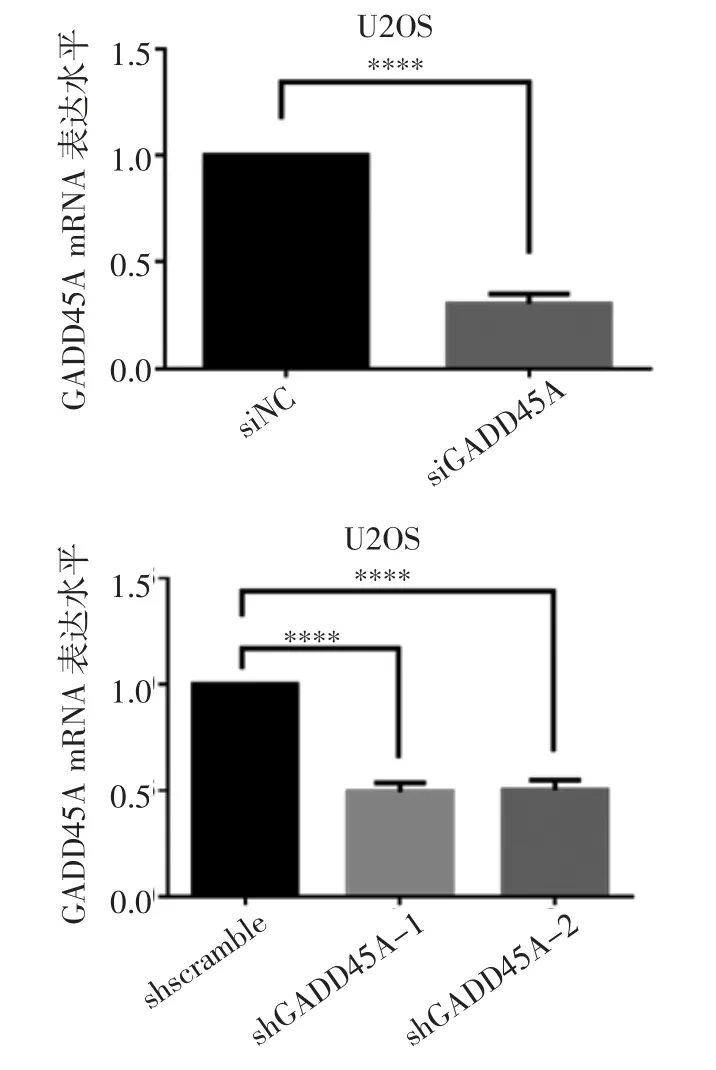
圖1 實時熒光定量PCR 檢測GADD45A mRNA 水平Fig 1 Expression of GADD45A mRNA detected by real-time fluorescence quantitative PCR

圖2 GADD45A 對骨肉瘤細胞增殖的影響Fig 2 The effect of GAD45A on the proliferation of osteosarcoma cells
2.2 GADD45A 對骨肉瘤端粒功能的影響 細胞中期分裂相-熒光原位雜交實驗顯示,siRNA 敲低GADD45A 與shGADD45A-1 和shGADD45A-2 穩定敲低處理后結果相同,染色體末端多端粒信號(Multiple Telomere Signals,MTSs)比例明顯升高(t=24.04、7.243、27.93,均P<0.01),端粒信號丟失(Telomere Signal Free Ends,SFEs)頻率也相應增加(t=8.222、16.61、6.781,均P<0.01)(圖3)。
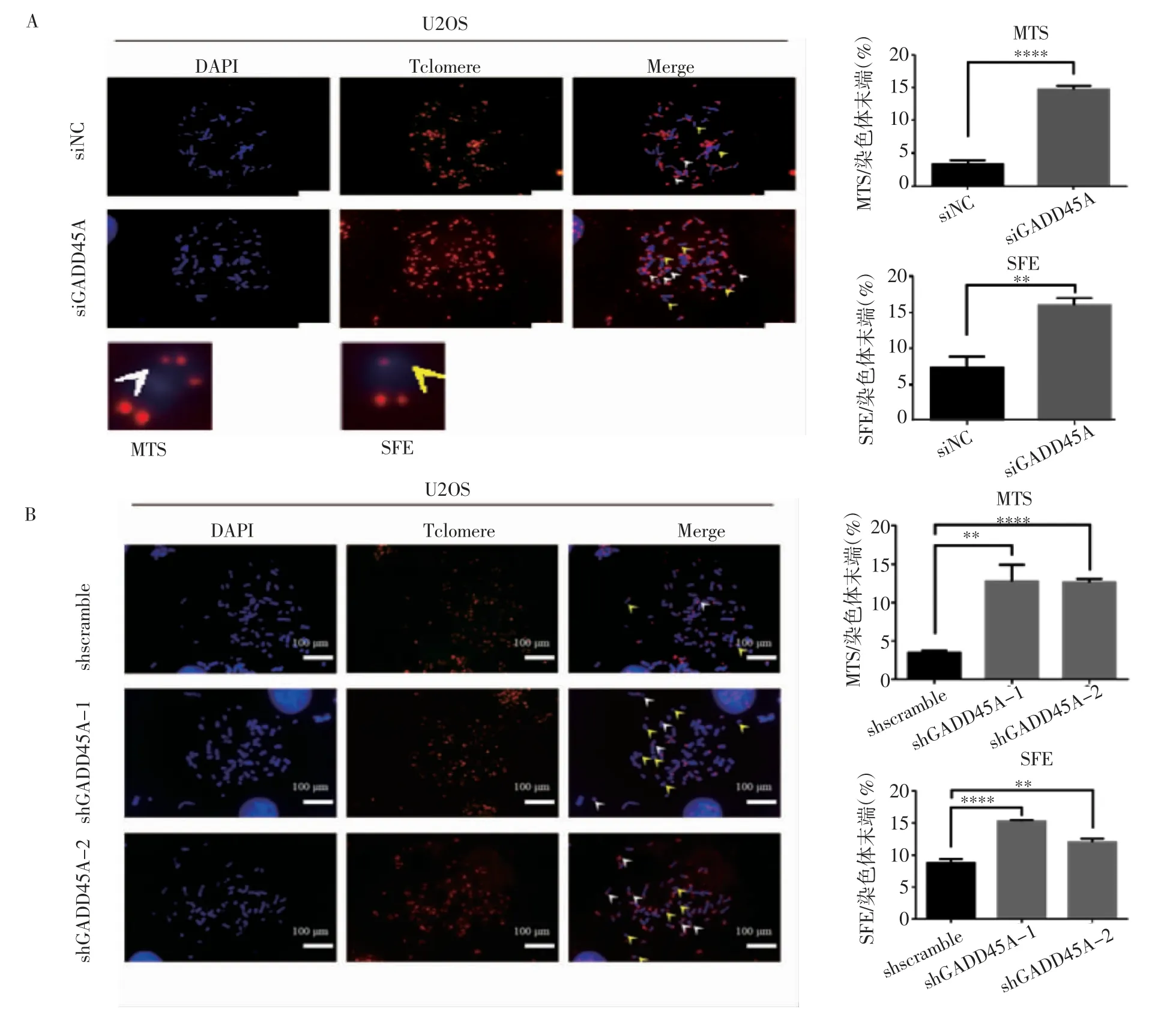
圖3 GADD45A 對骨肉瘤細胞端粒功能的影響Fig 3 The effect of GADD45A on the telomere function of osteosarcoma cells
2.3 GADD45A 對骨肉瘤端粒損傷造成的影響 免疫熒光-熒光原位雜交實驗結果顯示,在使用siRNA敲低GADD45A 和shGADD45A-1、shGADD45A-2 兩細胞系中,DNA 損傷指標γ-H2AX 與端粒共定位均上調(t=19.16、14.71、24.03,均P<0.000 1)(圖4)。
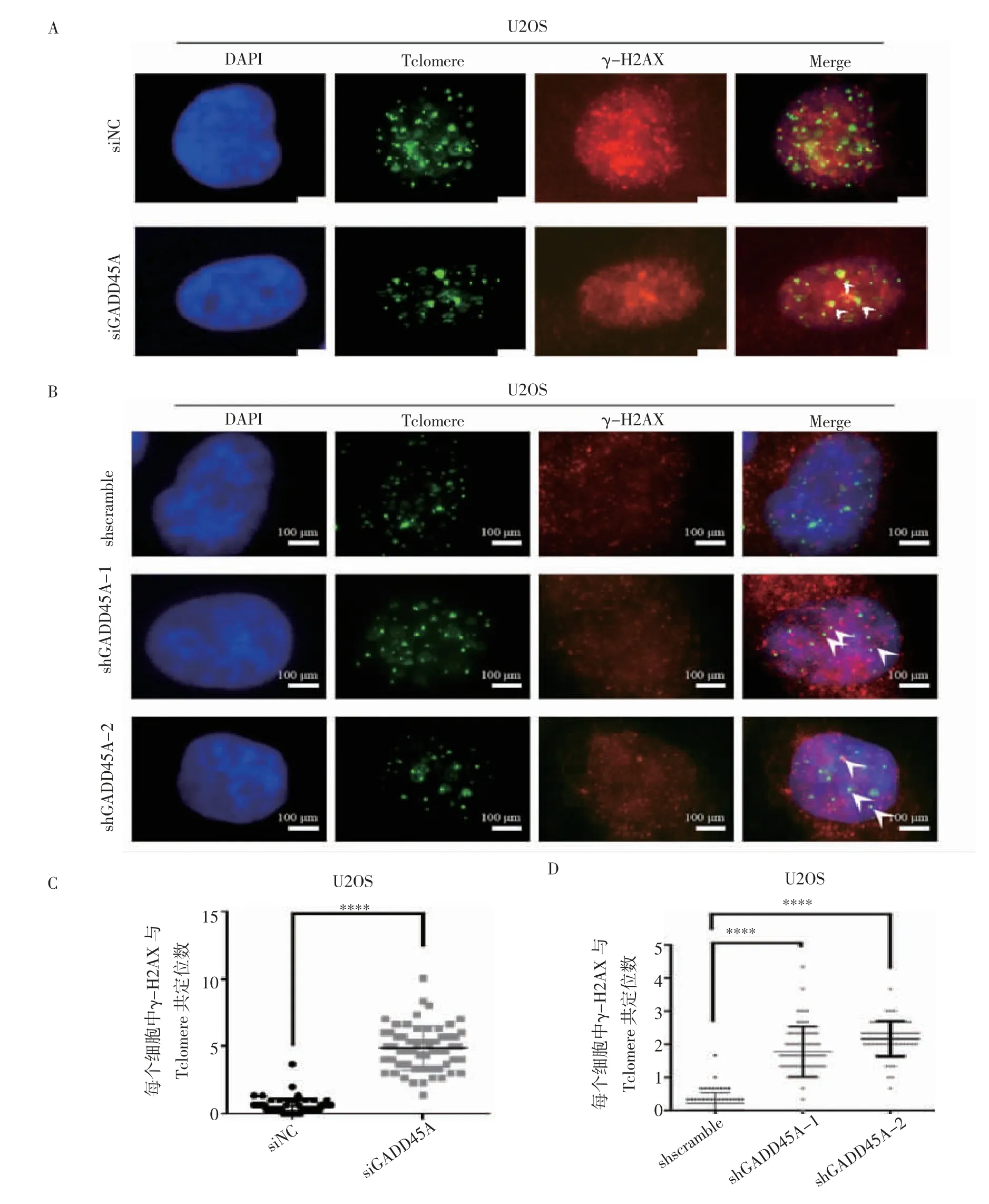
圖4 GADD45A 對骨肉瘤端粒損傷造成的影響Fig 4 The effect of GADD45A on the telomere damage of osteosarcoma cells
2.4 GADD45A 對骨肉瘤細胞ALT 活性的影響 C-circle 實驗結果表明,使用siRNA 敲低GADD45A 時,骨肉瘤細胞中C-circle 水平明顯上升(t=41.27,P<0.000 1)(圖5A)。采用shGADD45A-1 和shGADD45A-2 兩細胞系也顯示C-circle 水平升高(t=14.06、4.539,均P<0.05)(圖5B)。另一方面,通過免疫熒光-熒光原位雜交實驗,siRNA 和shRNA 的處理均導致PML 與端粒共定位顯著增多(t=10.65、16.69、18.10,均P<0.000 1),見圖5C~5D。
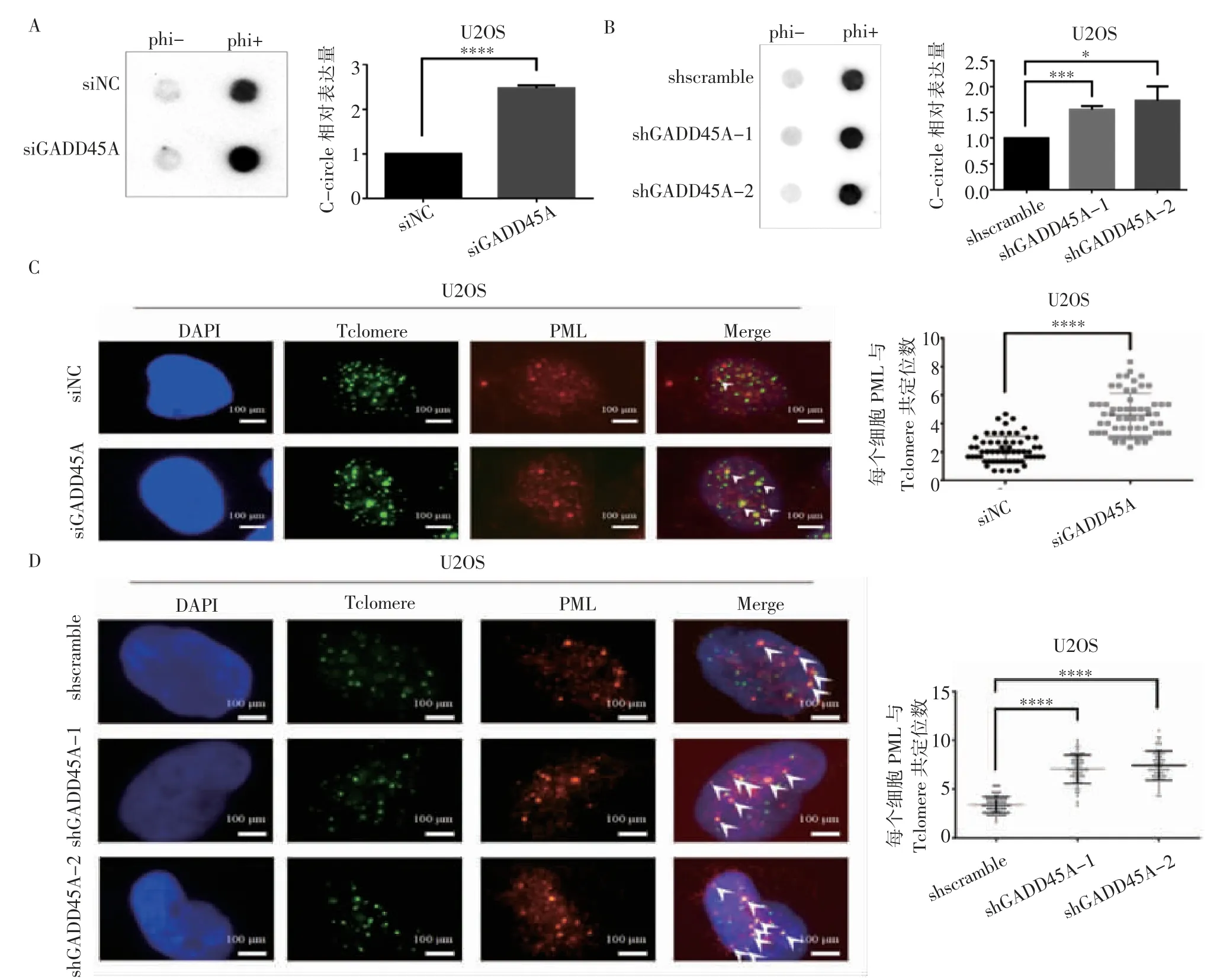
圖5 GADD45A 對骨肉瘤細胞ALT 活性的影響Fig 5 The effect of GADD45A on the ALT activity of osteosarcoma cells
3 討論
本研究發現敲低GADD45A 后抑制了骨肉瘤細胞的增殖,導致骨肉瘤細胞存活率下降。而端粒延長是腫瘤細胞維持增殖所必需的關鍵特征[9]。與端粒酶陽性腫瘤不同,ALT 陽性腫瘤端粒存在頻繁的DNA損傷。并且已有文獻報道GADD45A 參與了造血干細胞的DNA 損傷反應[10]。因此,本文通過H2AX 組蛋白變體(γ-H2AX)與端粒的定位來表征端粒損傷信號,首次發現敲低GADD45A 會引起骨肉瘤端粒損傷加劇。另一方面,本研究首次發現敲低GADD45A 會引起骨肉瘤細胞MTS 和SFE 顯著升高,表明GADD45A缺失會導致骨肉瘤端粒功能紊亂和復制性衰老并且引起端粒不穩定,加重脆性端粒的產生。
目前已有的研究表明,在間充質來源的腫瘤中,如中樞神經系統腫瘤和多種亞型的肉瘤,往往存在更高比例、更高水平的ALT 表型,這之中又以骨肉瘤最為突出[11]。APBs 是由早幼粒細胞白血病體(PML)、成簇聚集的端粒和一系列與DNA 復制、重組等功能相關的蛋白形成的復合體,是ALT 途徑延長端粒的場所。PML 是ALT 細胞形成APBs 并進一步維持端粒長度,造成端粒長度異質性的必要組分[12]。為了探究GADD45A 對ALT 的影響,筆者通過免疫熒光觀察PML 與端粒的共定位,發現在敲低GADD45A后,PML 與端粒的共定位明顯增多,說明GADD45A抑制了端粒到PML 的聚集,在ALT 途徑中發揮了負向調控作用。除此之外,C-circle 作為ALT 陽性腫瘤中的一種特異性且可定量的指標,可以更清晰地反映ALT 活性的變化。C-circle 是染色體外的CCCTAA 重復序列,來自于端粒的后隨鏈,與端粒TTAGGG 重復序列相對應[4]。本研究首次發現在敲低GADD45A 后,C-circle 水平顯著升高,與前面結果一致,說明GADD45A 抑制骨肉瘤細胞C-circle形成,限制了ALT 途徑發生。有文獻報道,ALT 途徑的過度激活會抑制ALT 細胞的生長[13-14]。本研究結果發現,敲低GADD45A 后,ALT 表型明顯增加,ALT 活性增加,細胞增殖受到抑制,有可能是因為GADD45A 的缺失導致ALT 被過度激活,這也會導致ALT 細胞的死亡。
目前研究認為,ALT 是一種以類似于同源重組的斷裂誘導復制(BIR)的方式進行端粒延長的途徑[15]。在端粒DNA 損傷斷裂后會激活RFC-PCNAPol δ 軸來促進APB 的組裝,開始后續RAD52 依賴和不依賴的兩種BIR 通路,整個過程還涉及BRCA1、RAD51 等多個同源重組相關蛋白[16-18]。本文已經證實GADD45A 確實參與了骨肉瘤的ALT 進程,結合以往文獻報道GADD45A 與PCNA 存在相互作用,提示其可能影響了DNA 損傷反應進而抑制骨肉瘤ALT 活性[19]。此外,已有研究表明GADD45A 可以與R-loop 結合介導CpG 島的局部去甲基化[20]。亞端粒區豐富的CG 含量提示這一過程在亞端粒位置同樣有可能實現[21]。GADD45A 介導的去甲基化過程可能抑制端粒區的異染色質化,使染色質收縮,從而阻礙ALT 相關蛋白與端粒的結合。這與過往研究中提到的ALT 端粒處松弛的染色質結構這一特點有所呼應[22]。另一方面,筆者推測GADD45A 與BRCA1的相互作用也有助于GADD45A 與R-loop 的結合,R-loop 進一步刺激同源重組過程也是引起ALT 相關表型變化的原因[19,23],后續筆者會對GADD45A 如何調控ALT 進行深入的探究。
綜上所述,GADD45A 缺失能夠抑制骨肉瘤增殖,并通過參與骨肉瘤ALT 徑影響端粒功能,造成端粒功能紊亂,加劇端粒DNA 損傷。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
