不同生境黑老虎根際和內生真菌多樣性分析
2023-08-20 05:13:34劉濤馬楠金吉芬彭志軍李興忠汪得凱
廣西植物 2023年7期
劉濤 馬楠 金吉芬 彭志軍 李興忠 汪得凱


摘 要:? 為探究不同生長條件下黑老虎根際和根部內生真菌群落組成和多樣性及其與土壤環(huán)境因子的相關性,該文應用Illumina高通量測序方法對貴州3個不同生境下黑老虎根際和根部內生真菌進行了研究。結果表明:(1)3種生境下,根際土壤真菌OTU數(shù)量(3 867)遠多于根部內生真菌(801),其中根際土壤真菌共有的OTU為72個,共注釋到5個門、49個屬,大多為子囊菌門;屬水平上被孢霉屬、外瓶柄霉屬、 柱孢屬占比較高;根部內生真菌共有的OTU為14個,共注釋到2個門、11個屬,子囊菌門(13個,占比92.9%)占絕對優(yōu)勢,屬水平上被孢霉屬、外瓶柄霉屬、 柱孢屬和叢赤殼屬占比最高; 所有樣本中,共有的OTU僅為6個,注釋到2個門、5個屬,子囊菌門(5個,占比83.3%)為優(yōu)勢門;在屬水平上,占比最高的為外瓶柄霉屬(2個,33.3%),其余分別為被孢霉屬、柱孢屬和叢赤殼屬 。Alpha多樣性分析表明,根際土壤的真菌群落多樣性和豐富度均顯著高于根部內生真菌,而野生生境的真菌多樣性高于栽培生境。(2)在門水平上,3個生境下主要內生真菌類群均為子囊菌門(Ascomycota)和擔子菌門(Basidiomycota),占總菌群的88.28%;在屬水平上,不同生境條件下,根際和根部內生真菌群落結構差異明顯;栽培生境下,根部內生真菌菌群具有一定的偏好性,而野生生境下,根部內生真菌菌群均勻度更高;FUNGuild真菌群落功能預測顯示,栽培生境下的病理-腐生營養(yǎng)型(pathotroph-saprotroph) 在根部內生真菌中占比較高,而野生生境下的腐生營養(yǎng)型(saprotroph)及共生營養(yǎng)型(symbiotroph)占比較高。(3)土壤環(huán)境因子對根部內生真菌和根際真菌的影響方式不同,其中土壤總鉀(TK)和土壤總磷(TP)與黑老虎根部內生真菌香農指數(shù)和辛普森指數(shù)顯著正相關,而土壤有機質(SOM)、總氮(TN)和速效氮(AN)與黑老虎根際土壤真菌Ace指數(shù)和Chao1指數(shù)顯著正相關。綜上表明,SOM、TN、AN是影響黑老虎根際土壤真菌群落的主要土壤環(huán)境因子。
關鍵詞: 黑老虎, ITS, Illumina高通量測序, 真菌群落, 土壤環(huán)境因子, 多樣性
中圖分類號:? Q939
文獻標識碼:? A
文章編號:? 1000-3142(2023)07-1222-14
收稿日期:? 2022-12-16
基金項目:? 浙江省重點研發(fā)項目(2021C02043); 浙江理工大學科研啟動基金(19042142-Y)。
第一作者: 劉濤(1982-),碩士,副研究員,研究方向為果樹栽培育種,(E-mail)737689429@qq.com。
通信作者:? 汪得凱,博士,研究員,研究方向為藥用植物資源,(E-mail)kay77@163.com。
Diversity of rhizosphere and root endophytic fungi in
Kadsura coccinea under different habitats
LIU Tao1, MA Nan2, JIN Jifen3, PENG Zhijun1, LI Xinzhong1, WANG Dekai2*
( 1. Citrus Fruit Research Institute in Guizhou Province, Guiyang 550100, China; 2. College of Life Sciences and Medicine, Zhejiang Sci-Tech University,
Hangzhou 310018, China; 3. Institute of Crop Germplasm Resources, Guizhou Academy of Agricultural Sciences, Guiyang 550100, China )
Abstract:? Kadsura coccinea is a perennial vine medicinal plant,? and its growth, development, and authentic medicinal qualities are affected by growing habitat, rhizosphere soil, and endophytic fungi. In order to explore the composition and diversity of fungal community in K. coccinea under different habitats, as well as their correlations with soil environmental factors, the rhizosphere and root endophytic fungi of K. coccinea from three different habitats in Guizhou were studied based on Illumina high-throughput sequencing method. The results were as follows: (1) Under three habitats, the number of OTU detected in rhizosphere soil (3 867) was much more than that in root endophyte (801). The common OTU of rhizosphere soil was 72, which were annotated to 5 phyla and 49 genera, most of which were ascomycetes. At the genus level, the proportions of Mortierella, Exophiala, and Cylindrocarpon were relatively high. The common OTU of root endophytic fungi was 14, which were annotated to 2 phylums and 11 genera. Ascomycetes (13, accounting for 92.9%) were overwhelmingly dominant. At the genus level, Mortierella, Exophiala, Cylindrocarpon and Nectria were dominant genra. There were only 6 OTU in total, annotated to 2 phylums and 5 genera, and the ascomycetes phylum (5, accounting for 83.3%) were the dominant phylum. At the genus level, Exophiala (2, 33.3%) accounted for the highest proportion? and the rest were Mortierella, Cylindrocarpon and Nectria, respectively. Alpha diversity analysis showed that the diversity and richness of fungal communities in rhizosphere soil were significantly higher than those in root endophytic fungi, while the diversity of fungi under wild habits was higher than that under cultivated habits. (2) At the phylum level, the main endophytic fungal groups under three habitats were Ascomycota and Basidiomycota, accounting for 88.28% of the total flora. At the genus level, the community structures of endophytic fungi in rhizosphere and root were significantly different under different habitats. The endophytic fungi flora in the root had a certain preference under the cultivation habitat, while the evenness under the wild habitat? was higher than that under cultivation habitat. The prediction functions of fungal community based on FUNGuild showed that pathotroph-saprotroph type in the root endophytic fungi was relatively high under the cultural habit, while the proportion of saprophytic type and symbiotroph type was relatively high under the wild habit. (3) Soil environmental factors had different effects on root endophytic fungi and rhizosphere fungi in K. coccinea. Total potassium (TK) and total phosphorus (TP) in soil were positively correlated with Shannon index and Simpson index of endophytic fungi in K. coccinea? root; whereas, soil organic matter (SOM), total nitrogen (TN) and available nitrogen (AN) were positively correlated with Ace index and Chao1 index of rhizosphere soil fungi. In conclusion, SOM, TN and AN are the main soil environmental factors affecting the rhizosphere soil fungal community of the K. coccinea.
Key words: Kadsura coccinea, ITS, Illumina high-throughput sequencing, fungal community, soil environmental factors, diversity
黑老虎 (Kadsura coccinea)隸屬五味子科(Schisandraceae)南五味子屬(Kadsura),其果實富含多種氨基酸、維生素C、粗蛋白質、可溶性糖等營養(yǎng)成分,是一種歷史悠久的民間藥材植物。近年來,黑老虎因豐富的營養(yǎng)和獨特的風味而深受消費者的歡迎,在貴州黔東南、湖北恩施及湖南通道等南方地區(qū),作為一種新型特色水果,成為脫貧致富的林下產業(yè)(劉濤等,2009;高漸飛等,2022)。黑老虎在醫(yī)藥、保健食品及食品領域展現(xiàn)出廣闊的應用前景(Sritalahareuthai et al., 2020;王麗軍等,2021)。
植物宿主與內生真菌、根際土壤真菌交互作用,形成復雜的調控網絡,共同維持生態(tài)系統(tǒng)的動態(tài)平衡(Jia et al., 2016;Lu et al., 2021)。植物內生真菌生活在宿主體內,不會引起宿主明顯病癥,與宿主協(xié)同進化,利用宿主營養(yǎng)供自身需要,同時在植物生長發(fā)育、增強逆境抵抗能力和生物系統(tǒng)演化等方面發(fā)揮重要作用(Huang et al., 2019)。此外,內生真菌在與宿主的長期相互作用中,通過影響和調節(jié)宿主次生代謝產物的合成及累積影響宿主活性物質的分布和含量,或可以產生與宿主植物類似的次生代謝物,廣泛應用于農學、醫(yī)學、工業(yè)和生物技術等領域(Tiwari & Bae, 2022)。根際真菌一方面利用植物中的碳水化合物進行生長發(fā)育,在有機質轉化、碳-氮循環(huán)和促進土壤環(huán)境健康等方面發(fā)揮重要作用,另一方面其自身新陳代謝所產生的產物反過來可以促進或抑制植物的生長(Bonfante & Anca, 2009; Fierer, 2017)。而同為木蘭科的五味子內生真菌研究有一些報道,五味子屬植物中含有豐富的內生真菌,從不同來源、不同組織中分離的內生菌具有抗氧化、拮抗植物病原菌等功能(潘爭艷等,2007;Gao et al., 2020;張譽薺等,2021)。此外,從五味子中陸續(xù)分離的一些內生真菌能夠產生與宿主植物類似的木脂素及三萜類,或具有轉化宿主次生代謝物的功能(Wang et al., 2017;Qin et al., 2019;Qin et al., 2020)。劉濤等(2023)利用高通量測序技術對黑老虎不同組織內生真菌進行了分析,明確了不同組織內生真菌群落的結構和功能特征。植物內生真菌和根際真菌群落組成和結構受物種、地理環(huán)境、氣候和土壤特性等多種因素影響(Patten & Glick,2002; Chaparro et al., 2013; 王艷等, 2019)。目前,對黑老虎及南五味子屬的研究多集中于營養(yǎng)成分和藥理學研究,而對黑老虎根際真菌和內生真菌組成和多樣性研究尚不多見。
本研究采用ITS高通量測序技術研究了栽培和野生生境下多年生成熟黑老虎根際真菌和根部內生真菌群落結構、組成和多樣性,并開展了真菌多樣性和土壤理化因子的相關性分析,以期為黑老虎資源保護、土壤根際微生態(tài)環(huán)境研究及有益微生物的發(fā)掘和利用提供基礎。
1 材料與方法
1.1 采樣地點及樣品處理
選取貴州省3個黑老虎代表性研究樣地(3個樣地)。貴陽(GY)樣地,貴陽市貴州果樹研究所黑老虎種質資源圃(106°39′38.33″ E、26°29′59.00″ N),海拔1 051 m,為多年生人工管理種質資源圃,土壤類型為中厚層紅黃壤;榕江(RJ)樣地,黔東南苗族侗族自治州榕江縣古州鎮(zhèn)(108°29′15.18″ E、25°55′52.43″ N),海拔361 m,為野生樣地,土壤類型為中厚層紅黃壤為主,富含有機質;福泉(FQ)樣地,黔南布依族苗族自治州福泉市金山街道(107°24′15.48″ E、26°44′36.62″ N),海拔928 m,為野生樣地,土壤類型以石灰石為主,富含P,福泉有“亞洲磷都”之美譽。3個樣地分別代表不同栽培生境、不同土壤類型和不同酸堿度,代表了貴州黑老虎生長的主要生境。
于果實采收季節(jié),分別在3個樣地選取生長多年、藤蔓高度木質化的健康植株,用鏟子去除地表植被和雜質,使用小鋤頭輕輕刨開土壤,尋找主根周邊的細根(直徑≤2 mm),挖取少量帶土的細根(Berhongaray et al., 2013)。每3株樣品合并為1個生物學重復,每個樣本設置3個生物學重復,將樣品收集至無菌采樣袋中并密封保存,迅速置于冷鏈箱內,于4 h內帶回實驗室處理。
將黑老虎根部的土壤抖落,留存附著在根部無法抖落的為根際土(rhizosphere soil,Rh)(Edwards et al., 2015),將根系樣品轉移至無菌50 mL離心管中(含20 mL無菌10 mmol·L-1 PBS溶液),置于搖床 (蘇州培英實驗設備有限公司,培英,型號:THZ-C-1),120 r·min-1,室溫下振蕩20 min(Yu et al., 2013)。使用無菌鑷子挑除根系,剩余懸浮液高速離心 (6 000 g,4 ℃) 20 min,沉淀即為根際土樣品,液氮速凍后,置于-80 ℃冰箱備用。剩余的根系樣品(root,R)用自來水沖去表面多余的根際土,進行表面消毒(75%乙醇浸泡2 min,5%次氯酸處理3 min,無菌水清洗3次)獲得表面消毒的根組織樣品,液氮速凍后,置于-80 ℃冰箱用于DNA 提取。
1.2 土壤理化指標測定
土壤pH值測定:稱取3 g土壤樣品于50 mL燒杯中,加入7.5 mL去除CO2的蒸餾水(水土比2.5∶1),用水平振蕩器劇烈振蕩2 min,使土粒充分分散后,放置30 min后用pH計測定,1 h內完成測定。每個樣品測定3次,計算平均值。土壤總氮(total nitrogen, TN)采用半微量凱氏定氮法,總磷(total phosphorus, TP)采用NaOH堿熔-鉬銻抗分光光度法,土壤總鉀(total potassium, TK)測定采用NaOH熔融法,土壤速效氮(avaliable nitrogen, AN)的測定采用堿解擴散法,土壤速效磷(available phosphorus, AP)的測定采用分光光度法,土壤速效鉀(available potassium, AK)的測定采用乙酸銨浸提法,土壤有機質(soil organic matter, SOM)的測定采用重鉻酸鉀容量法。每個樣品重復3次。
1.3 DNA提取及高通量測序
根系樣品DNA提取采用植物基因組DNA提取試劑盒(貨號:DP305,天根生化科技有限公司,北京)。根際土壤樣品DNA提取采用土壤DNA提取試劑盒(貨號:DP336,天根生化科技有限公司,北京)。ITS引物:fITS7(5′-GTGARTCATCGAA
TCTTTG-3′)和ITS4(5′-TCCTCCGCTTATTGATATG
C-3′)。反應體系 (25 μL):2×Phanta Max master mix 12.5 μL,正、反向引物(1 μmol·L-1)各2.5 μL,DNA μL (50 ng),ddH2O 6.5 μL。PCR 反應條件:94 ℃/1 min-(94 ℃/10 s-50 ℃/30 s-72 ℃/45 s) ×32個循環(huán)-72 ℃/10 min。PCR產物經2%瓊脂糖凝膠電泳確證,用AMPure XT beads (Beckman Coulter Genomics, Danvers, MA, USA)純化,Qubit(Invitrogen, USA)定量。擴增子文庫的大小和數(shù)量分別用Agilent 2100生物分析儀(Agilent,USA)和Illumina (Kapa Biosciences, Woburn, MA,USA)文庫定量試劑盒進行評估。高通量測序委托杭州聯(lián)川生物技術有限公司完成,所用平臺為高通量二代測序儀 (Illumina,HiSeq 4000,USA)。測序原始數(shù)據(jù)已提交至NCBI數(shù)據(jù)庫(項目號為PRJNA855296,樣品編號為SAMN29487930-SAMN29487947)。
1.4 數(shù)據(jù)處理及分析
高通量測序獲得的下機數(shù)據(jù)根據(jù)Barcode拆分后,去除Barcode和接頭序列,并進行質控、過濾,得到Clean data。使用PEAR(v0.9.6)軟件對reads進行拼接合并獲得原始Tags。用Fqtrim篩選Q20>20的序列。利用Vsearch(v2.3.4)去除嵌合體獲得最終有效數(shù)據(jù)用于進一步分析。樣本的α多樣性指數(shù)采用mothur(http://www.mothur.org/wiki/ Schloss_SOP#Alpha_diversity)軟件計算。應用Usearch 軟件(vesion 7.0 http://drive5.com/uparse/)對有效數(shù)據(jù)序列在97%的相似度下進行OTU (Operational Taxonomic Units)聚類,采用Qiime 2進行OTU物種注釋分析,獲得樣品OTU的分類學信息。韋恩圖、PCoA分析、RDA分析等利用聯(lián)川云平臺(https://www.omicstudio.cn/index)進行。利用FUNGuild軟件對真菌進行功能預測分析(Nguyen et al., 2016)。采用SPSS 26對土壤理化性質進行顯著性差異分析。
2 結果與分析
2.1 3個樣地黑老虎根際土壤理化特性
由表1可知,3個不同生境下,黑老虎根際土壤的理化指標存在顯著差異,野生生境土壤主要養(yǎng)分指標高于栽培生境。3個生境中,貴陽栽培生境(GY,pH=4.66)和榕江野生生境(RJ,pH=4.63)下的根際土壤為酸性(pH<6.5),福泉野生生境下的根際土壤pH呈中性(FQ,pH=7.22),主要是由于福泉土壤類型以石灰土最多,土壤pH呈中性,福泉素有“亞洲磷都”之美譽,土壤富含P,此外K含量也較多。RJ根際土壤的TN、AN和SOM含量在3個樣地中最高;FQ根際土壤的TP、TK、AP含量在3個樣地中最高;GY栽培生境下根際土壤TN、TP、TK、AN、AK、AP及SOM含量均顯著低于野生生境(RJ和FQ)。野生生境下的土壤營養(yǎng)元素和有機質含量高于栽培生境,并且生態(tài)系統(tǒng)更為平衡。原因可能為野生生境為茂密的樹林,存在較多腐殖質,有機質含量更為豐富且含水量較高,土壤較為疏松,礦質營養(yǎng)元素不易流失;而人工苗圃在管理過程中施用肥料不多,腐殖質和有機質偏少且土壤存在一定程度的板結,土壤中的N、P、K等元素隨雨水流失較為嚴重。
2.2 黑老虎根際土壤真菌及根部內生真菌測序
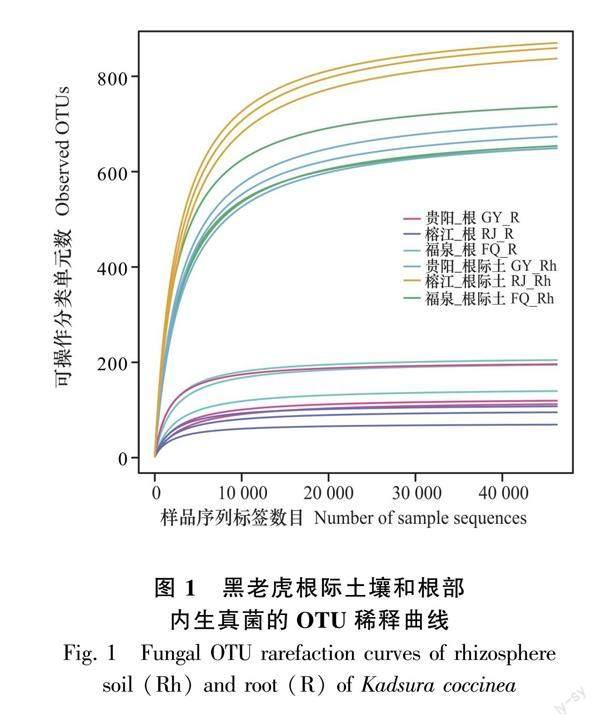
對Illumina高通量測序數(shù)據(jù)進行雙端拼接、質量控制、嵌合體過濾后,進行高質量數(shù)據(jù)統(tǒng)計,共獲得有效序列1 463 619個,樣品有效reads為76 878~84 980對,每個樣品平均為81 312對reads,有效率在93.59%~98.70%之間,Q20全部在99%以上。18個樣品的OTU稀釋曲線隨測序數(shù)目增加逐漸趨于平緩,并且各樣品測序覆蓋度均在99.95%以上(圖1),表明測序數(shù)據(jù)量能較全面地反映測序樣品的真菌群落組成,測序質量評價各項指標均符合ITS測序要求,可以進行后續(xù)分析和研究。
以≥97%的相似性聚類得到4 328個OTU,3個樣地根際土壤真菌和根部內生真菌分別檢測到OTU數(shù)為3 867個和801個,其中根際土壤中各地區(qū)OTU 數(shù)從多到少依次為RJ_Rh(1 358個)> FQ_Rh(1 090個)> GY_Rh(994個),共有的OTU為72個。共有的OTU共注釋到5個門,絕大多數(shù)為子囊菌門(50個,占比68.4%);共注釋到49個屬,占比前幾位的有未分類屬(7個,9.7%)、被孢霉屬(6個,8.3%), 外瓶柄霉屬、 柱孢屬、叢赤殼屬和隱球菌屬均為3個,各占比4.2%。根部內生真菌中,各地區(qū)OTU數(shù)從多到少依次為FQ_R(330)> GY_R(243)>RJ_R(153),共有的OTU為14個。14個共有的OTU共注釋到2個門,子囊菌門有13個,占比92.9%;共注釋到11個屬,占比最高的為外瓶柄霉屬(3個,21.4%),其余屬均為1個,各占比7.1%。根際土壤真菌和內生真菌所有樣品共有的OTU僅為6個,注釋到2個門,子囊菌門為5個,占比83.3%;注釋到5個屬,占比最高的為外瓶霉屬(2個,33.3%),其余分別為被孢霉屬、 柱孢屬和叢赤殼屬(圖2)。圖2結果顯示3個樣地樣品中,真菌OTU數(shù)量均為根際土壤>根內,F(xiàn)Q野生樣地OTU多樣性較高。共有的OTU中,被孢霉屬、外瓶柄霉屬、 柱孢屬和叢赤殼屬最為常見。
2.3 黑老虎根際土壤真菌和根部內生真菌多樣性
Alpha多樣性分析表明,3種生境下,根際土壤的真菌群落多樣性和豐富度均顯著高于根部內生真菌。3個樣地根際土壤真菌中,香農指數(shù)(Shannon index)和辛普森指數(shù)(Simpson index)無顯著差異,RJ_Rh中Chao1指數(shù)和Ace指數(shù)顯著高于GY_Rh和FQ_Rh。3個樣地根部內生真菌中,F(xiàn)Q_R香農指數(shù)顯著高于GY_R和RJ_R,GY_R辛普森指數(shù)顯著低于RJ_R和FQ_R,而RJ_R的Chao1指數(shù)和Ace指數(shù)顯著低于FQ_R,與GY_R無顯著差異(表2)。這說明了3個生境根際土壤真菌多樣性和優(yōu)勢度無顯著差異,RJ_R的根際真菌群落豐度高于其余兩地,3個樣地根部內生真菌中, FQ_R的真菌群落多樣性顯著高于其他兩地,GY_R栽培生境的真菌優(yōu)勢度顯著低于兩個野生生境(RJ_R和FQ_R)。
通過無權重距離矩陣(unweighted_unifrac_distance_matrix)算法對3個樣地樣品進行主坐標分析(PCoA)分析,結果顯示,主坐標1和主坐標2分別解釋了25.25%和13.76%的多樣性,合計解釋了39.01%的多樣性,3個生境下的根部內生真菌群落聚集于第二、第三象限,根際土壤真菌群落聚集于第一、第四象限,顯示3個不同生境中根際土壤真菌更為相似,而根部內生真菌群落更為相似(圖3)。結果表明,黑老虎根部內生真菌受地域影響較小,具有明顯的種屬特異性。另外,從PCoA結果可以看出,同一類樣本的3個生物重復均能很好地聚在一起,說明本研究所采用的樣本重復性好,數(shù)據(jù)可靠性高。
2.4 黑老虎根際土壤和根部內生真菌組成和結構
3個生境下共18個黑老虎根際土壤和根樣品中,共檢測到了真菌12個門,依次為GY(10個)、RJ(9個)和FQ(9個),3個樣地相差不大。3個樣地6組樣品中,真菌優(yōu)勢類群均為子囊菌門(Ascomycota)和擔子菌門(Basidiomycota),其中,子囊菌門占比最高, 占比范圍48.02%~88.38%,在栽培生境(GY_R)的根內(Y1)占比高達88.38%,而在栽培生境的根際土壤(GY_Rh)中占比最低(48.02%)。GY_Rh和FQ_Rh中,存在26.17%和13.68%的未分類真菌(圖4:A)。
在屬水平上,共注釋到509個屬,3個生境中注釋到的真菌屬依次為RJ(375)> FQ(299)>GY(274)。3個生境下,優(yōu)勢屬的結構和豐度存在明顯差異,根部內生真菌中,栽培生境(GY_R)下,外瓶霉屬(Exophiala,77.81%)優(yōu)勢明顯,其他屬占比均不超過4%,2個野生生境下,RJ_R的優(yōu)勢菌群為Cladophialophora(21.68%)、黃絲菌屬(Cephalotheca,19.46%)和Flagelloscypha(19.32%),F(xiàn)Q_R的優(yōu)勢菌群為蠟殼耳目未分類屬(19.7%)、外瓶霉屬(15.20%)和球囊菌門未分類屬(Glomeromycota_unclassified,12.12%)。3個樣地的根際土壤真菌,被孢霉屬(Mortierella)占比均最高,分別為8.57%(GY_Rh)、21.43%(RJ_Rh)和13.53%(FQ_Rh)。此外, GY_Rh優(yōu)勢菌群還有癬囊腔菌屬(Plectosphaerella,4.34%)和外瓶霉 屬(4.15%), RJ_Rh中優(yōu)勢菌群為新叢赤殼屬(Neonectria,7.15%),Y3中優(yōu)勢菌群為癬囊腔菌屬(5.97%)和柱孢屬(Cylindrocarpon,4.31%)(圖4:B)。結果顯示,栽培生境下,根部內生真菌菌群具有一定的偏好性,部分菌群具有優(yōu)越性,而野生生境下,菌群均勻度更高。
以相對豐度Top20的屬制作的熱圖顯示,3個生境下,根部內生真菌和根際真菌群落均存在顯著差異,其中根際土壤真菌群落聚為一類,與PCoA聚類圖結果一致,表明黑老虎根際土壤真菌群落結構受生境影響較小,黑老虎根部內生真菌群落具有一定的物種特異性(圖 5)。
2.5 黑老虎根際土壤真菌及根部內生真菌群落功能預測
利用FUNGuild軟件對黑老虎根際土壤及根部內生真菌進行功能預測,結果顯示真菌7個營養(yǎng)類型在3個生境的根際土壤和根組織中分布不均,其中病理-腐生營養(yǎng)型和腐生營養(yǎng)型兩個類型為優(yōu)勢菌群。3個樣地的根際土壤樣本中,腐生-共生營養(yǎng)型占比相當高, 而在根部內生真菌中占比較低。與內生真菌相比,根際真菌中不同營養(yǎng)類型真菌菌群的分布更為均勻。根部內生真菌中,栽培生境下(GY_R),病理-腐生營養(yǎng)型占比較高,RJ_R的腐生營養(yǎng)型占比較高,F(xiàn)Q_R的共生營養(yǎng)型占比較高(圖 6:A)。
3個生境下6組樣本預測得到61個生態(tài)共位群(guild),不同生態(tài)共位群在3個生境下的根際土壤真菌分布更為相似,GY_R群落中動物病原-真菌寄生蟲-未定義腐生營養(yǎng)型占比較高,而RJ_R中未定義腐生營養(yǎng)型占比較高,F(xiàn)Q_R中動物病原-真菌寄生蟲-未定義腐生營養(yǎng)型和叢枝菌根占比較高(圖6:B)。
2.6 黑老虎根際土壤真菌群落結構與土壤環(huán)境因子相關性
利用土壤理化因子與根部內生真菌和根際真菌群落多樣性指數(shù)分別做Pearson相關性分析。
根部內生真菌中,TK與根部內生真菌香農指數(shù)和辛普森指數(shù)正相關, TP與香農指數(shù)正相關(P<0.05),但未達極顯著水平(圖7:A)。根際土壤真菌中,土壤SOM、AN和TN與根際土壤真菌Ace指數(shù)和Chao1指數(shù)呈極顯著(P<0.01)正相關(圖7:B)。這說明土壤環(huán)境因子對黑老虎內生真菌和根際土壤真菌群落結構的影響不同,TK和TP主要影響內生真菌群落的多樣性,而SOM、TN和AN主要影響根際土壤真菌群落的豐度,對根際土壤真菌群落結構的影響大于內生真菌。
為探究土壤理化因子與根際真菌群落結構的相關性,通過冗余分析(redundancy analysis, RDA)分別研究了土壤理化因子與根際真菌與門水平和豐度排序前20的屬之間的相互關系(圖 8)。在門水平上,第一、第二排序軸累計解釋率分別為62.5%和22.5%, 累計解釋率為85.0%, 能較好地反映黑老虎土壤理化因子與根際真菌門水平的相互關系。SOM、AN、TN和TK與擔子菌門(Basidiomycota)和接合菌門(Zygomycota)呈現(xiàn)較強的正相關。在屬水平上,第一、第二排序軸累計解釋率分別為75.6%和15.0%,累計解釋率為90.6%,能較好地反映黑老虎根際真菌群落與土壤理化因子之間的相互關系。TN、AN、SOM和TK與新叢枝殼屬、黃絲菌屬、被孢霉屬等箭頭方向一致,呈現(xiàn)較強的正相關,與真菌_未分類(Fungi_unclassified)、外瓶霉屬、肉座菌屬(Hypocrea)等箭頭方向相反,呈現(xiàn)明顯的負相關。綜上表明,SOM、TN、AN和TK對黑老虎根際土壤真菌群落豐度影響較大,是影響黑老虎根際土壤真菌結構的主要因素。
3 討論與結論
本研究利用高通量測序技術研究了不同生境下黑老虎根內生真菌及根際土壤真菌群落組成和多樣性,3個樣地根際土壤和根部內生真菌共有的OTU分別為72個和14個,根際土壤真菌和內生真菌所有樣品共有的OTU僅為6個,各地特有的OTU有較大差異。這表明根際土壤真菌具有一定的地域專化性和宿主專一性,與其他物種結果相類似(周婕等,2019)。3個樣地中,黑老虎根際土壤真菌的OTU數(shù)量和多樣性均顯著高于根部內生真菌,土壤中含有微生物所需的食物來源和生長環(huán)境,是地球上微生物含量最為豐富的區(qū)域(Roesch et al.,2007),與重樓(王艷等,2019)、黃精(樊銳鋒等,2021)、川穹(張秫華等,2021)等研究結果一致。此外,3個樣地根際土壤真菌多樣性(香農指數(shù)和辛普森指數(shù))無顯著差異,原因可能為3個樣地均在貴州省內,地理位置、大氣候和生態(tài)環(huán)境具有一定的相似性。但是,3個樣地根際土壤真菌的豐富度(Chao1指數(shù)和Ace指數(shù))存在顯著差異,其中RJ_Rh根際土壤真菌豐富度顯著高于GY_Rh和FQ_Rh,de Boe等(2005)研究表明植物根系及其分泌物、動植物殘體和SOM是土壤真菌可利用的主要能源,榕江土壤豐富的有機質為真菌群落的繁殖提供了豐富的養(yǎng)分,可能是榕江根際土壤真菌豐富度高于另外兩地的原因。3個樣地根部內生真菌中, FQ_R的真菌群落多樣性顯著高于其他兩地,GY_R栽培生境的真菌優(yōu)勢度顯著低于兩個野生生境(RJ_R和FQ_R),可能是因為野生生境受人工干預,栽培生境的植被、土壤結構和營養(yǎng)元素趨于不均衡,導致栽培生境下黑老虎根內生真菌的多樣性偏低。
本研究發(fā)現(xiàn)子囊菌門和擔子菌門是黑老虎根際和根部內生真菌的優(yōu)勢類群,子囊菌門大都為陸生,可分解土壤中動植物腐爛的遺骸和有機質,釋放有機物供植物吸收利用,其含量的高低與土壤肥力有關。擔子菌門常與植物形成共生菌根,促進植物的生長和發(fā)育(Miguel et al., 2020)。本研究結果與五味子屬(江林春等,2020)及其他中藥材如重樓(王艷等,2019)、黃精(樊銳鋒等,2021)、川穹(張秫華等,2021)等一致。在屬水平上,根部內生真菌中, GY_R、RJ_R和FQ_R的優(yōu)勢菌屬分別為外瓶霉屬(77.81%)、Cladophialophora(21.68%)和蠟殼耳目未分類屬(19.7%)、外瓶霉屬(15.20%)。外瓶霉屬在栽培生境(GY)的根中豐度較高(77.81%)。其原因可能為外瓶霉屬有在土壤、水和空氣等不同生態(tài)系統(tǒng)中有著廣泛的分布,多為人、畜和水產魚類等的真菌病害,在植物中,有耐重金屬特性的報道(王璐,2011)。栽培生境下,長期施用化肥農藥,受到人類活動干擾較多,外瓶霉屬上升為優(yōu)勢菌群。根際土壤真菌中,3個樣地的優(yōu)勢菌屬均含被孢霉屬,而其他優(yōu)勢菌群各不相同。被孢霉屬為土壤碳及養(yǎng)分轉化的關鍵成員,在植物生長發(fā)育、代謝中均發(fā)揮關鍵作用(寧琪等,2022)。在屬水平上,對不同生境下根際土壤和根部內生真菌的α多樣性分析顯示,不同生境下黑老虎根際土壤和根部內生真菌群落的豐度和多樣性存在顯著差異。FUNGuild功能預測結果顯示,7個營養(yǎng)類型在6組樣本中分布不均,栽培生境下(GY)病理-腐生營養(yǎng)型及動物病原-真菌寄生蟲-未定義腐生營養(yǎng)型真菌在黑老虎根內生菌中比例最高,推測可能是由于在栽培生境下,由于生態(tài)平衡遭到破壞,造成植株體內病原菌和寄生蟲等侵害機會增加,病原菌潛伏或以條件性致病方式寄生于植物根內,是導致栽培生境下植物更容易發(fā)生病害的原因之一(Rodriguez et al., 2009)。王艷等(2019)發(fā)現(xiàn),野生生境下的重樓內生真菌和土壤真菌菌群豐度和多樣性顯著高于園區(qū)環(huán)境,與本研究結果較為一致。
植物根際土壤和根內真菌在植物生長發(fā)育、低于逆境和維持土壤微生態(tài)環(huán)境中發(fā)揮重要作用,其結構和組成受土壤環(huán)境因子如濕度、pH值、N、P、K、SOM等理化因素的影響(Roesch et al., 2007)。本研究結果顯示,3個不同生境下,黑老虎根際土壤的理化指標存在明顯差異,野生生境下的土壤營養(yǎng)元素和有機質含量高于栽培生境,說明栽培生境下, 要注意合理施用營養(yǎng)元素和有機肥,確保植物生長的需要和土壤生態(tài)環(huán)境平衡。同時,本研究結果也為后續(xù)的人工苗圃管理提供了重要信息。土壤理化因子與根部內生真菌多樣性相關分析顯示,土壤TK和TP與根部內生真菌香農指數(shù)相關顯著,表明TK和TP主要影響根部內生真菌的多樣性,對豐度和優(yōu)勢度影響不明顯。根際土壤真菌中,土壤SOM、AN和TN與根際土壤真菌Ace和Chao1指數(shù)極顯著相關。這表明土壤環(huán)境因子對根際土壤真菌群落結構影響大于根部內生真菌且主要影響根部內生真菌的多樣性,而對根際土壤真菌的影響主要為豐度。李銘杰等(2021)發(fā)現(xiàn),土壤SOM、pH、AP、AK與北蒼術根區(qū)土壤叢枝菌根真菌Shannon指數(shù)、Simpson指數(shù)呈正相關,其中SOM對叢枝菌根真菌群落多樣性影響最大。楊蕊毓等(2022)研究發(fā)現(xiàn),川麥冬根圍土壤叢枝菌根多樣性指數(shù)及群落組成與土壤理化因子TK、TP、TN相關性顯著。金輝等(2021)對紫花針茅根際土壤和內生真菌研究分析發(fā)現(xiàn),土壤理化因子pH、AK、Fe、Ca 和Mg 與Ace指數(shù)、Chao1指數(shù)和Shannon指數(shù)等真菌多樣性指數(shù)顯著相關(P<0.05)或極顯著相關(P<0.01),與本研究結果具有部分相似性。N是植物生長發(fā)育所需的大量元素,植物通過根系分泌物和物生物互作,形成植物-土壤的氮素化學循環(huán),因此N與土壤微生物群落和根內生菌均發(fā)生交互作用(Hobbie et al., 2007)。有機質含量影響土壤真菌的群落結構,主要由于SOM在微生物和酶的作用下發(fā)生分解,同時為土壤微生物提供豐富的養(yǎng)分來促進了土壤真菌的生長 (隋躍宇等, 2009; Smith et al., 2014)。
土壤TN、AN、SOM和TK是影響黑老虎根際土壤真菌結構的主要因素。RDA分析結果顯示,被孢霉屬與TN、AN和SOM呈現(xiàn)較強的正相關。被孢霉屬是接合菌亞門接合菌綱毛霉目被孢霉科,能顯著提高土壤可溶性有機碳、AN和AP的含量(寧琪等,2022)。本研究結果表明,多施有機肥、氮肥和鉀肥,可以改善土壤微生物菌群結構,促進黑老虎生長發(fā)育。本研究為今后黑老虎根內有益真菌的分離與鑒定提供了重要信息,同時也為黑老虎的引種馴化和生態(tài)栽培提供了理論依據(jù)。
參考文獻:
BERHONGARAY G, JANSSENS IA, KING JS, et al., 2013. Fine root biomass and turnover of two fast-growing poplar genotypes in a short-rotation coppice culture [J]. Plant Soil, 373(1-2):269-283.
BONFANTE P, ANCA IA, 2009. Plants, mycorrhizal fungi, and bacteria: a network of interactions [J]. Ann Rev Microbiol, 63: 363-383.
CHAPARRO JM, BADRI DV, VIVANCO JM, 2013. Rhizosphere microbiome assemblage is affected by plant development [J]. ISME J, 8(4):? 790-803.
DE BOER W, FOLMAN LB, SUMMERBELL RC, et al., 2005. Living in a fungal world:? impact of fungi on soil bacterial niche development [J]. FEMS Microbiol Rev, 29(4):? 795-811.
EDWARDS J, JOHNSON C, SANTOS-MEDELLíN C, et al., 2015. Structure, variation, and assembly of the root-associated microbiomes of rice [J]. Proc Natl Acad Sci, 112(8): E911-920.
FAN RF, WANG RF, DU YQ, et al., 2021. Community composition and ecological function of rhizosphere fungi and medicinal parts endophytic fungi in Polygonatum sibiricum [J]. Guihaia, 41(5): 799-807.? [樊銳鋒, 王若凡, 杜艷秋, 等, 2021. 黃精根際及藥用部位內生真菌群落組成和生態(tài)功能分析 [J]. 廣西植物, 41(5): 799-807.]
FIERERN, 2017. Embracing the unknown:? disentangling the complexities of the soil microbiome [J]. Nat Rev Microbiol, 15(10): 579-590.
GAO JF, ZHOU W, LIU N, 2022. Differential analysis of phenolic acids from different parts of Kadsura coccinea? [J]. J Chin Med Mat, 45(2): 351-358.? [高漸飛, 周瑋, 劉妮, 2022. 黑老虎不同部位酚酸類成分差異分析 [J]. 中藥材, 45(2): 351-358.]
GAO Y, ZHOU J, RUAN HL, 2020. Trichothecenes from an endophytic fungus Alternaria sp. sb23 [J]. Planta Med, 86(13/14): 976-982.
HOBBIE SE, OGDAHL M, CHOROVER J, et al., 2007. Tree species effects on soil organic matter dynamics:? the role of soil cation composition? [J]. Ecosystems, 10(6):? 999-1018.
HUANG SZ, DUAN LP, WANG H, et al., 2019. Two new AChE inhibitors isolated from Li folk herb Heilaohu “Kadsura coccinea” stems? [J]. Molecules, 24(19): 3628.
JIA M, CHEN L, XIN HL, et al., 2016. A friendly relationship between endophytic fungi and medicinal plants:? a systematic review [J]. Front Microbiol, 7: 906.
JIANG LC, WEI JM, BAI SS, et al., 2020. Characteristics of rhizosphere soil fungi community of Schisandra sphenanthera Rehd. Et Wils [J]. J Yanan Univ (Nat Sci Ed), 39(4): 15-20.? [江林春, 韋潔敏, 白尚尚, 等, 2020. 華中五味子根際土壤真菌群落特征研究 [J]. 延安大學學報(自然科學版), 39(4): 15-20.]
JIN H, YANG XY, LIU HY, et al., 2021. The structure of rhizosphere soil and endophytic fungal communities associated with Stipa purpurea and their correlation with soil environmental factors [J]. Acta Microbiol Sin, 61(11): 3520-3541.? [金輝, 楊曉燕, 柳皓月, 等, 2021. 紫花針茅根際和體內真菌群落結構及與土壤環(huán)境因子的相關性研究 [J]. 微生物學報,? 61(11): 3520-3541.]
LI MJ, ZHOU ZJ, XING LJ, et al., 2021. Correlation analysis between diversity of AM fungi and nutrient in root-zone soil of Atractylodes chinensis [J]. Chin J Biol Control, 37(6): 1288-1297.? [李銘杰, 周志杰, 邢禮軍, 等, 2021. 北蒼術根區(qū)土壤中AM真菌多樣性及其與土壤養(yǎng)分相關性分析 [J]. 中國生物防治學報, 37(6): 1288-1297.]
LIU T, MA N, JIN JF, et al., 2023. Composition structure and ecological function analyses of endophytic and rhizosphere soil fungi in Kadsura coccinea [J]. Guihaia, 43(5): 869-879.? [劉濤, 馬楠, 金吉芬, 等, 2023. 黑老虎內生真菌及根際土壤真菌的群落結構與生態(tài)功能分析? [J]. 廣西植物, 43(5): 869-879.]
LIU T, WEI Q, WU XB, et al., 2009. Research status and development and utilization prospect of wild fruit species Kadsura coccinea (Lem.) A. C. Smith [J]. J Anhui Agric Sci, 37(23): 10971-10972.? [劉濤, 韋茜, 吳小波, 等, 2009. 野生水果黑老虎的研究現(xiàn)狀及開發(fā)利用前景 [J]. 安徽農業(yè)科學, 37(23): 10971-10972.]
LU HY, WEI TY, LOU HH, et al., 2021. A critical review on communication mechanism within plant-endophytic fungi interactions to cope with biotic and abiotic stresses [J]. J Fung (Basel), 7(9): 719.
MIGUEL RM, JOSE GMV, MEIKE P, et al., 2020. Diversity of fungi in soils with different degrees of degradation in Germany and Panama? [J]. Mycobiology, 48(1): 20-28.
NGUYEN NH, SONG Z, BATES ST, et al., 2016. FUNGuild:? An open annotation tool for parsing fungal community datasets by ecological guild [J]. Fung Ecol, 20:? 241-248.
NING Q, CHEN L, LI F, et al., 2022. Effects of Mortierella on nutrient availability and straw decomposition in soil [J]. Acta Pedol Sin, 59(1): 206-217.? [寧琪, 陳林, 李芳, 等, 2022. 被孢霉對土壤養(yǎng)分有效性和秸稈降解的影響 [J]. 土壤學報, 59(1):? 206-217.]
PAN ZY, FU JF, ZHOU RJ, et al., 2007. Preliminary study of soil fungi in Schisandra chinensis garden and screening antagonism strains [J]. J Jilin Agric Univ, 29(6): 636-639.? [潘爭艷, 傅俊范, 周如軍, 等, 2007. 五味子園根際真菌多樣性初探及拮抗菌株篩選 [J]. 吉林農業(yè)大學學報, 29(6): 636-639.]
PATTENCL, GLICK BR, 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system [J]. Appl Environ Microb, 68(8): 3795-3795.
QIN D, SHEN WY, WANG JQ, et al., 2019. Enhanced production of unusual triterpenoids from Kadsura angustifolia fermented by a symbiont endophytic fungus, Penicillium sp. SWUKD4.1850 [J]. Phytochemistry, 158: 56-66.
QIN D, SHEN WY, GAO TC, et al., 2020. Kadanguslactones A-E, further oxygenated terpenoids from Kadsura angustifolia fermented by a symbiotic endophytic fungus, Penicillium ochrochloron SWUKD4.1850 [J]. Phytochemistry, 174: 112335.
RODRIGUEZ RJ, WHITE JF, ARNOLD AE, et al., 2009. Fungal endophytes:? diversity and functional roles [J]. New Phytol, 182(2): 314-330.
ROESCH LF, FULTHORPE RR, RIVA A, et al., 2007. Triplett EW. Pyrosequencing enumerates and contrasts soil microbial diversity [J]. ISME J, 1(4):? 283-290.
SMITH AP, MARíN-SPIOTTA E, DE GRAAFF MA, et al., 2014. Microbial community structure varies across soil organic matter aggregate pools during tropical land cover change [J]. Soil Biol Biochem, 77:? 292-303.
SRITALAHAREUTHAI V, TEMVIRIYANUKUL P, ON-NOM N, et al., 2020. Phenolic profiles, antioxidant, and inhibitory activities of Kadsura heteroclita (Roxb.) Craib and Kadsura coccinea (Lem.) A.C. Sm [J]. Foods, 9(9): 1222.
SUI YY, JIAO XG, GAO CS, et al., 2009. The relationship among organic matter content and soil microbial biomass and soil enzyme activities [J]. Chin J Soil Sci, 40(5):? 1036-1039. [隋躍宇, 焦曉光, 高崇生, 等, 2009. 土壤有機質含量與土壤微生物量及土壤酶活性關系的研究 [J]. 土壤通報, 40(5):? 1036-1039.]
TIWARI P, BAE H, 2022. Endophytic fungi:? key insights, emerging prospects, and challenges in natural product drug discovery [J]. Microorganisms, 10(2): 360.
WANG L, 2011. Resistance screening on heavy metal and taxonomic studies of strains belong to Phialophora and Exophiala? [D]. Kumming: Yunnan University.? [王璐,? 2011. 瓶霉屬(Phialophora)和外瓶霉屬(Exophiala)菌株重金屬耐性篩選及分類學研究 [D]. 昆明: 云南大學.]
WANG L, QIN D, ZANG K, et al., 2017. Metabolites from the co-culture of nigranoic acid and Umbelopsis dimorpha SWUKD3.1410, an endophytic fungus from Kadsura angustifolia [J]. Nat Prod Res, 31(12): 1414-1421.
WANG LJ,LIAO SQ, LIANG J, et al., 2021. Analysis and evaluation of nutritional components of Kadsura coccinea seeds [J]. Chin Oils Fats, 46(12): 112-117.? [王麗軍, 廖蘇奇, 梁潔, 等, 2021. 黑老虎種子的營養(yǎng)成分分析及評價 [J]. 中國油脂, 46(12): 112-117.]
WANG Y, CHANG F, CHENG HY, et al., 2019. Fungal diversity and differential analysis of fungi community composition in Paris polyphylla var. chinensis rhizosphere and endophytic [J]. Chin Trad Herb Drugs, 50(5): 1232-1237.? [王艷, 常帆, 程虎印, 等, 2019. 重樓根際及藥用部位內生真菌多樣性與群落結構差異分析 [J]. 中草藥, 50(5): 1232-1237.]
YANG RY, DENG QZ, TIAN LP, et al., 2022. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of Ophiopogon japonicus in different habitats [J]. Acta Bot Boreal-Occident Sin, 42(1): 145-153.? [楊蕊毓, 鄧錡璋, 田麗平, 等, 2022. 不同生境下川麥冬根圍土壤叢枝菌根真菌多樣性 [J]. 西北植物學報,? 42(1): 145-153.]
YU J, ZHOU XF, YANG SJ, et al., 2013. Design and application of specific 16S rDNA-targeted primers for assessing endophytic diversity in Dendrobium officinale using nested PCR-DGGE [J]. Appl Microbiol Biotechnol, 97(22): 9825-9836.
ZHANG SH, FANG Q, JIA HM, et al., 2021. Difference analysis of fungal community among bulk soil, rhizosphere, and rhizomes of Ligusticum chuanxiong Hort [J]. Biotechnol Bull, 37(4): 56-69.? [張秫華, 方千, 賈紅梅, 等, 2021. 川芎非根際、根際及根莖內生真菌群落差異分析 [J]. 生物技術通報, 37(4): 56-69.]
ZHANG YQ, XU YH, WEN XS, et al., 2021. Response surface methodology for optimizing fermentation process of the endophytic fungus Purpureocillium lilacinum WG9 preventing Schisandra chinensis brown spot disease [J]. Mycosystema, 40(6): 1561-1574.? [張譽薺, 許永華, 文湘穗, 等, 2021. 響應面法優(yōu)化北五味子褐斑病內生生防真菌淡紫擬青霉WG9發(fā)酵工藝及發(fā)酵產物穩(wěn)定性研究 [J]. 菌物學報, 40(6): 1561-1574.]
ZHOU J, MIAO YF, FANG K, et al., 2019. Diversity of the endophytic and rhizosphere soil fungi of Ageratina adenophora [J]. Ecol Sci, 38(5):? 1-7.? [周婕, 苗一方, 方楷, 等, 2019. 紫莖澤蘭內生真菌及其根際土壤真菌的多樣性研究 [J]. 生態(tài)科學, 38(5): 1-7.]
(責任編輯 李 莉 王登惠)
