
瀕危植物秀麗兜蘭葉綠體基因組特征與系統發育分析
2023-07-20 05:11:38范繼征李秀玲李明智卜朝陽何荊洲曾艷華
熱帶作物學報 2023年6期
范繼征 李秀玲 李明智 卜朝陽 何荊洲 曾艷華

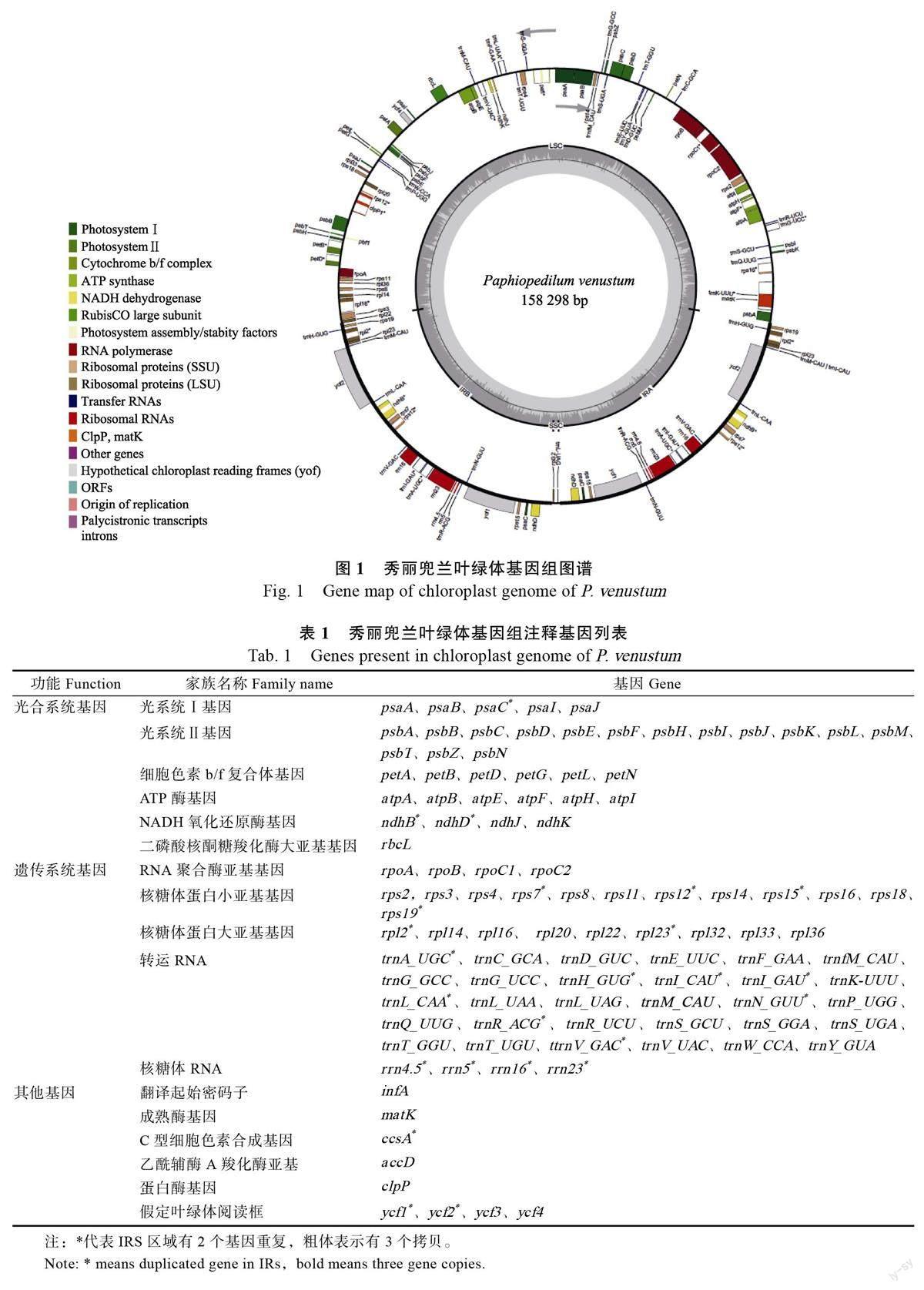
關鍵詞:秀麗兜蘭;葉綠體基因組;特征分析;系統發育分析
中圖分類號:S 682.31 文獻標識碼:A
秀麗兜蘭[Paphiopedilum venustum (Wall. exSims) Pfitzer]隸屬于蘭科杓蘭亞科兜蘭屬,是第一個被人工栽培的兜蘭屬植物[1],主要分布于我國西藏東南部至南部,以及孟加拉國、不丹、印度東北部、尼泊爾等國家和地區,野外花期1—3月[2]。秀麗兜蘭植株直立、叢生,株高7.0~9.0 cm,葉片數2~3 片,長7.5~12.0 cm,寬2.5~3.5 cm,呈狹矩圓狀橢圓形,表面有網格斑,背面有密集的紫色斑點, 花葶長9.2~15.4 cm, 花朵橫徑5.4~8.6 cm,縱徑2.9~3.4 cm,花朵色澤艷麗,花期長,花和葉均極具觀賞價值。但由于人類活動以及自身生長要求和生長特點等原因,秀麗兜蘭已成為瀕危物種(EN),受到國際社會的嚴格保護[3-4]。
葉綠體是植物光合作用發生的細胞器,具有一套完整的基因組[5],與核基因組等其他基因組相比,葉綠體基因組具有高度的保守性以及自我復制機制、進化相對獨立、基因組小、結構穩定、突變率低等特點[6-8]。隨著二代測序技術的飛速發展,通過測序組裝和注釋獲得的完整的葉綠體基因組序列成本降低、速度加快[9],葉綠體基因組也越來越多的為進化分析、系統發育及分類鑒定等提供信息[10]。目前有關葉綠體基因組特征分析已被廣泛應用于多種兜蘭屬植物,有對單獨一個種進行葉綠體基因組分析,例如白旗兜蘭(P. spicerianum)[11]、白花兜蘭(P. emersonii)[12]、格麗兜蘭(P. gratrixianum)[13]、麻栗坡兜蘭(P. malipoense)[14]、紫紋兜蘭(P. purpuratum)[15]、飄帶兜蘭(P.parishii)[16]、陳蓮兜蘭(P. tranlimianum)[17]等,也有對多個種進行比較分析[18-19]。現有研究結果表明兜蘭屬植物葉綠體基因組大小為154689~162 590 bp,其結構與大多數陸生植物葉綠體結構相似,為典型的四分結構,包括大單拷貝區(large single copy, LSC)、小單拷貝區(small singlecopy, SSC)和2 個反向重復區(inverted repeat,IR),各區GC 含量差異較大,其中IR 的GC 含量較高,不同的蛋白質編碼在同義密碼子的使用上有所不同,偏好A 或U(T)結尾的密碼子。此外,系統發育分析打破了形態學和細胞學等的局限性,能夠更好地反映物種之間的親緣關系[11-19]。
有關秀麗兜蘭的研究多集中在非共生萌發和繁殖技術[20]、細胞學研究[21-23]、病害鑒定[24]、分布地區和生長環境[25]等,尚無秀麗兜蘭葉綠體基因組的報道。本研究通過高通量測序方法獲得秀麗兜蘭完整的葉綠體基因組信息,對其進行注釋,并對基因組中序列重復、SSR 位點及蛋白編碼基因的密碼子使用偏好性進行分析,同時分析了系統發育關系。本研究結果可為秀麗兜蘭及其他兜蘭屬植物種質資源鑒定、系統發育、遺傳育種、種群恢復以及生物多樣性保護等提供理論依據。
1 材料與方法
1.1 材料
供試材料為遷地保存在廣西農業科學院花卉研究所蘭科植物種質資源圃(22°48′N,108°22′E)的秀麗兜蘭,取長勢良好、無病蟲害的新鮮幼嫩葉片擦拭干凈,用液氮速凍后于?80 ℃保存備用。
1.2 方法
1.2.1 DNA 提取、文庫構建及測序 取葉片100 mg,采用改良CTAB 法[26]提取總DNA,樣品基因組DNA 質量檢測合格后,用超聲波將合格的DNA 片段化,然后對片段化的DNA 進行片段純化、末端修復、3′端加A、連接測序接頭,再用瓊脂糖凝膠電泳進行片段大小選擇,進行PCR擴增形成測序文庫,并進行文庫質檢,質檢合格的文庫用BGISEQ-500 平臺進行測序(廣州佰德生物科技有限公司)。
1.2.2 葉綠體基因組的組裝與注釋 對測序得到的原始數據(raw data)進行數據過濾,去除其中的接頭序列及低質量reads 獲得高質量的cleandata。使用Bowtie2 v.2.3.4.3 軟件[27]比對近緣物種葉綠體基因組數據庫,將比對上的reads 數量最多的3 個CDS 序列作為項目樣品的葉綠體基因組起始組裝序列,采用NOVOPlasty v4.2.1 軟件[28]組裝葉綠體基因組。利用GeSeq 和blast 軟件對葉綠體基因組CDS 序列進行注釋,并利用GeSeq 和hmmer 軟件對葉綠體基因組rRNA序列進行注釋,使用tRNAscan-SE 和ARAGORN軟件對葉綠體基因組tRNA 序列進行注釋,并進行手動調整和確認獲得最終注釋結果,最后采用OrganellarGenomeDRAW軟件制作葉綠體基因組圖譜[29-35]。將組裝注釋好的秀麗兜蘭基因組序列上傳至Gen-Bank 數據庫,獲得登錄號:MZ150831。
1.2.3 葉綠體基因組SSR 分析 利用MISAv1.01(MIcroSAtellite identification tool)軟件對秀麗兜蘭進行cpSSR 的分析[36]。參數設置為單核苷酸(mononucleotide)、二核苷酸(dinucleotide)、三核苷酸( trinucleotide)、四核苷酸(tetranucleotide)、五核苷酸(pentanucleotide)和六核苷酸(hexanucleotide)的重復數閾值分別為10、6、5、5、5 和5。
1.2.4 密碼子使用分析 計算秀麗兜蘭cpDNA 密碼子的使用情況和相對密碼子的偏好性(relativesynonymous codon usage, RSCU),RSCU 值則以1為限,若RSCU>1,該密碼子使用頻率較高;若RSCU<1,該密碼子使用頻率較低。
1.2.5 系統發育關系分析 從NCBI 數據庫中下載16 個兜蘭屬和2 個杓蘭屬植物,利用mafft 7.0軟件進行cpDNA 序列比對,使用fasttree2.1.10軟件建立系統發育樹,選擇GTR+Gamma 模型,Shimodaira-Hasegawa 檢測[37-38]。
2 結果與分析
2.1 秀麗兜蘭葉綠體基因組的基本特征
秀麗兜蘭葉綠體基因組具有典型的環狀四分體結構,包括1 個大單拷貝區(large single-copyregion, LSC)、1 個小單拷貝區(small single-copyregion, SSC)和2 個反向重復區(IR),總長度為158 298 bp,其中LSC、SSC 和2 個IR 長度分別為87 775、949、34 787 bp,LSC、SSC 和IR 區域的GC 含量分別為32.6%、25.1%和39.1%。秀麗兜蘭葉綠體基因組共含129 個基因,包括79 個蛋白編碼基因,38 個tRNA 基因、8 個rRNA 基因和4 個假基因(2 個ndhD、1 個ndhJ、1 個ndhK),其中22 個完整的基因位于IR 重復區,包括10個蛋白編碼基因,8 個tRNA 和4 個rRNA,1 個基因rpl22 位于LSC-IRB 連接處。IR 重復區域還含有1 個假基因ndhD 以及rps12 基因的第2 和第3 個外顯子,nps12 的第1 個外顯子位于LSC 區域(圖1)。秀麗兜蘭葉綠體基因組注釋基因信息見表1。
2.2 秀麗兜蘭葉綠體基因組簡單序列重復
秀麗兜蘭葉綠體基因組簡單序列重復(SSR)信息見表2,從中共鑒定出78 個SSR 位點,未發現五核苷酸重復,最豐富的是單核苷酸重復,有66 個,占總SSRs 的84.62%,其次是二核苷酸重復8 個,占總SSRs 的10.26%,三核苷酸重復和四核苷酸重復均為1 個,六核苷酸重復2 個。其中63 個單堿基重復都是由A 或T 構成,因此,SSRs 的堿基組成偏向使用A/T 堿基。
2.3 秀麗兜蘭葉綠體基因組密碼子偏好性
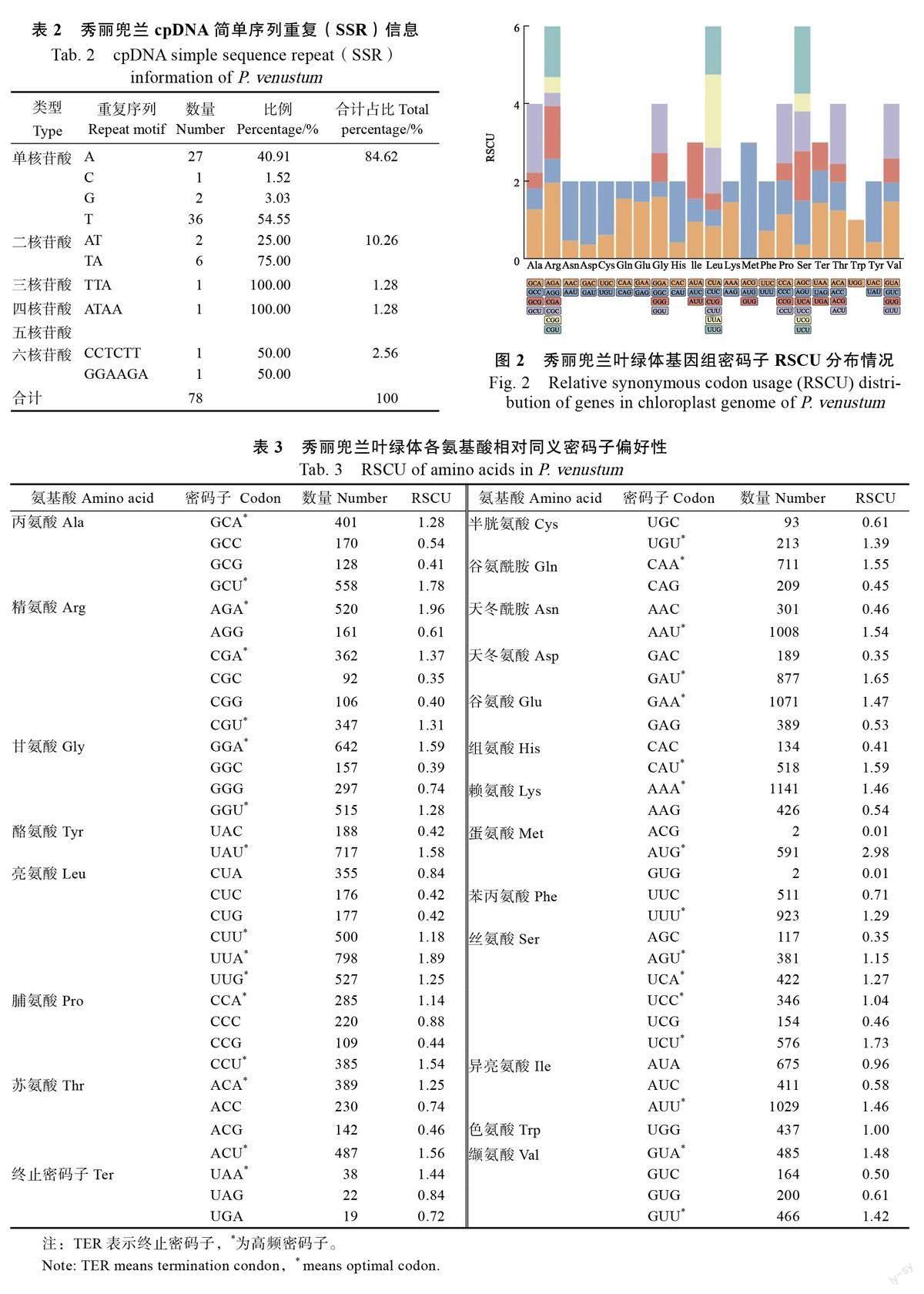
秀麗兜蘭葉綠體基因組中各氨基酸的相對同義密碼子使用頻率(relative synonymous codeusage, RSCU)分析結果見圖2 和表3。從中可以看出,高頻密碼子(RSCU>1)共有32 個,其中,16 個以U 結尾,13 個以A 結尾,以G 結尾的有2 個,以C 結尾的密碼子有1 個,說明秀麗兜蘭葉綠體基因密碼子偏好性以U 和A 結尾,與較低的基因組和蛋白編碼基因GC 含量一致。
2.4 系統進化分析
基于葉綠體基因組序列對18 個蘭科植物(含兜蘭屬物種16 個、近緣屬杓蘭屬物種2 個)進行系統發育分析,并構建系統發育樹(圖3)。從圖中可以看出,18 個物種分為3 大類,其中,紫花杓蘭和西藏杓蘭與其他物種遺傳距離較遠,被單獨聚成一類;麻栗坡兜蘭、硬葉兜蘭等5 個物種均歸為一類,即寬瓣亞屬;秀麗兜蘭和帶葉兜蘭、紫紋兜蘭等11 個物種歸為一類,屬于兜蘭亞屬,且秀麗兜蘭與紫紋兜蘭關系最近。
3 討論
兜蘭是蘭科(Orchidaceae)兜蘭屬(PaphiopedilumPfitzer)植物的統稱,因具有拖鞋狀的唇瓣又被稱為“拖鞋蘭”或“仙履蘭”,具有極高的觀賞價值和商業價值[1, 39-40]。本研究采用第二代高通量測序技術對秀麗兜蘭葉綠體基因組進行測序、組裝和注釋,獲得了秀麗兜蘭完整的葉綠體基因組。研究發現秀麗兜蘭葉綠體基因組與前人[11-19]研究的兜蘭屬其他植物相似,具有典型的四分體環狀結構,GC 含量遠小于AT 含量,其中IR 區域GC 含量最高,這與rRNA 的GC 含量較高,且全部定位在這些區域密切相關[41]。
簡單序列重復(SSR)多存在于真核生物的細胞器基因組中,具有豐富的多態性,高度的重復性以及良好的可靠性和共顯性,常被用于對種質資源進行遺傳結構分析、物種鑒定和群體遺傳學研究[19, 42]。本研究從秀麗兜蘭葉綠體基因組中鑒定出78 個SSR 位點,單核苷酸有66 個,占總SSRs 的94.62%,表明單核苷酸重復可能提供了更多的系統發育信息,63 個單堿基重復和其他堿基重復都是由A 或T 構成,A/T 重復類型占據主要地位,這與前人[18-19]對其他幾種兜蘭屬植物研究結果一致,得到的這些SSR 序列可為兜蘭屬植物篩選SSR 標記和指紋圖譜研究提供依據。
在植物進化過程中,密碼子的使用普遍表現出偏好性,自然選擇、物種突變和遺傳漂變是影響密碼子偏好性的主要原因,密碼子偏好性用相對同義密碼子使用頻率(RSCU)表示,若密碼子無偏好性,則RSCU 值為1,如果該密碼子比其他同義密碼子來說出現更頻繁,RSCU 值大于1,反之亦然[43-45]。本研究秀麗兜蘭葉綠體基因組RSCU>1 的密碼子共有32 個,其中以A 和U 結尾的有29 個,超過90.6%,而RSCU<1 的密碼子則多以C 或G 結尾,表明秀麗兜蘭葉綠體基因組偏向于使用以A 或U 結尾的同義密碼子,與丁銳等[46]對杓蘭屬的研究結果一致,這可能與葉綠體基因組較低的GC 含量有關,進一步表明密碼子偏好性可能受基因組GC 含量的影響[47]。根據秀麗兜蘭葉綠體基因組的密碼子偏好性可對目的基因進行優化,進一步提高基因轉化和表達效率,這一研究結果可為秀麗兜蘭遺傳育種及葉綠體基因工程的外源基因高效、穩定表達提供科學依據。
從18 種蘭科植物葉綠體基因組系統發育分析發現,兜蘭屬和杓蘭屬是明顯分開的兩大類,兜蘭屬又分兩大類,一類包含白花兜蘭、硬葉兜蘭、麻栗坡兜蘭、德氏兜蘭和杏黃兜蘭;另一類包括小葉兜蘭、陳蓮兜蘭、格麗兜蘭、白旗兜蘭、紫紋兜蘭、秀麗兜蘭、帶葉兜蘭、長瓣兜蘭、飄帶兜蘭、同色兜蘭和雪白兜蘭,其中秀麗兜蘭與紫紋兜蘭親緣關系密切。這一分類與其他學者利用形態學、細胞學等方法得到的分類結果相似[48-49],與SUN 等[18]和胡國家[19]通過葉綠體基因組序列建立的系統發育樹基本一致,顯示出葉綠體基因組序列在物種分類和進化研究中具有更高的分辨率和可靠性[50],證明了植物葉綠體基因組系統發育樹的構建在物種鑒定方面的準確性,同時還需要更加全面密集的取樣,才能更準確地反映發育關系。
4 結論
秀麗兜蘭葉綠體基因組為典型的四分結構,總基因組大小為158 298 bp,總GC 含量為35.4%,含129 個基因,共鑒定出78 個SSR 位點,堿基組成偏向使用A/T 堿基,密碼子偏好性以U 和A結尾,系統發育分析顯示秀麗兜蘭與紫紋兜蘭為姐妹種關系。研究結果可為兜蘭屬植物物種鑒定、生物進化、系統發育、資源保護及分子生物學等研究提供理論參考。
