河北省大豆孢囊線蟲致病型分化研究
2023-09-02 09:44:12李秀花王容燕高波馬娟高占林趙惟萬艷爭陳書龍
河北農業科學 2023年4期
李秀花,王容燕,高波,馬娟,高占林,趙惟,萬艷爭,陳書龍
(河北省農林科學院植物保護研究所/ 河北省農業有害生物綜合防治工程技術研究中心/ 農業部華北北部作物有害生物綜合治理重點實驗室,河北 保定 071000)
大豆孢囊線蟲(soybean cyst nematode,SCN;Heterodera glycines,Ichinohe) 屬于一種土傳的定居性內寄生線蟲,分布遍及世界主要大豆生產國[1],嚴重威脅著大豆的安全生產。SCN 引起的大豆孢囊線蟲病是世界上為害大豆嚴重的病害之一,在美國每年造成的經濟損失超過10 億美元[2]。目前,SCN 在我國黑龍江、吉林、遼寧、山東、江蘇、河北等22 個省(直轄市、自治區)均有發生[3],每年造成的大豆受害面積超過150 萬hm2,嚴重發病地塊大豆減產30%以上,甚至絕收[4]。
SCN 是專性寄生物,具有明顯的遺傳多樣性。1970 年Golden 等提出用“生理小種”來表示SCN 的種內致病性變異,并且用Pickett、Peking、PI88788 和PI90763 這4 個大豆品種作為SCN 生理分化的鑒別寄主,用感病大豆品種Lee 68 作為標準對照[5]。1988 年美國線蟲學家Riggs 等利用SCN 在4 個鑒別寄主上的可能反應,理論上把SCN 劃分成16 個生理小種[6],稱之為Race 鑒定體系。但由于該鑒定體系包含的抗病基因較少,并不能完全把攜帶不同致病基因的SCN 群體區分開來,因此,Niblack 等在2002年又建立了一個新的SCN 基因多樣性分類系統——HG 型分類鑒定體系[7],以感病寄主對應的寄主序號來命名HG 型的類型。截至目前,采用Race 鑒定體系我國已報道有1、2、3、4、5、6、7、9、13、14號共計10個生理小種[8~18]。通過HG 鑒定體系已發現12 個HG 型,分別是0、7、2.7、5.7、1.3.7、2.5.7、1.2.5.7、1.2.3.5.7、1.2.3.4.5.6.7、1.2.3.5.6.7、1.3 和2.5.7[19~21]。使用這2 種鑒定體系對SCN 群體進行鑒定[20],得到的結果并不完全對應。HG 型鑒定體系更能體現出SCN 不同毒力表型的多樣性,但使用鑒別寄主過多,且鑒定所需的孢囊線蟲群體數量遠遠多于Race 鑒定體系,因而,對于數量較少的SCN群體就無法進行有效的HG 型鑒定。
20 世紀90 年代河北省大豆種植面積最高達到52萬hm2左右,之后逐年下滑。自2018 年國家實施大豆振興計劃以來,河北省大豆種植面積又開始出現恢復性增長,2022 年純大豆種植面積達9 萬hm2、大豆/玉米帶狀復合種植新模式面積約7 萬hm2,大豆孢囊線蟲病呈嚴重上升趨勢。種植抗病品種是防治大豆孢囊線蟲病最經濟有效的措施,而深入了解SCN 的生理小種或HG 型是開展大豆抗孢囊線蟲品種選育以及品種合理布局利用的重要前提。早在2006 年,河北省SCN群體已報道有1、2、5 號生理小種,無HG 型鑒定報道。SCN 的致病型變異與品種的更迭密切相關。截至目前,大豆種植面積銳減后的迅速擴張以及感病品種的大面積種植是否會引起河北省SCN 群體的毒性變異尚不明確。基于此,在河北省近幾年大豆種植面積大且SCN發生重的地市采集土樣,對其SCN 群體均采用Race和HG 型2 種鑒定體系進行鑒定,旨為大豆抗孢囊線蟲品種選育以及大豆孢囊線蟲的有效防治提供理論依據。
1 材料與方法
1.1 試驗材料
大豆孢囊線蟲致病型鑒別寄主共9 個,分別為Lee、Peking(PI548402)、PI88788、PI90763、Pickett、PI437654、PI209332、PI89772 和PI548316,其中Lee為感病對照。均由沈陽農業大學惠贈,并在河北省農林科學院植物保護研究所隔離繁育后使用。
1.2 試驗方法
1.2.1 大豆孢囊線蟲病土采集 2020 年10 月大豆收獲后,在河北省大豆主產區的邯鄲市磁縣、保定市安國市北揚城鄉、唐山市豐臺區岳龍鎮、滄州市滄縣周莊子和承德市灤平區平坊村大豆孢囊線蟲病重病田,用軍用鐵鏟在根圍取土樣,裝入塑料袋,置于室外陰涼處。翌年4 月使用。
1.2.2 大豆鑒別寄主種子催芽 將9 個大豆鑒別寄主的種子,先用質量分數0.5%的NaClO 溶液表面消毒3 min,而后用無菌水沖洗干凈,置于25 ℃左右的溫水中浸泡6~7 h。將浸泡好的大豆種子置于內墊2 層滅菌濾紙的培養皿內,向濾紙上滴加無菌水2 mL,置于25 ℃恒溫培養箱中催芽。經過3~4 d,待種子幼苗根長2 cm、芽長1 cm 左右時播入花盆。
1.2.3 大豆孢囊線蟲病土配制 2021 年4 月下旬將采集的大豆孢囊線蟲病土過篩,混合均勻后取200 mL,采用漂浮法[16]分離土樣中的孢囊,統計飽滿孢囊的數量。重復3 次取樣。
為了減少試驗誤差,使各鑒別土壤中含有的線蟲數量差別不是太大,對含有孢囊數量較多的土壤摻入一定量的滅菌細沙土(土、細沙體積比為1∶3),將各地區病土中的飽滿孢囊數量調整至300~600 個/L(土樣)。再次分離混合土樣中的孢囊,確定孢囊數量。
1.2.4 大豆栽種與管理 用燒杯量取混合好的病土500mL 裝入花盆(上口直徑11cm、下口直徑10.5cm),栽種1 株已催芽的鑒別寄主,覆土后澆水使土壤濕透,置于陽光溫室(溫度18~28 ℃)內,正常管理。每處理重復8 盆。
1.2.5 大豆病害調查 大豆生長42 d 后,在雌蟲集中出現時進行調查。首先統計根系上的雌蟲數量;然后利用漂浮法分離土壤中的孢囊[16],在體式顯微鏡下統計孢囊數量。將根系上的雌蟲數量與土壤中的孢囊數量之和,計為大豆孢囊線蟲在大豆播種后42 d 內為害大豆并在大豆根系上所形成的雌蟲總數。
1.2.6 致病性鑒定 主要依據孢囊線蟲在不同鑒別寄主上的繁殖能力,即每個鑒別寄主的平均雌蟲指數(female index,FI),對大豆孢囊線蟲的生理小種類型[6]和HG 型[7]進行鑒定。FI計算公式為:
FI=鑒別品種每株根系上的平均雌蟲數/感病對照品種每株根系上的平均雌蟲數×100
其中,FI≥10 時為感病;FI<10 時為抗病。
2 結果與分析
2.1 不同地區土壤中的孢囊數量
測定結果(表1)顯示,5 個地區根圍土樣中含有的大豆孢囊線蟲孢囊數量為340~2 763 個/L,均滿足盆栽試驗鑒別所需的發病條件。其中,滄州市土壤中的孢囊數量最高,承德市土壤中的孢囊數量最低,土壤中含有孢囊數量最多量是最少量的8 倍以上。
不同地區原有土樣中的孢囊數量差異較大,為了減少試驗誤差,對含有孢囊數量較多的土壤摻入一定量的滅菌細沙土進行調整。調整后各鑒別土壤中的孢囊數量為340~603 個/L。
2.2 大豆孢囊線蟲群體的生理小種鑒定
鑒定結果(表2) 顯示,感病品種Lee 對5 個SCN 群體均表現為高度感病,平均雌蟲數為195.9~295.5 個/株;5 個SCN 群體在4 個鑒別寄主上形成的雌蟲數均較少,FI均<10,鑒別寄主均表現為抗病,參照大豆孢囊線蟲Race 鑒定體系生理小種劃分標準[4],滄州滄縣群體、唐山豐臺區群體、保定安國市群體、邯鄲磁縣群體和承德灤平區群體的生理小種均屬于3號。
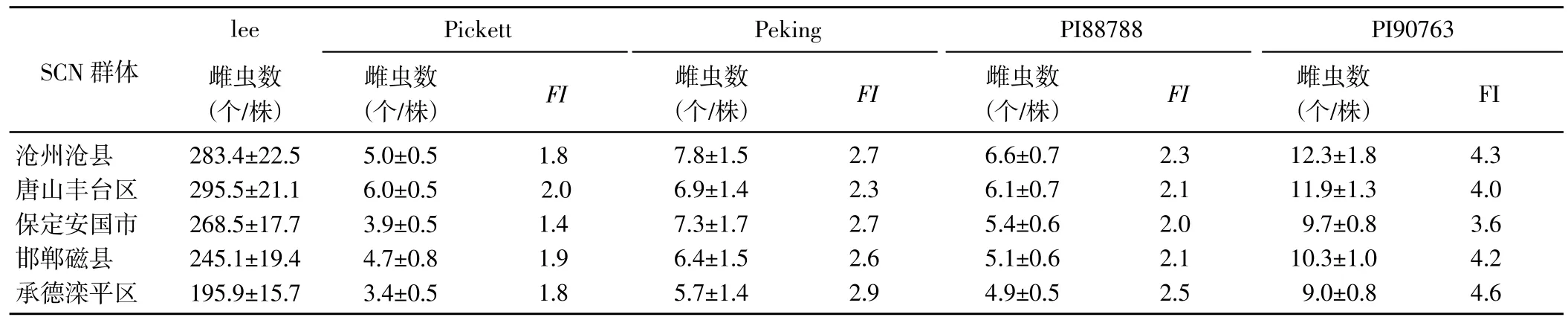
表2 河北省5 個市大豆孢囊線蟲群體的生理小種鑒定結果Table 2 Identification of races of 5 SCN populations in Hebei Province
2.3 大豆孢囊線蟲群體的HG 型鑒定
鑒定結果(表3) 顯示,感病品種Lee 對5 個SCN 群體均表現為高度感病,平均雌蟲數為181.9~275.3 個/株;5 個SCN 群體在7 個鑒別寄主上形成的雌蟲數量均較少,FI均<10,鑒別寄主反應均表現為抗病,參照Niblack 的HG 型鑒定體系劃分標準[5],滄州滄縣群體、唐山豐臺區群體、保定安國市區群體、邯鄲磁縣群體和承德灤平區群體的HG 型均屬于0 型。
3 結論與討論
3.1 討論
大豆孢囊線蟲不同群體間具有明顯的寄生性分化,目前有生理小種[5,6]和HG 型[7]2 套鑒定體系。根據生理小種鑒定體系,截至目前,世界上報道SCN 生理小種類別最多的國家是美國,已報道有1、2、3、4、5、6、7、8、9、10、11、13、14、15 號共計14 個生理小種,其中3 號和6 號為其優勢小種[22]。我國已報道有1、2、3、4、5、6、7、9、13、14 號共計10 個生理小種,其中3 號小種分布于9 個省、4 號小種分布于8 個省、1 號小種分布于5 個省。此前河北省已報道有1、2、5 號生理小種[18]存在,分別分布在邯鄲、藁城和滄縣。本研究在邯鄲市磁縣、滄州市滄縣、保定市安國市、唐山市豐臺區和承德市灤平區又發現了3 號小種,至此,河北省已有1、2、3、5 號4 個生理小種的存在。根據HG 型鑒定體系,目前在我國已報道SCN 有12 個HG 型,分別為0、7、2.7、5.7、1.3.7、2.5.7、 1.2.5.7、 1.2.3.5.7、 1.2.3.4.5.6.7、 1.2.3.5.6.7、1.3、2.5.7[19~21]。本項目5 個SCN 群體的HG 型均鑒定為0 型。不同省份或者同一省份不同區域存在的生理小種或HG 型類型不同,可能與當地的氣候條件、種植方式和栽培品種不同有關。
同一田塊發生的SCN 群體通常是幾個生理小種的混合體,很少存在單一的生理小種,通過鑒別寄主反應鑒別出來的只是某個占優勢的生理小種[11]。在連年種植抗性寄主的選擇壓力下,SCN 生理小種的變異也常有發生[23],如在黑龍江大慶市和安達市大豆孢囊線蟲重病地連續種植抗病品種,導致這兩地生理小種發生變異[11,24],黑龍江省大慶市3 號小種分化為1 號或14 號小種[24,25],安達市3 號小種分化為4 號、6 號或14 號小種[26,27]。本研究結果表明,河北省的SCN 出現了毒性最弱的3 號小種。而以前滄縣是5 號小種,邯鄲市是1 號小種[18]。探究原因,可能是本研究樣品采集地點與原有采集地點不同;也有可能是近幾年種植結構的調整,大豆種植規模變小;還有可能是農民選擇種植一些缺乏抗性的農家品種,對SCN 造成的選擇壓力變小而使其毒性減弱。同樣在HG 型鑒定體系中,同一地塊SCN 群體通常存在幾個HG 型。如,美國愛荷華州立大學2008 年對同一地塊的4 個不同區域分別進行取樣獲得4 個樣,對混合樣品的HG 型測定結果為1.2.5.6.7,但其中2 個區域為1.2.5.7 HG 型,另2 個區域為1.2.3.5.6.7 HG 型[28]。這也說明,無論采用哪種鑒定體系,得到的鑒定結果均僅是SCN 群體的特征表現,而非某個個體線蟲的特征表現。同時也說明,田間SCN 群體的復雜性[6]。同一塊地SCN 致病型分化是動態的,常隨著作物品種的變化而變化,因此,應加強SCN 致病型分化監測,為抗病品種選育與合理布局提供依據。
3.2 結論
對河北省5 個市的大豆孢囊線蟲群體,通過2 套鑒別寄主,分別測定了每個鑒別寄主在鑒定體系中的FI;根據鑒定體系中每個寄主的FI劃分所屬的生理小種類型或HG 型。結果表明,河北省滄州滄縣群體、唐山豐臺區群體、保定安國市區群體、邯鄲磁縣群體和承德灤平區群體的生理小種均屬于3 號小種,HG型均屬于0 型。
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
現代企業(2015年1期)2015-02-28 18:43:18
新高考·高一物理(2014年1期)2014-09-18 01:26:07
中國火炬(2010年7期)2010-07-25 10:26:09
