不同pH 和培養方式對海帶配子體生長發育的影響
2023-07-12 04:34:44陳書秀李曉捷李霞孫娟趙楠史良
水產學雜志 2023年3期
關鍵詞:生長
陳書秀,李曉捷,李霞,孫娟,趙楠,史良
(山東東方海洋科技股份有限公司,國家海藻與海參工程技術研究中心,山東省海藻與海參技術創新中心,山東省海藻遺傳育種與栽培技術重點實驗室,山東 煙臺 264003)
海帶是一種具有很高營養價值和經濟價值的大型褐藻,也是最早實現全人工養殖和育苗的藻類之一,在食品、化工、藥用等的市場需求量日益增加。海帶養殖也在防治赤潮,促進海區生態平衡方面起到重要的作用。因此,海帶養殖具良好的發展前景。苗種繁育是海帶大規模養殖的前提,目前采用的育苗方式主要有兩種:以海帶生活史為依據的傳統夏苗培育和以海帶配子體特性為依據的克隆育苗方法。與前者相比,克隆育苗方法工藝簡單,品種純度較高,且不受季節限制,是推動海帶育苗產業化的新模式。
配子體克隆育苗方法主要由配子體大規模培養、采苗和育苗管理3 部分構成,其中配子體人工高密度培養是該方法產業化的關鍵。目前已對配子體種質資源的保存[1-3]、培養條件[4-10]、擴培方法[11-14]等進行了大量研究,得出了海帶配子體適宜生長的溫度、光照及所需氮磷濃度范圍;可采取的擴培方法有打碎接種和通氣懸浮培養等。配子體培養不僅需要N、P、Fe 等營養的供應,無機碳也是配子體生長利用量最大的營養成分之一。岳國峰等[13,14]研究發現,海帶雌雄配子體只能利用游離的CO2作為碳源。海水中游離的CO2僅占無機碳總量的0.5%,當海帶配子體高密度培養時,無機碳必將成為最主要的限制因子。目前采用通空氣懸浮培養方法,主要作用是攪拌混勻和防止氧積累,對無機碳的補充作用不大。海水總無機碳濃度主要由溫度、堿度、鹽度和pH 決定,在其他條件一致的情況下,pH 可直接顯示無機碳的變化[15]。隨著無機碳濃度的改變,所引起的pH 的變化也會對海藻的生長發育產生一定的影響[16-19]。因此,在補充無機碳的過程中,必須保證pH 變化范圍與所培養海藻的適應范圍相一致。
海帶“奔牛”是榮成等養殖時間較長、較廣的海帶品種,是中國海洋大學劉濤教授及其團隊與山東尋山集團公司歷時5 年(2006—2011)聯合培育出的優良海帶品種,其與海帶親緣關系很近[20,21],與海帶的分化未達到種的水平,仍屬于海帶[22]。該研究嘗試在此海帶配子體通空氣高密度連續培養過程中,以CO2配氣裝置補充無機碳源,并通過控制培養液的pH 調整CO2的補充量及補充頻率,使海帶配子體在充足的碳源和適宜的pH 的范圍內生長,提高海帶配子體的生長率,以期為海帶配子體高密度培養提供一種更加高效的方式。
1 材料與方法
1.1 材料
取自實驗室保存的海帶‘奔牛’雌雄配子體在8℃~10℃、24 h 光照和光照強度30 μmol/(m2·s)下加富滅菌海水(10 mg/L NaNO3-N,1 mg/L KH2PO4-P)培養。
1.2 方法
1.2.1 配子體克隆的處理
取適量的雌雄配子體于容積為5 000 mL 的加入3 000 mL 滅菌海水的三角瓶中,利用小循環泵破碎處理10~15 min。粉碎后細胞液經300 目篩絹過濾,使雌雄配子體細胞段大小基本達到2~5 個細胞。
1.2.2 不同pH 對海帶配子體生長的影響
將處理過的雌雄配子體分別等體積、等密度加入于培養皿中,讓其自然附著于載玻片上。在海水中添加NaNO3和KH2PO4,總有效N 為10 mg/L,有效P 為1 mg/L 作為培養液。培養條件為:24 h 光照、光照強度40 μmol/(m2·s)、溫度為10℃。3 d 后更換不同初始PH(6.0、7.0)的培養液,以正常海水(pH8.2)培養液為對照組(組③),初始pH6.0 為組①、初始pH7.0 為組②。每6 h 測定pH 并更換培養液,經過30 d 培養之后,拍照(Olympus Ⅸ51),并采用Image-pro 統計數據。每個處理組設3 個培養皿,每個培養皿測定了3 個數據,即每個處理組共記錄9 個數據(n=9)。根據初始配子體平均長度(L0)和實驗結束時配子體平均總長度(L1)計算相對生長速率(RGR,%/d),即RGR=100×[ln(L1)-ln(L0)]/t,其中t 為試驗天數(d)。pH 采用1 mol/L HCl 和1 mol/L NaOH 調節,用pH 計測定。
1.2.3 不同培養方式對海帶配子體生長的影響
在18.9 L 礦泉水桶(有效水體15 L)中,以0.5 g/L 的密度接種處理過的海帶雌雄配子體。在海水中添加NaNO3和KH2PO4總有效N 為10 mg/L,有效P 為1 mg/L 作為培養液。在24 h 光照,光照強度40 μmol/(m2·s),溫度為10℃條件下培養。每5 d 更換一次培養液,15 d 時克隆液倒入篩絹過濾至不滴水時測量鮮重,計算特定生長率(SGR)。計算公式SGR=100%×[ln(W2)-ln(W1)]/(t2-t1),其中W1=起始重量,W2=結束重量,t1=起始時間,t2=結束時間。利用pH 計測定培養液48 h 的pH 變化。采用3 種培養模式,每種培養模式設置3 個重復。
具體方式如下:Ⅰ:靜置培養,每天搖瓶3 次,避免配子體克隆貼于瓶底;Ⅱ:通空氣懸浮培養,充氣機和培養容器之間安裝空氣過濾系統,充氣速率為400 mL/min,充氣頻率為每30 min 充氣5 min,每天共充氣24 次,以觀察到克隆上下翻轉為準。Ⅲ:定時定量補充CO2進行懸浮培養:在培養方式Ⅱ的基礎上,每2 d 每桶用純CO2壓力罐補充CO2(1~2 min),直到培養液pH7.0,充氣量為120 L/h。
1.2.4 不同配子體擴增培養方式對海帶配子體發育的影響
取1.2.3 實驗中3 種繼代培養方式培養60 d 的海帶雌雄配子體,分別用300 目篩絹過濾,獲得含2~5 個細胞的細胞段懸濁液,然后每組的雌雄配子體按照質量比2∶1 的比例混合均勻。取一定量的雌雄混合懸濁液加入到12 cm(直徑)的培養皿中,放置在溫度10℃,鹽度31,光照強度30 μmol/(m2·s),光照周期均為10L∶14D 條件下培養。在海水中添加NaNO3、KH2PO4、檸檬酸鐵作為培養液,培養液中總有效氮為10 mg/L,有效磷為1 mg/L,有效鐵為0.5 mg/L。每個處理組3 個重復。每個培養皿隨機觀察10 個100×視野,分別統計卵囊形成、排卵、幼孢子體和總細胞數,按卵囊形成數、排卵數和孢子體數之和占總細胞數的百分比計算發育率。
1.3 統計分析
2 結果與分析
2.1 不同pH 對海帶配子體生長的影響
間隔6 h 更換調整一次,處理組①、②、③的pH分別維持在6.0~6.1、7.0~7.1,及8.2~8.3。
由表1 可知,所用模型有統計學意義。pH(P<0.001)、性別(P<0.05)對海帶’配子體生長有顯著影響,但pH 與性別的交互作用對海帶配子體生長無顯著影響(P>0.05)。Eta2pH>Eta2性別,表明pH對海帶’奔牛’配子體克隆生長的影響大于性別的影響。pH 及性別對海帶配子體的生長有顯著影響。
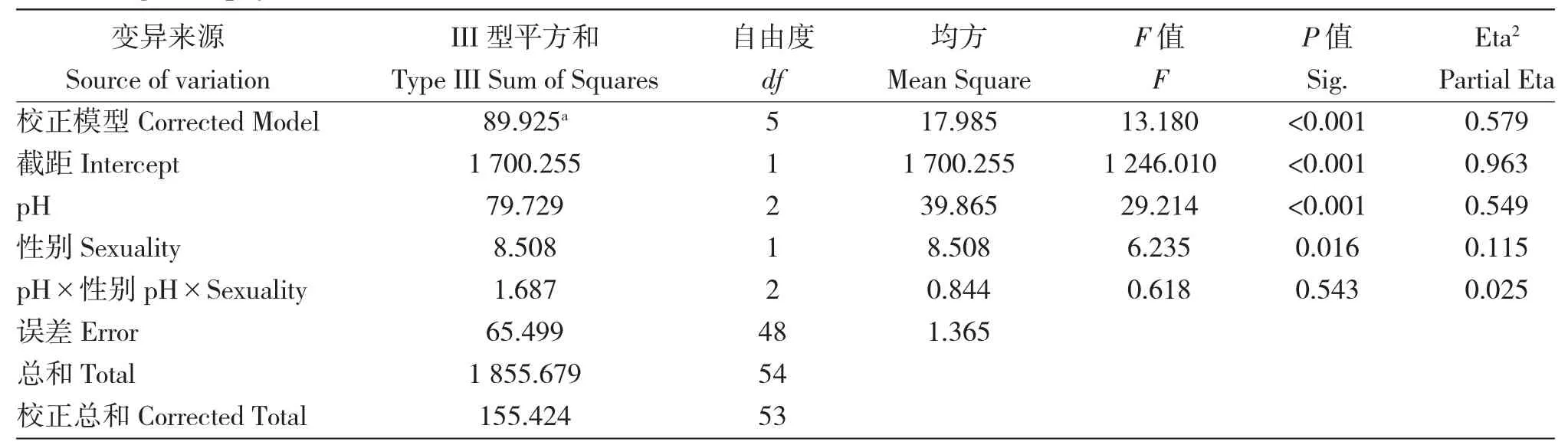
表1 不同pH、性別對海帶’奔牛’配子體生長影響的方差分析Tab.1 Variance analysis of effects of different pH values and sexuality on the growth of kelp S.japonica’Benniu’gametophytes
圖1 為雌雄配子體克隆的相對生長率在不同處理組的變化情況。結果顯示,處理組①(pH6.0~6.5),雌雄配子體相對生長率[♀(4.61±1.28)%,♂(3.35±1.23)%]顯著低于其他2 組,雌配子體的相對生長率(4.61±1.28)%顯著高于雄配子體(3.35±1.23)%。處理組②(pH7.0~7.3)雌雄配子體的相對生長率與對照組③無顯著差異(P>0.05),且在2 組培養條件下,雌、雄配子體的相對生長率無顯著差異(P>0.05)。
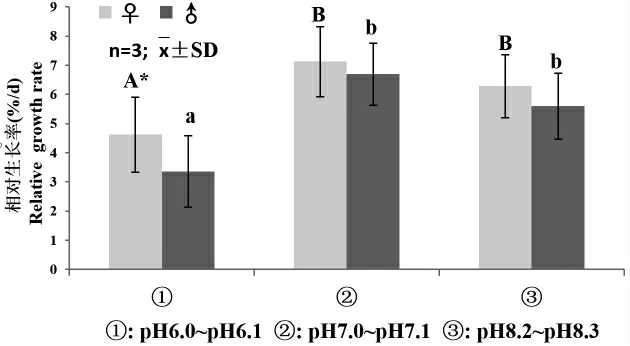
圖1 不同pH 下海帶’奔牛’雌雄配子體相對生長率Fig.1 Relative growth rate of female and male gametophytes of kelp S.japonica’Benniu’exposed to different pH values.Data are shown as means and SD(n=3)
2.2 不同培養方式及性別對海帶’奔牛’配子體生長的影響
48 h 內培養方式Ⅰ、Ⅱ、Ⅲ培養液的pH 變化范圍分別為8.2~8.9,8.2~8.8 和7.0~8.3(圖2)。
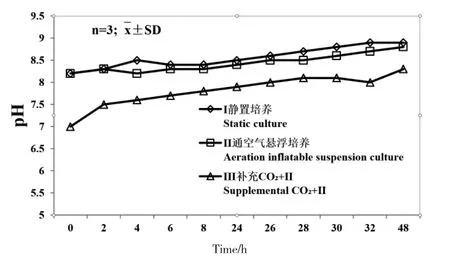
圖2 海帶配子體培養液48 h 內pH 變化Fig.2 The range of medium pH during 48 h
由表2 可知,所用模型有統計學意義,即培養方式(P<0.001)及性別(P<0.001)極顯著影響海帶’奔牛’配子體克隆的生長,但培養方式與性別的交互作用對海帶配子體克隆生長無顯著影響(P>0.05)。Eta2培養方式的影響>Eta2性別的影響,即培養方式對配子體克隆生長的影響大于性別。

表2 不同培養方式、性別對海帶’奔牛’配子體生長影響的方差分析結果Tab.2 Variance analysis of effects of different culture methods and sexuality on the growth of gametophytes in kelp S.Japonica‘Benniu’
不同實驗組的雌雄配子體克隆的特定生長率不同。靜置培養方式下,雌雄配子體特定生長率(♀10.03%,♂9.21%)顯著低于其他2 種培養方式。在3 種培養方式條件下,雌配子體(10.03%~15.41%)的特定生長率顯著高于雄配子體(9.21%~13.70%)。在補充CO2+通空氣懸浮培養條件下,雌雄配子體的特定生長率(♀15.41%;♂13.70%)顯著高于其他2種培養方式(圖3)。
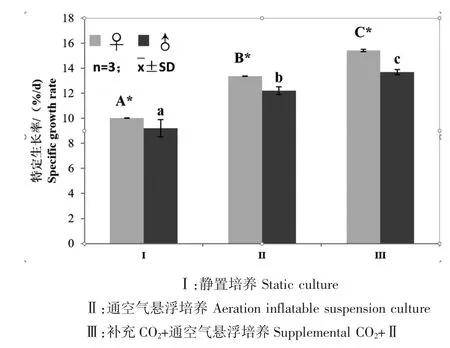
圖3 不同培養方式下海帶’奔牛’雌雄配子體特定生長率Fig.3 Specific growth rate of female and male gametophytes in kelp S.japonica‘Benniu’under the different cultivation methods.Data are shown as means and SD(n=3)
2.3 不同配子體擴增培養方式對海帶配子體發育的影響
在3 種配子體擴增培養方式下培養60 d 的配子體均可正常發育,且各組間的發育率無顯著差異(P>0.05,表3)。隨著發育時間的延長,各組的發育率顯著增加(發育時間P<0.001,表3)。3 種擴增培養方式培養的配子體均在第14 d 出現卵囊并排卵(表4),第27 d 的小苗率均在60%以上,最終發育率無顯著差異(P>0.05,表5)。
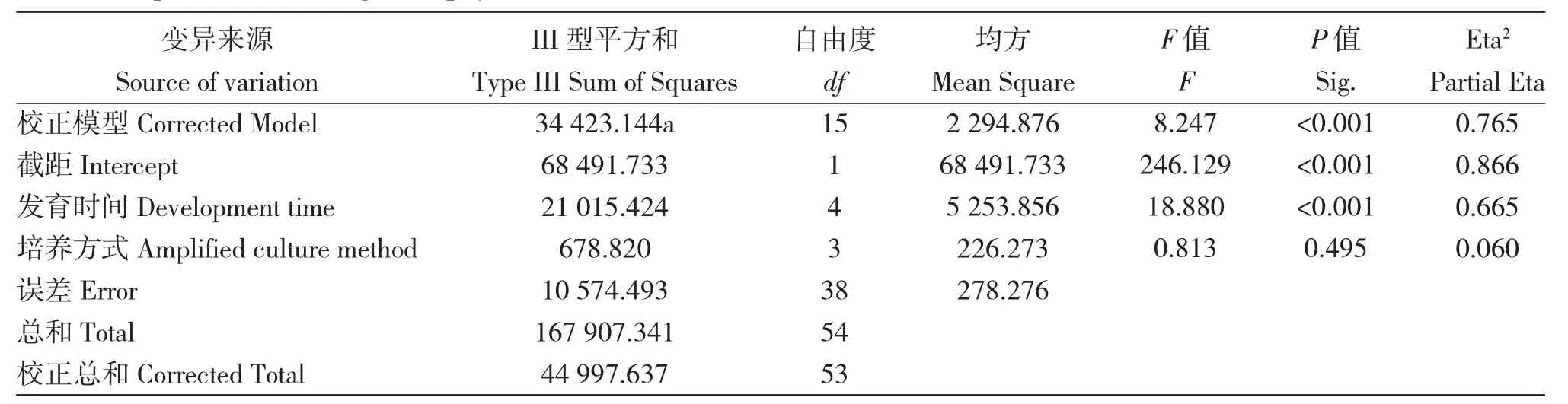
表3 不同配子體擴增培養方式及發育時間對海帶’奔牛’配子體發育影響的方差分析結果Tab.3 Variance analysis of effects of different culture methods and development periods on the development of kelp S.Japonica‘Benniu’gametophytes
表4 不同擴增培養方式對海帶’奔牛’配子體發育的影響Tab.4 Effects of different culture methods on gametophyte development rate of kelp S.japonica ’Benniu’n=3;D;%

表4 不同擴增培養方式對海帶’奔牛’配子體發育的影響Tab.4 Effects of different culture methods on gametophyte development rate of kelp S.japonica ’Benniu’n=3;D;%
表5 不同擴增培養方式對海帶’奔牛’配子體發育率的影響Tab.5 Effects of different culture methods on gametophyte development rate of kelp S.japonica ’Benniu’.Data are shown as means and SD(n=3)n=3;(D;%

表5 不同擴增培養方式對海帶’奔牛’配子體發育率的影響Tab.5 Effects of different culture methods on gametophyte development rate of kelp S.japonica ’Benniu’.Data are shown as means and SD(n=3)n=3;(D;%
注:同一列上的相同大寫字母表示發育率在相同培養時間不同培養方式下差異不顯著(P>0.05);同一行上的不同小寫字母表示發育率在相同培養方式不同培養時間差異顯著(P<0.05)。Note:Means with the same capital letters in the same column are not significant differences between culture methods(P>0.05);means with the different letters in the same line are significant differences between sampling times in the same culture method(P<0.05).
3 討論
3.1 不同pH 和培養方式對海帶配子體生長的影響
海水中的無機碳是海洋藻類生長利用量最大的營養成分之一,是進行光合作用的必需碳源,但不同藻類利用無機碳的形式不同,不同藻類即使在相同生長條件下所利用的無機碳濃度也不盡相同[13-15,23-25]。岳國峰等[13,14]比較了海帶配子體和幼孢子體無機碳素營養的利用,發現海帶雌雄配子體只能利用游離的CO2為碳源,海水中CO2濃度的升高有效加快了雌雄配子體利用無機碳的速度;還發現海水pH 升高會抑制海帶配子體對無機碳的利用。pH8.7 時配子體對無機碳的吸收利用率降低40%。本實驗結果表明,pH(P<0.001)顯著影響海帶’奔牛’配子體生長,pH7.0~7.5 組配子體相對生長率最高。靜置培養與傳統通空氣懸浮培養條件下的特定生長率也顯著低于補充CO2培養組,前者的pH 在48 h 達到8.7~8.9,這可能是配子體生長受限的重要因素之一。海水總無機碳濃度主要由溫度、堿度、鹽度和pH 決定,在其他條件一致的情況下,pH 可直接顯示無機碳的變化。Skirrow 等[15]研究表明,天然海水中(pH8.1~8.3),CO2濃度隨著pH 的升高而逐漸降低,當pH 接近于9 時,游離CO2濃度接近于0。因此,靜置培養與傳統通空氣懸浮培養條件下的pH 達到8.7~8.9,表明培養液中CO2已成為限制因素,若不及時更換培養液,配子體生長將會受到抑制。
該實驗嘗試在傳統通氣懸浮培養的基礎上定時定量補償CO2,將培養液pH 持續穩定在7.0~8.4左右,并保持CO2的含量一直處于較高的水平,由此保證碳源充足,減少pH 升高引起的負面效應。結果表明,該方法培養的配子體特定生長率顯著高于其他2 組。此結論與張栩等[26]結論相同,但與之相比,本文所采用的補充CO2的方法簡單易操作,節約成本,更有利于在育苗基地推廣使用。
3.2 不同pH 和培養方式對海帶配子體發育的影響
CO2進入海水后,會發生一系列化學反應,改變海水酸化和酸堿度平衡等,不可避免地影響海水中的海藻等初級生產者。高濃度CO2對海藻的生長速率、光合作用、呼吸作用、生化組分、營養鹽的吸收等都有顯著影響,且存在明顯的種間差異[27-32]。這些差異性不僅與藻體無機碳利用機制有關,也有可能是CO2加富使水體中pH 下降引起。如pH 降低會對某些褐藻的生長和光合作用造成較大的負面效應[33],也會影響海帶如巨藻(Macrocystis pyrifera)等早期生命史階段游孢子萌發、配子體性別比例、配子體發育等[16-19]。Roleda 等[17]研究表明,pH 降低至7.59~7.60,巨藻游孢子的萌發率顯著降低,配子體生長也受到一定的抑制,但同時加入可溶解無機碳,有效減少了pH 降低引起的這種負面效應。Leal等[19]研究發現,pH 降低提高了巨藻和裙帶菜(Undaria pinnatifida)游孢子的萌發率,促進了配子體的生長,對配子體性別比例及其后期發育無顯著影響。該研究表明,適量增加海水培養液中CO2對海帶配子體生長有明顯的促進作用;當配子體長期處在pH<7 的環境中,生長顯著受抑制。
為了檢驗配子體長期在較高濃度CO2的海水培養液中生長,是否會對其發育能力產生不利的影響。以在3 種不同方式下培養60 d 的海帶’奔牛’雌雄配子體以質量比2∶1 混合后,在適宜條件下進行了發育培養,實驗結果顯示,3 種培養方式下培養的配子體均可正常發育,發育進程及發育率無顯著差異。由此可以看出,在傳統通空氣擴增培養的基礎上,根據培養液pH 變化,定時定量補充CO2可以用于海帶配子體大規模高密度培養,與光生物反應器相比,成本低廉,操作簡便,適合在育苗基地大規模推廣。
3.3 結論
比較發現:配子體生長的最適生長pH 范圍為7.0~8.3。其次從配子體培養成本出發,通空氣懸浮培養過程中每2 d 補充一次CO2,利用純CO2壓力罐(出氣量:120 L/h)向15 L 海水中充氣1~2 min,保證了充足的碳源,使培養液pH 范圍控制在7.0~8.4,排除了pH 對配子體生長不利影響,避免了頻繁測定pH 對配子體的混種或雜藻等污染。發育實驗檢驗了該方法使配子體長期處在較高濃度CO2的海水培養液中生長,并未對其發育能力產生不利的影響。該方法可用于海帶配子體大規模高密度培養成本低廉,操作簡便,適合在育苗基地大規模推廣。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
