基于高通量測序技術探究套種靈芝的茶樹根際土壤真菌群落結構特征
2023-08-30 14:19:18韓海東周柳婷黃小云俞成然黃秀聲
茶葉科學 2023年4期
韓海東,周柳婷,黃小云,俞成然,黃秀聲
基于高通量測序技術探究套種靈芝的茶樹根際土壤真菌群落結構特征
韓海東,周柳婷*,黃小云,俞成然,黃秀聲*
福建省農業科學院農業生態研究所/福建省丘陵地區循環農業工程技術研究中心,福建 福州 350013
茶園套種靈芝是將茶葉加工的廢棄物資源化再利用的生態循環間作模式,所形成的復合群體可以更徹底地覆蓋地面,在改善土壤微生物群落結構,維持土壤微生態系統平衡等方面發揮重要作用。以未套種(CK)、套種1年(A1)、套種2年(A2)、套種3年(A3)的茶樹根際土壤為研究對象,利用Miseq PE300高通量測序技術探究土壤真菌群落結構變化特征。結果表明,與CK相比,套種靈芝顯著提高了茶樹根際土壤堿解氮、速效磷、速效鉀及有機碳含量,其中A3土壤升幅最高,各養分含量分別達32.36%、13.01%、69.21%和9.56%。多樣性指數表明,茶樹根際土壤真菌群落Observed species指數和Chao1指數呈現CK>A3>A1>A2;ACE指數、Shannon指數和Simpson指數呈現A3>CK>A1>A2。多樣性指數表明,A2與CK、A1、A3的茶樹根際土壤真菌群落組成結構差異相對較大。分類學分析發現,茶樹根際土壤真菌分布于18門48綱135目309科632屬。在門水平上,子囊菌門(Ascomycota)是CK、A1和A3的優勢菌門,相對豐度分別為71.28%、68.74%和51.79%;擔子菌門(Basidiomycota)是A2的優勢菌門,相對豐度為64.48%。在屬水平上,與CK相比,套種靈芝的茶樹根際土壤A1中角擔菌屬()、被孢霉屬()、毛孢子菌屬()和三形菌屬()分別顯著上升59.14、1.34、3.70倍和1.92倍(<0.05);A2土壤的古根菌屬()顯著下降76.81%,而棉革菌屬()和支孢瓶霉屬()分別顯著上升788.43倍和36.24倍(<0.05);A3土壤的被孢霉屬和靈芝屬()分別顯著上升1.09倍和0.81倍(<0.05)。綜上所述,茶園套種靈芝可有效調節茶樹根際土壤真菌群落組成結構,改善茶園土壤微生態環境,為茶園可持續綠色發展提供理論依據。
茶園;套種靈芝;根際土壤;真菌群落結構
福建省茶產業發展歷史悠久,茶產業既是千億產業鏈“三產融合”的特色產業,也是我國助力全面推進新農村發展和鄉村振興的優勢產業[1]。然而,由于福建省“八山一水一分田”的地形特征,大部分茶園由丘陵山地開發而來,茶農為了追求茶葉產量而不斷進行茶園徹底清雜,使茶園生物多樣性遭受嚴重破壞和損失,導致土壤養分流失,保水透氣性降低。此外,茶樹屬于喜酸富鋁植物,進一步加劇了茶園土壤酸化、微生物數量減少、微生物群落多樣性降低等一系列生態環境問題,嚴重威脅茶樹根際土壤質量[2]。
科學的間套作模式是茶園生境管理的有效措施之一,可以提高茶園生物多樣性,改善和豐富土壤微生物種類、數量及活性,提高茶園保益控害的生態系統服務功能[3]。研究表明,茶園間套種白三葉草、黑麥草、圓葉決明、平托花生、百喜草等綠肥作物后,土壤微生物種類和數量顯著增加,尤其土壤中有益真菌菌屬的相對豐度顯著提高[4-8]。茶園間作食用菌有利于提高水、熱、光和空間等生境資源的利用率,進而改變土壤微生物生命活動的物質基礎和生存條件[9-10]。有研究表明,茶樹套種長根菇的間作模式可顯著提高土壤速效氮、磷、鉀含量,但對土壤有機質和pH影響不明顯[11]。間作靈芝后茶園土壤變形菌門(Proteobacteria)相對豐度顯著提高21.18%,而酸桿菌門(Acidobacteria)和芽單胞菌門(Gemmatimonadetes)的相對豐度顯著降低15.09%和53.52%;在屬水平上,間作靈芝處理顯著提高了土壤有益微生物菌屬相對豐度[12]。目前關于茶園套種食用菌間作模式的研究多集中于土壤理化性質、茶葉品質及產量、土壤細菌群落種類和數量等方面,對茶樹根際土壤真菌群落組成結構的研究未見報道。
高通量測序技術具有通量高、靈敏度高、準確度高、樣本量較低等優點,可以更深入全面地揭示環境中微生物組成結構及多樣性變化[13]。因此,本研究以未套種(CK)、套種1年(A1)、套種2年(A2)和套種3年(A3)的茶樹根際土壤為研究對象,采用Miseq PE300高通量測序技術探討套種靈芝對茶樹根際土壤真菌群落結構的影響,為該生態循環間作模式服務茶產業可持續發展提供理論依據。
1 材料與方法
1.1 試驗小區概況與設計
1.1.1 試驗區域概況
試驗樣地選址于武夷山市張天福有機茶園示范基地(117°5'50'' E,27°38'21'' N)。該基地占地面積3?000?m2,地處中亞熱帶季風濕潤氣候區,平均海拔335?m。試驗茶樹品種為奇丹,樹齡5年。試驗區土壤質地為砂壤土,理化性質如下:pH 5.60、有機質35.70?g·kg-1、全氮1.75?g·kg-1、全磷0.95?g·kg-1、全鉀7.23?g·kg-1、堿解氮174.06?mg·kg-1、速效磷76.30?mg·kg-1、速效鉀266.01?mg·kg-1。
1.1.2 試驗小區設計
在試驗園區內地形地勢及茶樹長勢基本一致的山地上設置4個樣地:未套種(CK)、套種1年(A1)、套種2年(A2)和套種3年(A3),每個樣地隨機設3個100?m2的重復小區,共12個小區,小區間設保護行。分別于2017年(A3)、2018年(A2)、2019年(A1)開溝套種靈芝。套種方法:每年5月上旬茶葉采收完畢后,在每個小區的茶樹樹冠垂直下方土層開鑿約20?cm深溝,將250根靈芝菌棒脫去塑料外膜并填埋于深溝內,覆土。每年7月中上旬采摘靈芝,菌棒留置于土壤中。靈芝菌棒質量約2?kg,長度約40?cm,配方為70%茶梗、10%木粉、15%麥麩及5%其他,有機碳和有機氮含量分別為165?g·kg-1和3.2?g·kg-1。
1.2 試驗方法
1.2.1 土壤樣品采集
2020年9月,在每個小區內隨機選擇5株長勢相近的茶樹,逐層鏟除表層覆土,剪下茶樹細根分枝并輕輕抖落根圍土,用小毛刷收集仍緊貼于細根表面的根際土壤至無菌袋中,并迅速用液氮冷凍保存。將每個小區內取得的5份根際土壤混勻為1份土樣,共獲得12份土壤樣品。土樣通過2?mm篩去除細根、碎石等雜物后,將每份土樣混勻分成2份裝入無菌袋,每份5?g,一份保存于4?℃冰箱用于測定土壤基礎理化指標,另一份用液氮冷凍運輸至北京奧維森基因科技有限公司進行ITS1 MiSeq PE300測序。
1.2.2 土壤真菌群落結構分析
根際土壤真菌DNA提取與PCR擴增:采用PowerSoil DNA Isolation Kit試劑盒(MoBio Laboratories,Inc.,CA)抽提根際土壤基因組DNA,利用1%瓊脂糖凝膠電泳檢測基因組DNA片段大小,并通過Nanodrop 2000(Thermo Scientific,美國)測定DNA濃度。以稀釋至1?ng·μL-1的基因組DNA為模板,對土壤真菌ITS1區進行PCR擴增,擴增引物為ITS1-F(5'-CTTGGTCATTTAGAGGAAGTA A-3')和ITS2-R(5'-TGCGTTCTTCATCGATGC-3'),采用1%瓊脂糖凝膠電泳檢測擴增產物質量。
1.3 數據分析
通過Illumina MiSeq平臺進行Paired-end測序,下機數據經QIIME(v 1.8.0)軟件過濾、拼接、去除嵌合體后,剔除堿基模糊、引物錯配或長度小于150?bp的低質量序列。利用QIIME(v 1.8.0)和Uclust(v 1.2.22)對97%相似度的有效序列分配運算分類單元(Operational taxonomic unit,OTU),與Silva數據庫比對后,得到每個OTU對應的物種分類信息。運用Mothur軟件(v 1.31.2)進行多樣性分析;基于Weighted Unifrace距離矩陣,利用R(v 3.1.1)軟件包的非加權組平均法(UPGMA)進行聚類分析;通過Bray-Curtis算法計算多樣性距離矩陣,基于系統進化信息比較各樣品間的物種群落差異。具體的數據分析軟件和算法參考北京奧維森基因科技有限公司官方網站的說明。使用SPSS 16.0進行數據方差分析,通過LSD比較法進行差異顯著性分析(<0.05)。
2 結果與分析
2.1 套種靈芝后茶樹根際土壤理化性質變化特征
由表1可知,套種靈芝的栽培模式提高了茶樹根際土壤pH值,但差異不顯著,即茶園套種靈芝在一定程度上可能會緩解土壤酸化現象。4個土壤樣品中,A2土壤的pH值最高(5.83),CK的pH值最低(5.63)。與CK相比,堿解氮、速效磷、速效鉀及有機碳含量在A1、A2和A3的土壤中均顯著上升(<0.05),且在A3根際土壤中的含量均最高,分別比CK顯著上升32.36%、13.01%、69.21%和9.56%;A2和A3茶樹根際土壤的全氮和全磷顯著高于CK和A1,全氮含量最高的A3比A1顯著上升22.83%,全磷含量最高的A2比CK顯著上升71.13%(<0.05);與CK相比,A1和A2的微生物量碳含量顯著下降27.16%和21.84%(<0.05),在A3中略有上升,但差異不顯著。
2.2 套種靈芝后茶樹根際土壤真菌群落α多樣性分析
多樣性是常用于反映微生物群落豐富度和均勻性的綜合指標。本試驗共產生4?069個OTU,經過抽平處理剩余3?797。基于抽平后的OTU聚類結果進行多樣性分析,結果表明,隨著測序深度增加,Shannon-Wiener稀釋曲線趨于平坦(圖1),表明樣本測序量已基本飽和,能夠反映樣本大部分真菌的物種信息。由表2可知,與CK相比,A1和A2茶樹根際土壤觀察到的物種數、Chao1指數、ACE指數、Shannon指數和Simpson指數均隨著靈芝套種年限增加而降低,A2的降幅分別為32.39%、24.21%、22.85%、41.99%和29.90%。與A1和A2相比,A3觀察到的物種數、Chao1指數、ACE指數、Shannon指數和Simpson指數均升高,其中A3觀察到的物種數、ACE指數和Shannon指數與A1和A2差異顯著(<0.05)。A3茶樹根際土壤的ACE指數、Shannon指數和Simpson指數均最高,分別比最低的A2顯著提高29.72%、83.58%和44.12%。
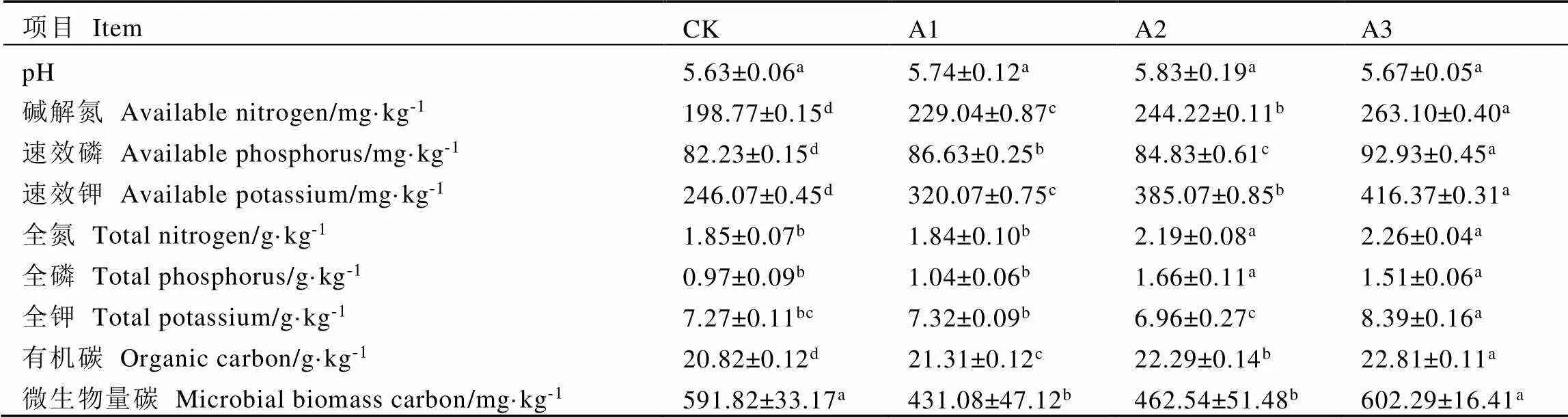
表1 不同處理茶樹根際土壤理化性質
注:每行不同字母表示差異達顯著水平(<0.05,n=3)
Note: Different letters in each line indicate significant differences (< 0.05, n=3)
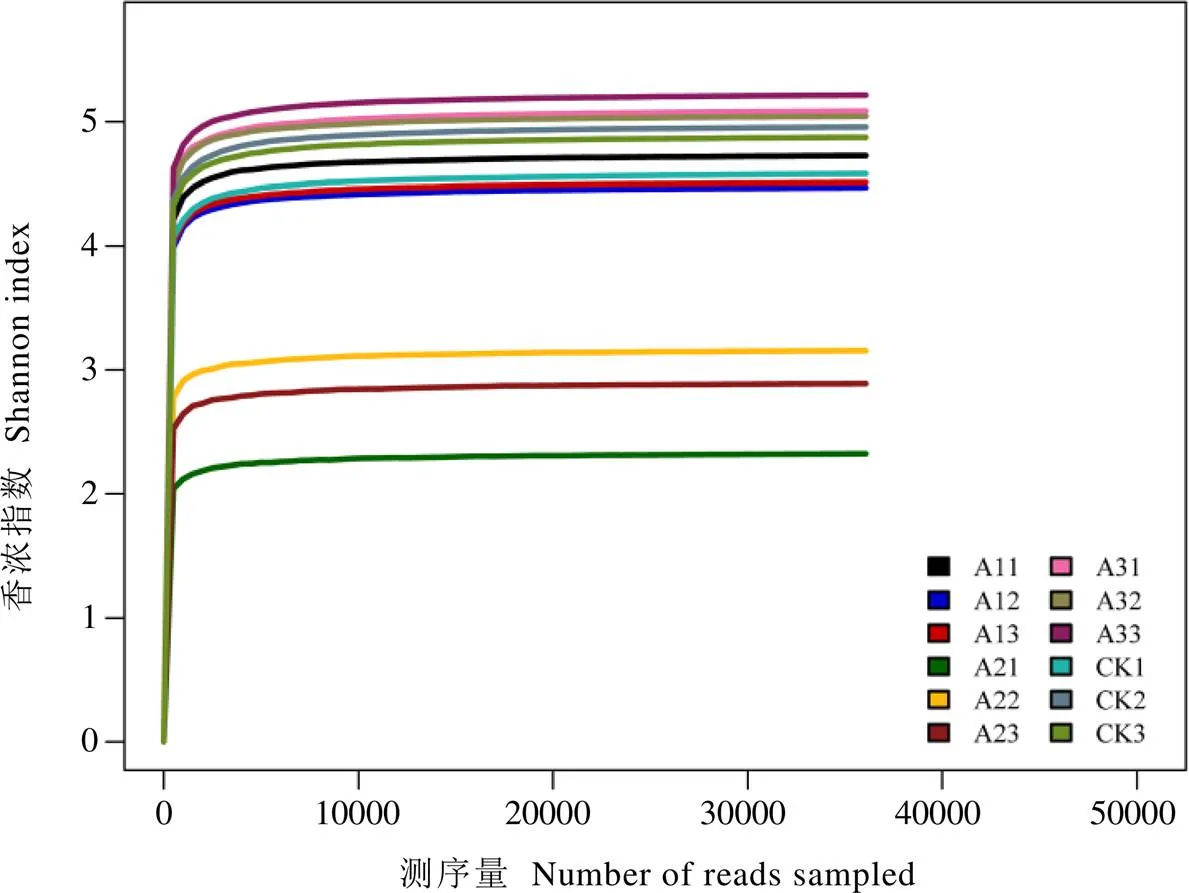
圖1 不同處理茶樹根際土壤真菌群落Shannon-Wiener曲線

表2 不同處理茶樹根際土壤真菌群落多樣性指數
注:每列不同字母表示差異達顯著水平(<0.05,n=3)
Note: Different letters in each column indicate significant differences (< 0.05, n=3)
2.3 套種靈芝后茶樹根際土壤真菌群落β多樣性分析
基于OTU的茶樹根際土壤真菌群落PCA分析如圖2所示,相同顏色的3個重復樣本聚集在一起,說明不同處理組內生物學重復性較好。主成分1(PC1)與主成分2(PC2)分別解釋變量方差的45.64%和25.21%,兩者累計貢獻率達70.85%。PC1將A2的OTU群落與CK、A1、A3明顯區分開,PC2將A1、A2的OTU群落與CK、A3明顯區分開。CK與A3距離較近,與A1、A2距離較遠,說明套種靈芝菌棒1年和2年對茶樹根際土壤真菌群落多樣性的影響較大,套種3年影響程度減小。由層次聚類分析可知(圖3),CK土壤的OTU群落首先與A3聚集為1個群體,之后再與A1聚集,最后與A2聚集形成系統發生樹。多樣性分析結果表明,不同處理的茶樹根際土壤真菌群落組成結構存在較大差異,尤其是CK土壤與A2土壤的真菌群落多樣性差異較為明顯。
2.4 套種靈芝后茶樹根際土壤真菌群落組成結構變化
2.4.1 基于門水平不同處理茶樹根際土壤真菌群落結構
在茶樹根際土壤真菌群落中共檢測到18個門,其中,相對豐度前5位的優勢類群依次是子囊菌門(Ascomycota,29.01%~71.28%)、擔子菌門(Basidiomycota,13.51%~64.48%)、未定義菌門(unidentified,3.55%~9.28%)、被孢霉門(Mortierellomycota,1.01%~3.19%)和球囊菌門(Glomeromycota,1.39%~2.78%)(表3)。子囊菌門是CK、A1和A3土壤的優勢菌門,相對豐度分別為71.28%、68.74%和51.79%;而A2土壤的優勢菌門為擔子菌門,相對豐度為64.48%。
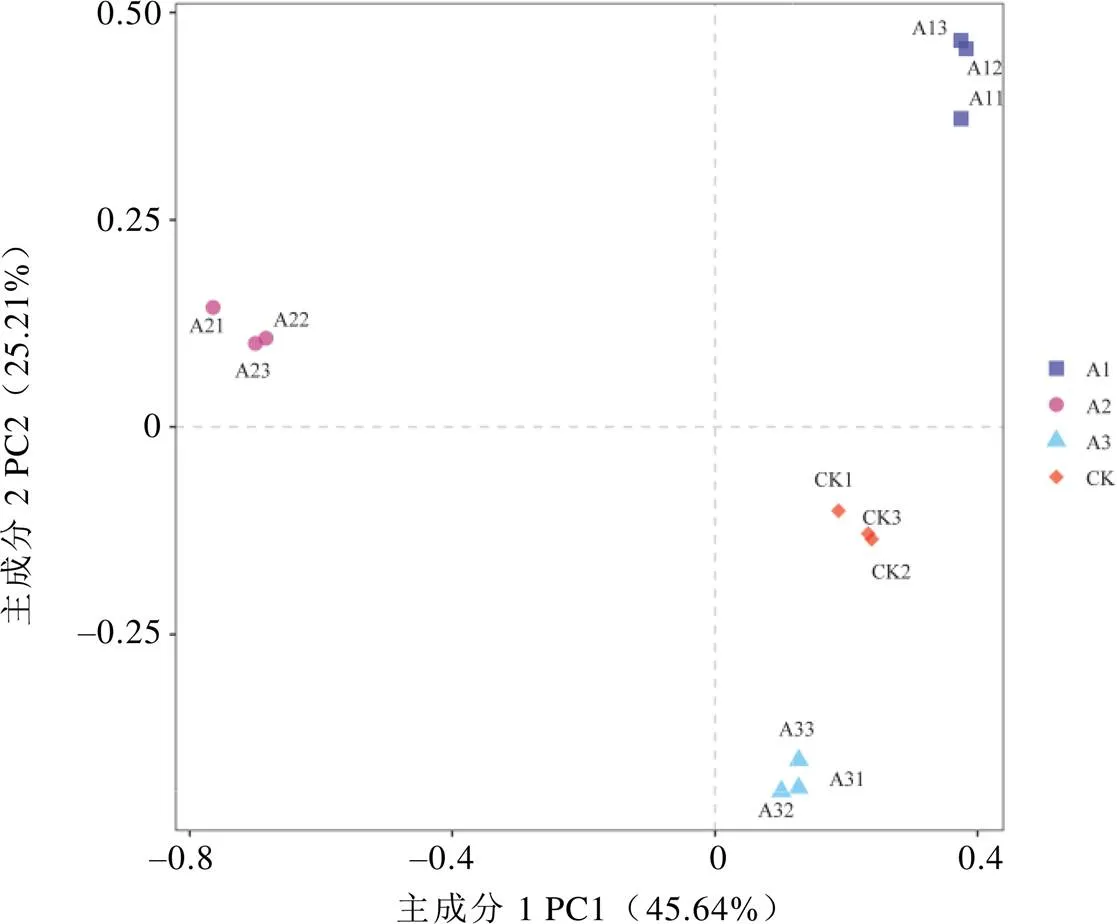
圖2 不同處理茶樹根際土壤真菌群落PCA分析
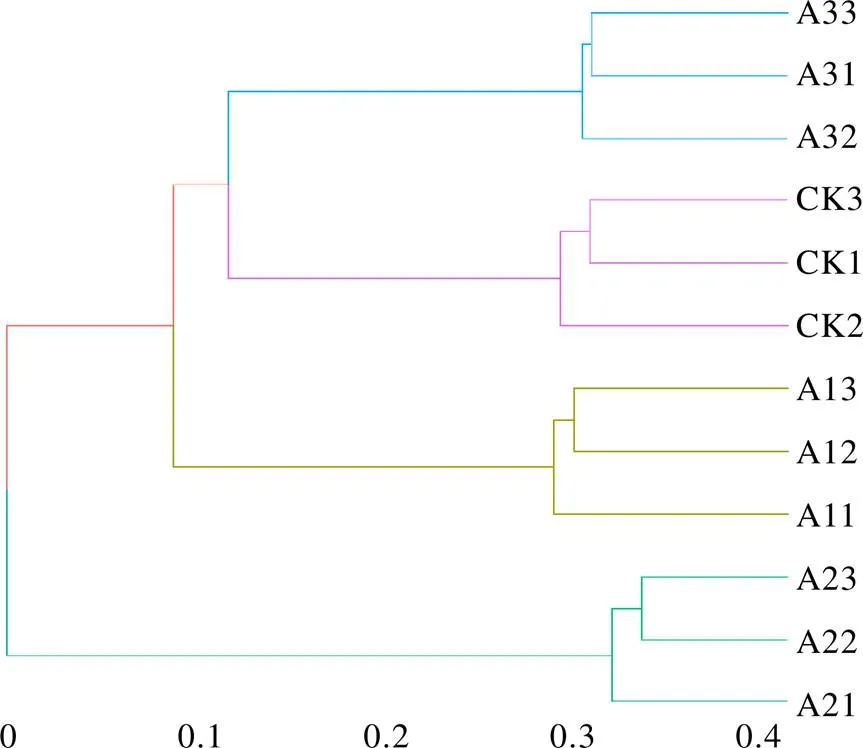
圖3 不同處理茶樹根際土壤真菌群落的層次聚類分析

表3 不同處理茶樹根際土壤真菌優勢菌門相對豐度
注:每行不同字母表示差異達顯著水平(<0.05,n=3)
Note: Different letters in each line indicate significant differences (< 0.05, n=3)
2.4.2 基于屬水平不同處理茶樹根際土壤真菌群落結構
由表4可知,基于屬水平上,未定義菌屬(unidentified)、古根菌屬()、棉革菌屬()、角擔菌屬()和支孢瓶霉屬()是套種靈芝茶樹根際土壤的優勢菌屬(前5)。其中,CK、A1和A3土壤中豐度最高的是未定義菌屬,相對豐度分別為45.02%、29.70%和53.63%;而A2土壤中棉革菌屬豐度最高,相對豐度為55.26%。與CK相比,A1土壤中角擔菌屬、被孢霉屬()、毛孢子菌屬()和三形菌屬()分別顯著上升59.14、1.34、3.70倍和1.92倍(<0.05);A2土壤的古根菌屬顯著下降76.81%,而棉革菌屬和支孢瓶霉屬分別顯著上升788.43倍和36.24倍(<0.05);A3土壤的被孢霉屬和靈芝屬()分別顯著上升1.09倍和0.81倍(<0.05)。此外,熱圖結果也表明,不同處理茶樹根際土壤真菌群落組成結構發生明顯變化(圖4)。
2.4.3不同處理茶樹根際土壤真菌群落差異物種分析
多級物種差異判別分析(Linear discriminant analysis effect size,LEfSe)可實現多個分組之間的比較,還可在分組內部進行亞組比較分析,從而找到組間在豐度上有顯著差異的物種(即生物標志物,Biomaker)(LDA score≥4,<0.05)。由門到屬的分類層級可知,部分真菌群落類群富集特征顯著。如子囊菌門、被孢霉門和擔子菌門分別可作為CK、A1和A2根際土壤真菌群落的生物標志物,而羅茲菌門(Rozellomycota)和未定義菌門在A3土壤中豐度最高(圖5)。在CK、A1、A2和A3土壤中分別有10個(1門2綱3目4科)、20個(1門4綱8目7科)、6個(1門1綱2目2科)、16個(2門3綱6目5科)具有統計學差異的生物標志物。
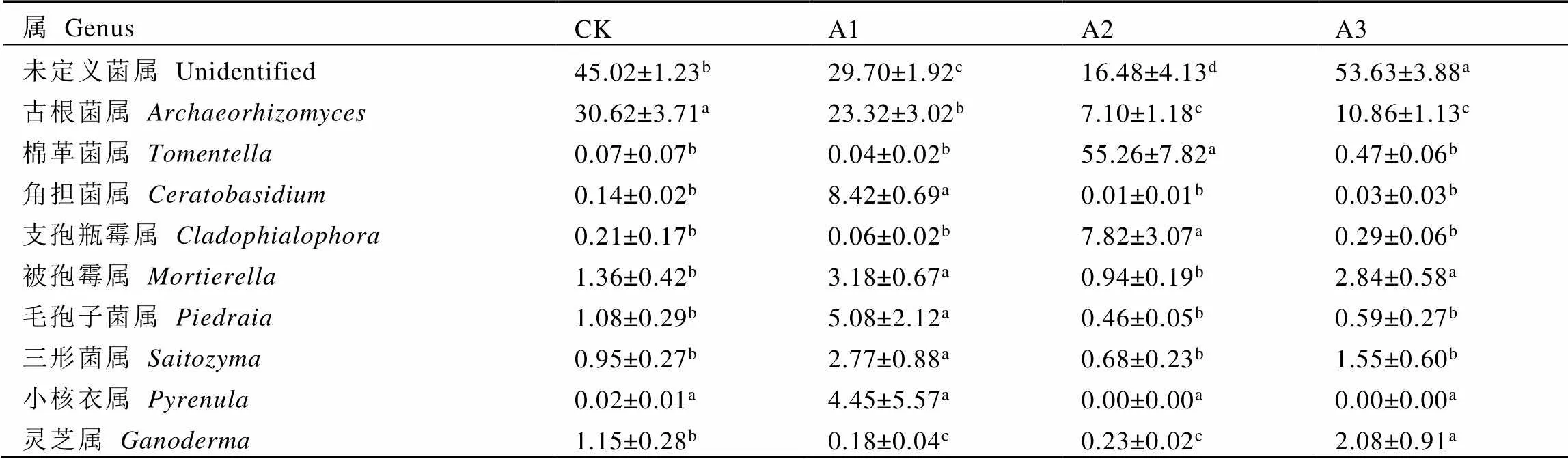
表4 不同處理茶樹根際土壤優勢真菌相對豐度(屬水平)
注:每行不同字母表示差異達顯著水平(<0.05,n=3)
Note: Different letters in each line indicate significant differences (< 0.05, n=3)

圖4 不同處理茶樹根際土壤真菌群落熱圖分析(屬水平)
3 討論
3.1 套種靈芝提高茶樹根際土壤肥力
茶樹根際土壤理化性質結果表明,套種靈芝的栽培模式提高了茶樹根際土壤pH值,即茶園套種靈芝在一定程度上可能會緩解土壤酸化進程;茶樹根際土壤堿解氮、速效磷、速效鉀及有機碳含量在套種靈芝后顯著提高(表1),研究結果與蔣玉蘭等[11]和李振武等[14]通過茶園間套作食用菌的結果相似。這可能是因為食用菌采收后的菌棒中仍含有大量菌絲殘體,可為土壤提供蛋白質、氨基酸和礦物質等營養成分,進而促進茶樹根系吸收營養物質。可見,茶園套種靈芝的生態栽植模式在平衡土壤酸堿度、改善土壤肥力方面具有調節作用。
3.2 套種靈芝改變茶樹根際土壤真菌群落多樣性
根際土壤微生物多樣性是植物生長和土壤健康的重要驅動力,與植物生長、養分循環、病害抑制、污染物降解等土壤生態功能息息相關[15],豐富度和均勻度較高的微生物群落擁有更廣泛的生活史策略,可以提高土壤生態系統應對外界負面干擾的抵抗力和恢復力[16]。前人研究表明,合理的間套作可顯著提高根際土壤微生物群落多樣性及穩定性,改善植物根際微環境進而提高作物產量及品質[17-18]。本研究中,由不同處理茶樹根際土壤真菌群落多樣性指數可知,套種靈芝后A1和A2的茶樹根際土壤真菌群落多樣性指數顯著降低,而A3土壤中真菌群落多樣性指數與CK相比無顯著性差異(表2)。傅海平等[19]探究綠肥間作對茶樹根際土壤真菌群落多樣性的影響,其結果與本研究相似。研究表明,栽植于地表的植被是驅動根際土壤真菌群落多樣性變化的主要因素[20]。因此,我們推測茶樹套種靈芝后,其根際微域可能形成了“茶—靈芝—土壤”緊密互作的共生網絡,進而影響土壤真菌群落結構多樣性。此外,本研究采用UPGMA方法對樣本進行層次聚類分析,通過樹枝結構描述和比較多個樣本間的相似性與差異性,分枝長度越短,樣本相似度越高[21]。結果表明,套種靈芝的A2茶樹根際土壤與CK、A1、A3的真菌群落組成結構差異相對較大(圖2和圖3)。大量研究證實,根際土壤特性是影響土壤真菌群落結構多樣性的關鍵因素,尤其是土壤pH值[22-23],推測套種靈芝后茶樹根際土壤真菌群落結構多樣性的差異可能受土壤pH值影響較大。
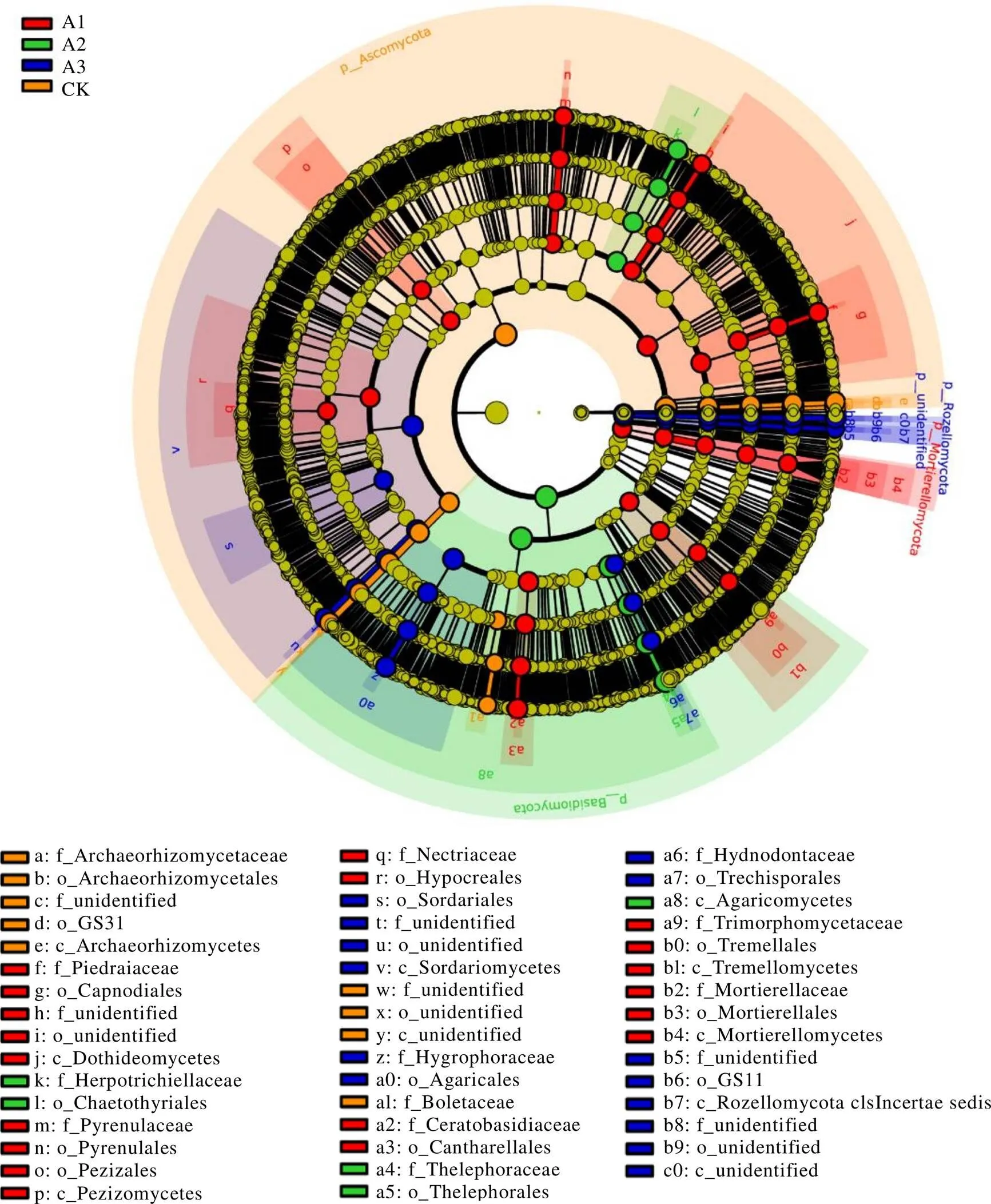
圖5 不同處理茶樹根際土壤真菌群落LEfSe生物標志物
3.3 套種靈芝改善茶樹根際土壤真菌群落結構
根際微生物是土壤生態系統能量流動、物質循環和信息傳遞的主要驅動者,對維持土壤生產力至關重要[24]。相較于細菌,真菌具有更強的吸收營養物質和降解復雜有機物的能力[25]。研究表明,目前已知98%的土壤生態系統真菌均屬于子囊菌門和擔子菌門,且屬于子囊菌門的真菌數量大于擔子菌門[26]。本研究中,土壤真菌門水平分布特征為常見類群比例較高,其中子囊菌門和擔子菌門的相對豐度之和達81.54%~93.49%,其余門類僅占6.51%~18.46%。擔子菌門大多為腐生菌,可以分解木質素、纖維素等大分子化合物[27]。套種靈芝顯著提高了擔子菌門的相對豐度。靈芝子實體采收后,將其菌棒覆土歸埋的栽培模式為茶樹根際土壤提供大量營養源,進而為擔子菌門菌群定殖及發展提供了較好的營養物質基礎。基于屬水平而言,套種靈芝顯著提高茶樹根際土壤角擔菌屬、被孢霉屬、毛孢子菌屬、三形菌屬、棉革菌屬、支孢瓶霉屬和靈芝屬的相對豐度,而古根菌屬的相對豐度顯著降低。有研究表明,被孢霉屬具有溶磷性[28]、分解纖維素及木質素[29]、提高土壤酶活性[30]、增加土壤養分含量[31]的潛力。棉革菌屬是常見的可與共生植物根系形成外生菌根(Ectomycorrhiza,ECM)的類群,菌根共生體具有提高宿主植物對土壤礦質營養元素的吸收和累積、增強植物抗逆性等特征[32]。可見,靈芝采收后其菌棒覆土回歸茶園的栽培模式可富集有益菌加速菌棒降解或形成菌根共生體,有利于茶樹土壤養分均衡供給,促進植株生長發育。大量研究證實,套種模式必然會改變土壤溫度、光照、水分、孔隙度、養分等一系列土壤環境因子,植物根系分泌物和組織脫落物、凋落物等也會隨之改變,對根際微生物的代謝和生長發育產生直接或間接影響,進而對根際微生物種類、數量和分布產生影響[33-34]。因此,推測套種靈芝后茶樹根際產生的生態效應對土壤真菌具有一定選擇性作用,最終導致茶樹真菌群落組成結構產生差異。然而,根際土壤環境因子與微生物的互作對影響土壤微生物群落結構是否具有協同效應仍需進一步研究。
本研究揭示了茶樹根際土壤真菌群落多樣性及組成結構對靈芝不同套種年限的響應,為恢復茶園生態系統多樣性及根際土壤環境質量提供綠色途徑。
[1] 李艷姣, 林春桃, 范水生, 等. 福建茶產業發展與供給側結構性改革互動協調研究[J]. 農業展望, 2021, 17(9): 108-114.Li Y J, Lin C T, Fan S S, et al. Study on the interaction and coordination between tea industry development and supply-side structural reform in Fujian [J]. Agricultural Outlook, 2021, 17(9): 108-114.
[2] 廖萬有, 王宏樹, 蘇有鍵, 等. 我國茶園土壤的退化問題及其防治[C]//中國茶葉學會. 茶葉科技創新與產業發展學術研討會論文集, 重慶: [出版者不詳], 2009: 185-193. Liao W Y, Wang H S, Su Y J, et al. Soil degeneration and its prevention in Chinese tea gardens [C]//China Tea Science Society. Proceedings of the symposium on technological innovation and industrial development of tea. Chongqing: [s.n.], 2009: 185-193.
[3] 史凡, 黃泓晶, 陳燕婷, 等. 間套作功能植物對茶園生態系統服務功能的影響[J]. 茶葉科學, 2022, 42(2): 151-168. Shi F, Huang H J, Chen Y T, et al. Effects of intercropping functional plants on the ecosystem functions and services in tea garden [J]. Journal of Tea Science, 2022, 42(2): 151-168.
[4] 齊龍波, 周衛軍, 郭海彥, 等. 覆蓋和間作對亞熱帶紅壤茶園土壤磷營養的影響[J]. 中國生態農業學報, 2008, 16(3): 593-597. Qi L B, Zhou W J, Guo H Y, et al. Phosphorus nutrient characteristics of tea plantation soils under rice-straw mulch and white-clover intercropping in hilly red-soils of South China [J]. Chinese Journal of Eco-Agriculture, 2008, 16(3): 593-597.
[5] 宋莉, 廖萬有, 王燁軍, 等. 套種綠肥對茶園土壤理化性狀的影響[J]. 土壤, 2016, 48(4): 675-679. Song L, Liao W Y, Wang Y J, et al. Effects of interplanting green manure on soil physico-chemical characters in tea plantation [J]. Soils, 2016, 48(4): 675-679.
[6] 沈程文, 肖潤林, 徐華勤. 覆蓋與間作對亞熱帶丘陵區茶園土壤微生物量的影響[J]. 水土保持學報, 2006, 20(3): 141-144. Shen C W, Xiao R L, Xu H Q.Effects of cover and intercropping on soil microbial biomass of tea plantations in subtropical hilly region [J]. Journal of Soil and Water Conservation, 2006, 20(3): 141-144.
[7] 徐華勤, 肖潤林, 宋同清, 等. 稻草覆蓋與間作三葉草對丘陵茶園土壤微生物群落功能的影響[J]. 生物多樣性, 2008, 16(2): 166-174. Xu H Q, Xiao R L, Song T Q, et al. Effects of mulching and intercropping on the functional diversity of soil microbial communities in tea plantations [J]. Biodiversity Science, 2008, 16(2): 166-174.
[8] 林黎. 草種組合套種對山地茶園土壤性狀及茶葉品質的影響[J]. 茶葉學報, 2017, 58(4): 174-178. Lin L. Physiochemical properties of soil and quality of tea affected by combination inter-cropping in hilly tea plantation [J]. Acta Tea Sinica, 2017, 58(4): 174-178.
[9] Li Y C, Li Z, Li Z W, et al. Variations of rhizosphere bacterial communities in tea (L.) continuous cropping soil by high-throughput pyrosequencing approach [J]. Journal of Applied Microbiology, 2016, 121(3): 787-799.
[10] 黃偉. 淺論食用菌返生態野生栽培[J]. 中國食用菌, 2008, 27(5): 33-34. Huang W. Superficial view on ecological wild cultivation of edible fungi [J].Edible Fungi of China, 2008, 27(5): 33-34.
[11] 蔣玉蘭, 張海華, 潘俊嫻, 等. 茶樹和長根菇間作試驗研究[J]. 中國食用菌, 2018, 37(6): 32-35, 39. Jiang Y L, Zhang H H, Pan J X, et al. Experiment study on intercropping of tea trees and[J]. Edible Fungi of China, 2018, 37(6): 32-35, 39.
[12] 李艷春, 林忠寧, 陸烝, 等. 茶園間作靈芝對土壤細菌多樣性和群落結構的影響[J]. 福建農業學報, 2019, 34(6): 690-696.Li Y C, Lin Z N, Lu Z, et al. Microbial diversity and structure in soil under tea bushesintercropping [J]. Fujian Journal of Agricultural Sciences, 2019, 34(6): 690-696.
[13] 嚴麗君, 王普, 施啟龍, 等. 動物食性分析在生態學中的應用研究進展——基于DNA宏條形碼技術[J]. 生態學報, 2023, 43(8): 3007-3019. Yan L J, Wang P, Shi Q L, et al.Applications of animal diet analysis based on DNA metabarcoding in ecological research [J]. Acta Ecologica Sinica, 2023, 43(8): 3007-3019.
[14] 李振武, 韓海東, 陳敏健, 等. 套種食用菌對茶園土壤和茶樹生長的效應[J]. 福建農業學報, 2013, 28(11): 1088-1092. Li Z W, Han H D, Chen M J, et al. Effects of intercroppingon tea garden soil and tea growth [J]. Fujian Journal of Agricultural Sciences, 2013, 28(11): 1088-1092.
[15] Labouyrie M, Ballabio C, Romero F, et al. Patterns in soil microbial diversity across Europe [J]. Nature Communications, 2023, 14(1): 3311. doi: 10.1038/s41467-023-37937-4.
[16] Saleem M, Hu J, Jousset A. More than the sum of its parts: microbiome biodiversity as a driver of plant growth and soil health [J]. Annual Review of Ecology, Evolution, and Systematics, 2019, 50(1): 145-168.
[17] 趙佳佳, 李季, 杜相革. 不同生產模式對土壤微生物種群數量的影響[J]. 華北農學報, 2011, 26(s1): 220-224. Zhao J J, Li J, Du X G. Dynamics of soil microbial community in the experiment of organic, green and conventional vegetable production systems [J]. Acta Agriculturae Boreali-Sinica, 2011, 26(s1): 220-224.
[18] 林雁冰, 薛泉宏, 顏霞. 不同栽培模式下玉米根系對土壤微生物區系的影響[J]. 西北農林科技大學學報(自然科學版), 2008, 36(12): 101-107. Lin Y B, Xue Q H, Yan X. Effects of the maize root on soil microbial flora under different cultivation patterns [J]. Journal of Northwest A&F University (Natural Science Edition), 2008, 36(12): 101-107.
[19] 傅海平, 周品謙, 王沅江, 等. 綠肥間作對茶樹根際土壤真菌群落的影響[J]. 茶葉通訊, 2020, 47(3): 406-415. Fu H P, Zhou P Q, Wang Y J, et al. Effects of intercropping different green manures on fungal community characteristics in rhizosphere soil of tea plant [J]. Journal of Tea Communication, 2020, 47(3): 406-415.
[20] Christopher W S, Anna R. Comment on “global diversity and geography of soil fungi” [J]. Science, 2015, 348(6242): 1438-1438.
[21] Wei Z, Yu D. Analysis of the succession of structure of the bacteria community in soil from long-term continuous cotton cropping in Xinjiang using high-throughput sequencing [J]. Archives of Microbiology, 2018, 200(4): 653-662.
[22] Zhang T, Wang N F, Liu H Y, et al. Soil pH is a key determinant of soil fungal community composition in the Ny-?lesund region, Svalbard (High Arctic) [J].Frontiers in Microbiology, 2016, 7: 227. doi: 10.3389/fmicb.2016.00227.
[23] Lauber C L, Strickland M S, Bradford M A, et al. The influence of soil properties on the structure of bacterial and fungal communities across land-use types [J]. Soil Biology and Biochemistry, 2008, 40(9): 2407-2415.
[24] Prakash J, Mishra S. Role of beneficial soil microbes in alleviating climatic stresses in plants [M]//Kumar A, Singh J, Ferreira L F R. Microbiome under changing climate: implications and solutions. Cambridge:Woodhead Publishing, 2022: 29-68.
[25] Peroh D. Plant-associated fungal communities in the light of meta'omics [J]. Fungal Diversity, 2015, 75: 1-25.
[26] Wang H Y, Guo S Y, Huang M R, et al. Ascomycota has a faster evolutionary rate and higher species diversity than Basidiomycota [J]. Science China life Sciences, 2010, 53(10): 1163-1169.
[27] Lynch M D J, Thorn R G. Diversity of Basidiomycetes in Michigan agricultural soils [J].Applied and Environmental Microbiology, 2006, 72(11): 7050-7056.
[28] Osorio N W, Habte M. Soil phosphate desorption induced by a phosphate-solubilizing fungus [J]. Communications in Soil Science & Plant Analysis, 2014, 45(4): 451-460.
[29] 陳力力, 劉金, 李夢丹, 等. 水稻-油菜雙序列復種免耕、翻耕土壤真菌多樣性[J].激光生物學報, 2018, 27(1): 60-68, 59. Chen L L, Liu J, Li M D, et al.Diversity of filamentous fungus community in paddy fields with different tillage methods [J]. Acta Laser Biology Sinca, 2018, 27(1): 60-68, 59.
[30] Zhang H, Wu X, Li G, et al. Interactions between arbuscular mycorrhizal fungi and phosphate-solubilizing fungus (sp.) and their effects ongrowth and enzyme activities of rhizosphere and bulk soils at different salinities [J]. Biology and Fertility of Soils, 2011, 47(5): 543-554.
[31] Li F, Chen L, Redmile-Gordon M, et al.roles in organic agriculture and crop growth promotion in a mineral soil [J]. Land Degradation and Development, 2018, 29(6): 1642-1651.
[32] 林雙雙, 孫向偉, 王曉娟, 等. 我國菌根學研究進展及其應用展望[J]. 草業學報, 2013, 22(5): 310-325. Lin S S, Sun X W, Wang X J, et al. Mycorrhizal studies and their application prospects in China [J]. Acta Prataculturae Sinica, 2013, 22(5): 310-325.
[33] Li S M, Fan W, Xu G, et al. Bio-organic fertilizers improvegrowth by remolding the soil microbiome and metabolome [J]. Frontiers in Microbiology, 2023, 14: 1117355. doi: 10.3389/fmicb.2023.1117355.
[34] Zhou X G, Zhang J Y, Rahman M K U, et al. Interspecific plant interaction via root exudates structures the disease suppressiveness of rhizosphere microbiomes [J]. Molecular Plant, 2023, 16(5): 849-864.
The Characteristics of Fungal Community Structure in Tea Rhizosphere Soil Interplanted withBased on High-throughput Sequencing Technology
HAN Haidong, ZHOU Liuting*, HUANG Xiaoyun, YU Chengran, HUANG Xiusheng*
Agricultural Ecology Institute, Fujian Academy of Agricultural Sciences. Fujian Engineering and Technology Research Center for Recycling Agriculture, Fuzhou 350013, China
The tea plantation withis an ecological cycle intercropping model of resource utilization of tea processing waste, and the composite community formed can cover the ground more thoroughly and play an important role in improving the soil microbial community structure and maintaining the balance of soil microbiological system. In this study, we investigated the changes of tea rhizosphere soil fungal community structure in uncropped (CK), intercropping 1 year (A1), intercropping 2 years (A2) and intercropping 3 years (A3) using Miseq PE300 high-throughput sequencing technology. The results show that: (1) compared with CK, interplantingsignificantly increased the contents of available nitrogen, available phosphorus, available potassium and organic carbon in tea rhizosphere soil, with soil of A3 having the highest increase, reaching 32.36%, 13.01%, 69.21% and 9.56%, respectively. (2) Thediversity index shows that the observed species and Chao1 index of tea rhizosphere soil fungal community were CK>A3>A1>A2. ACE index, Shannon index and Simpson index showed A3>CK>A1>A2. (3) Thediversity index shows that the composition and structure of fungal community in tea rhizosphere soil of A2 were relatively different from those of CK, A1 and A3. (4) Through taxonomic analysis, it is found that tea rhizosphere soil fungi were distributed in 18 phyla, 48 classes, 135 orders, 309 families and 632 genera. At phylum level, Ascomycota was the dominant phylum of CK, A1 and A3, with relative abundances of 71.28%, 68.74% and 51.79%, respectively. Basidiomycota was the dominant phylum of A2 with a relative abundance of 64.48%. At the genus level, compared with CK, the contents of,,andin A1 were significantly increased by 59.14, 1.34, 3.70 and 1.92 times, respectively (<0.05). The relative abundance ofin A2 decreased significantly by 76.81%, while that ofandincreased by 788.43 and 36.24 times, respectively (<0.05). Theandin A3 soil significantly increased 1.09 and 0.81 times, respectively (<0.05). In summary, the interplantingin tea gardens can effectively regulate the composition and structure of tea rhizosphere soil fungal community, improve the soil micro-ecological environment and this study provided a theoretical basis for the technical model to promote the sustainable green development of tea plantation.
tea plantation, intercropping, rhizosphere soil, fungal community structure
S571.1;S154.3
A
1000-369X(2023)04-513-12
2023-05-24
2023-07-10
福建省公益類科研院所專項(2020R1021004、2020R1021002、2023R1019002)、福建省人民政府-中國農業科學院“5511”協同創新工程(XTCXGC2021010)、福建省農業科學院項目(CXTD2021009-1)
韓海東,男,副研究員,主要從事生態循環農業與山地茶園有機種植研究。*通信作者:zlt0616@126.com;hxs706@163.com
