基于高通量測序的磷石膏上芒草內部細菌多樣性分析
2023-09-06 06:24:14李劍峰郭欣楊張淑卿倪明秀張煜冕郭金梅
南方農業 2023年11期
關鍵詞:植物
周 煬,李劍峰*,郭欣楊,張淑卿,倪明秀,張煜冕,郭金梅
[1.貴州省高等學校生物資源開發利用重點實驗室,貴州貴陽 550018;2.貴州師范學院喀斯特生境土壤與環境生物修復研究所,貴州貴陽 550018;3.貴州磷化(集團)有限責任公司,貴州貴陽550000]
磷石膏是濕法磷酸生產過程中的副產物,主要成分為二水硫酸鈣,除此之外還有部分重金屬元素、可溶性磷酸鹽及氟化物等雜質[1]。經計算每生產1 t 濕法磷酸,將排放出約5 t 的磷石膏廢渣[2]。目前,國內對于磷石膏利用率不足40%,大部分是通過堆存方式處理,這種處理方式不僅占據了大量的土地資源,而且其潛在的污染物泄露和垮堆風險也會對當地居民的身體和生態環境造成威脅[3]。磷石膏的低成本處理與有效利用始終是我國磷化工業的一大難題。
芒草(Miscanthus Sinensis)為禾本科(Gramineae)芒屬植物[4],通常分布在熱帶、亞熱帶及溫帶亞洲地區。芒草在我國分布較廣,其對于環境的適應能力非常強,在各種惡劣環境下都能夠生存[5]。目前已有將芒草應用于重金屬土壤修復的相關研究報道,并發現芒草能夠耐受一種甚至多種高濃度重金屬元素的脅迫[6]。貴州作為磷石膏生產排放的集中區域,且磷石膏的產出量在逐年增長,對于磷石膏的有效處理已成為當今貴州乃至全國的熱點問題。目前,有關利用芒草植株修復磷石膏堆渣污染的相關研究報道還較為少見,因此具有較大的研究意義。
1 材料與方法
1.1 材料
2021 年7 月,在貴州省貴陽市開陽縣龍景灣貴州磷化集團開陽露天堆場(東經106.852 3°,北緯27.151 3°)采集磷石膏上生長的芒草植株和種子樣品,采集時芒草呈深綠色,群落單一,株高在0.4~1.2 m。
1.2 方法
1.2.1 樣品處理
將植株根系挖出,用無菌水清洗表面后放入陰涼通風處自然風干,待植株風干后與種子一同裝入無菌自封袋內放入4 ℃冰箱暫存。
1.2.2 DNA提取及測序
根據試劑盒要求提取樣本的DNA,利用1%的瓊脂糖凝膠電泳測定DNA 純度。采用16SrDNA 對于V3-V4 引物進行擴增,引物序列為(上游引物338F:ACTCCTACGGGAGGCAGCAG,下游引物 806R:GGACTACHVGGGTWTCTAAT)PCR 反應參數:a.1×(3 min at 95 ℃)b.循環數×30 s at 95 ℃;30 s at 55 ℃;45 s at 72 ℃)c.10 min at 72 ℃,10 ℃until halted by user。根據各樣品的測序量,將其配置成對應的配比,送至生工(上海)生物工程有限公司,利用Illumina 公司的Miseq PE300,對其進行高通量測序。
2 結果與分析
2.1 細菌多樣性分析
2.1.1 測序數據處理結果
如表1所示,樣品經過高通量測序共獲得124 097條原始序列,經對原始數據初步處理,獲得了123 363條有效的序列。供試樣本有效序列條數在51 383~71 980 條,其中phps(芒草種子)有效序列條數大于php(芒草植株)。通過聚類一共獲得274個OTUS,通常將相似性值≥97%的序列定義為同一個OTUS。為了比較直觀地分析和統計,對于測序樣本的共有OTUS和特有OTUS 進行分析比較,結果顯示,php 與phps的共有OTUS 有138 個,php 特有OTUS 為66 個,phps特有OTUS 為70 個,即php 與phps 的微生物群落組成較為相似。

表1 芒草植株及種子有效序列數據統計
2.1.2 α多樣性
樣品稀釋曲線傾斜度在不斷降低逐漸趨于平緩(見圖1),同時樣品中測序覆蓋度均為1(見表2),說明測序數據量比較合理,基本上能夠反映出樣品細菌群落的組成情況[7]。通過計算Ace指數、Chao指數、Shannon指數等三個指標結果表明,php細菌多樣性大于phps。
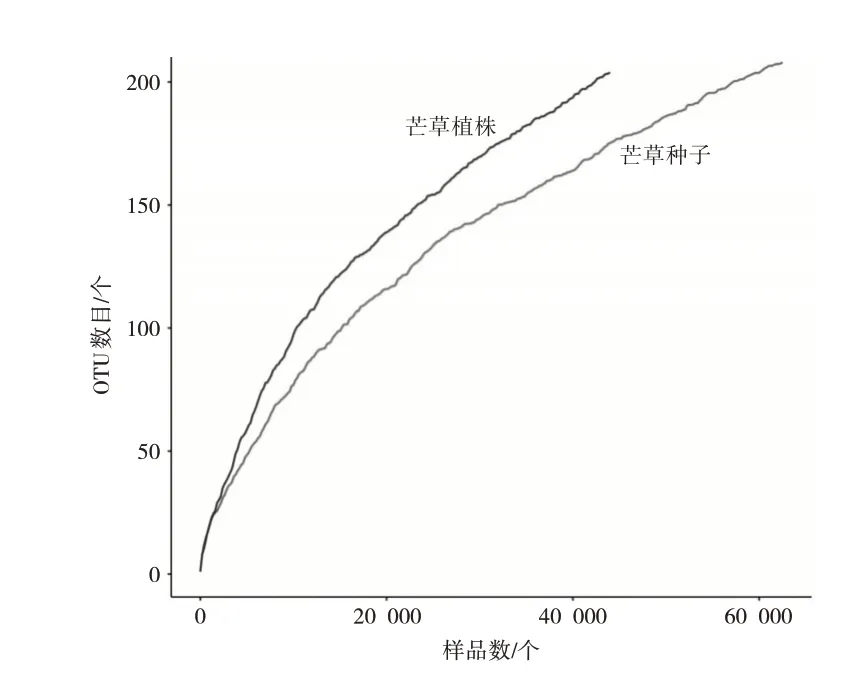
圖1 基于OTUS豐度的樣品微生物群落稀釋曲線

表2 芒草植株及種子的細菌α多樣性
Rank Abundance 曲線圖能夠直觀明確地展示各樣本中OTUS 的均勻度及豐富度,如圖2 所示,當OTU數量小于100時,php與phps的物種相對豐度較高,但曲線下降速度比較快,說明php 與phps 的分布均勻程度較低。當OTU 數量超過100 的情況下,物種的相對豐度較低,但是曲線比較平坦,表明物種分布較為均衡。圖中php 與phps 樣本間的曲線寬度、平滑程度都比較一致,表明其物種豐度及均勻度都無顯著差異。
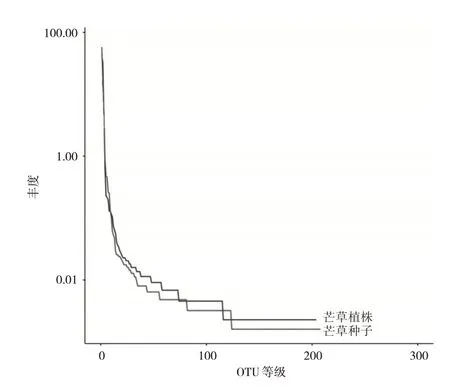
圖2 基于OTUS水平Rank Abundance曲線分析
2.1.3 樣本主成分
通過主成分(PCA)分析可知,主成分1(PC1)和主成分2(PC2)貢獻率分別為58.23% 和40.5%,合計為98.73%。如圖3 所示,兩樣本點的距離十分接近,且位于第一象限有明顯的聚集,表明php 與phps 的細菌相似性較高。
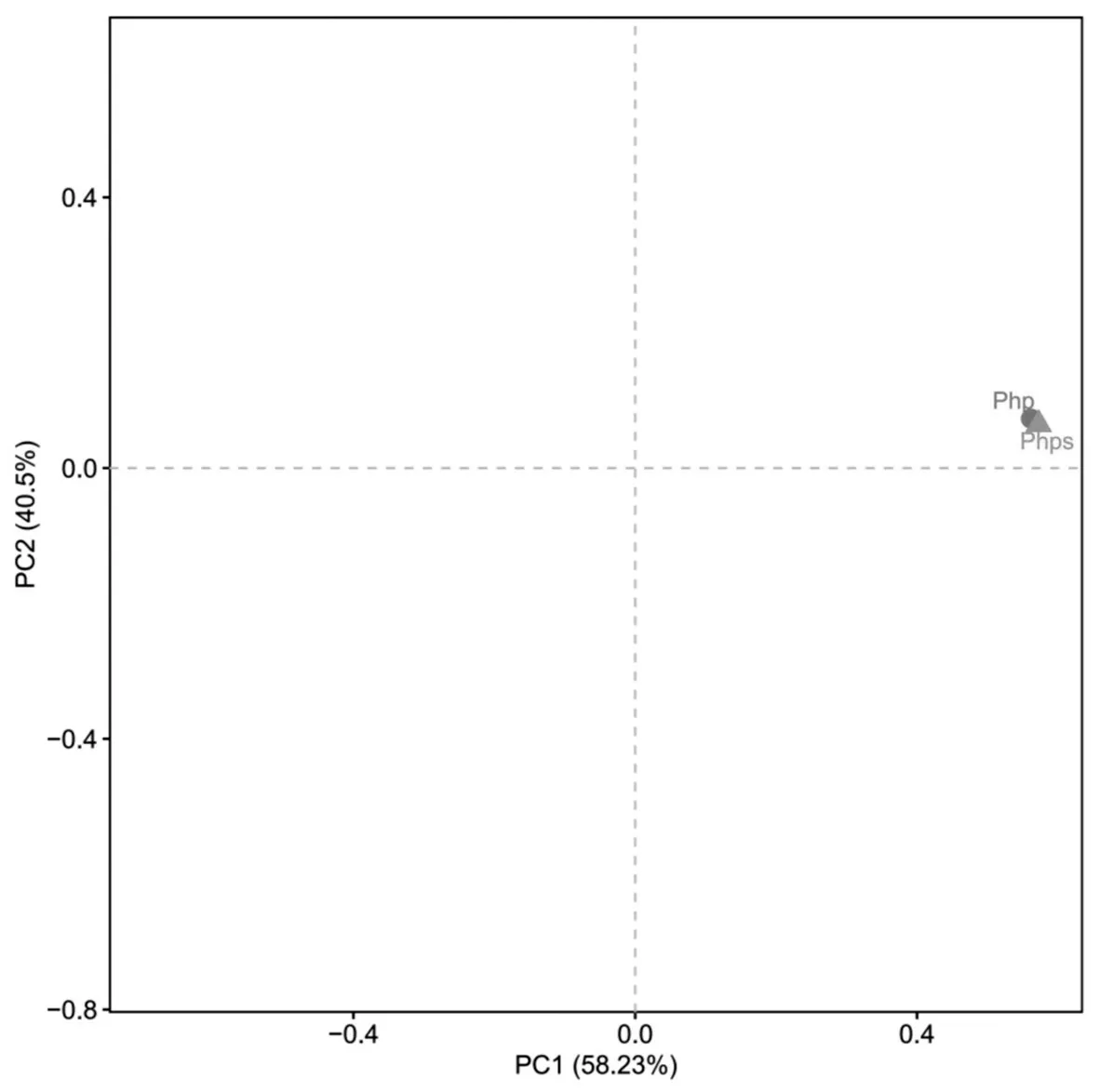
圖3 芒草植株和種子OTU水平PCA圖
2.2 細菌群落結構差異分析
2.2.1 門水平下陳化磷石膏上生長芒草植株及種子細菌群落結構
在門水平下,一共鑒定出17 個門類細菌,如表3所示,豐度低于1%的細菌門合并為Others,其中,芒草植株(php)及種子(phps)共有的優勢細菌門為藍菌門(Cyanobacteria_Chloroplast)。如圖4 所示,藍菌門細菌在芒草植株、種子內部的相對豐度分別為59%、79.3%,即藍菌門的相對豐度phps大于php。原因可能是:1)種子相對于植株而言,營養物質積累較多,較多的營養物質有利于藍菌門的生長,所以種子內部的藍菌門豐度較大;2)在細菌功能方面,藍菌門的菌類具有較強的抗紫外線輻射能力,在較強紫外線的輻射下能夠形成一種代謝響應機制,使其適應紫外輻射的脅迫[8]。
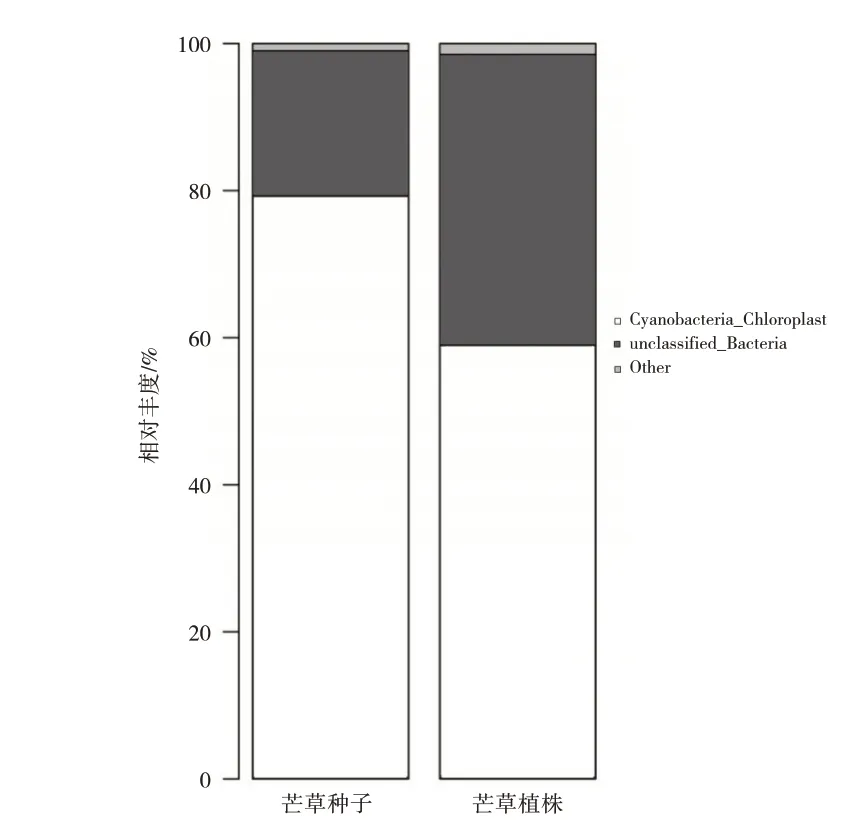
圖4 門水平下芒草植株及種子細菌相對豐度圖
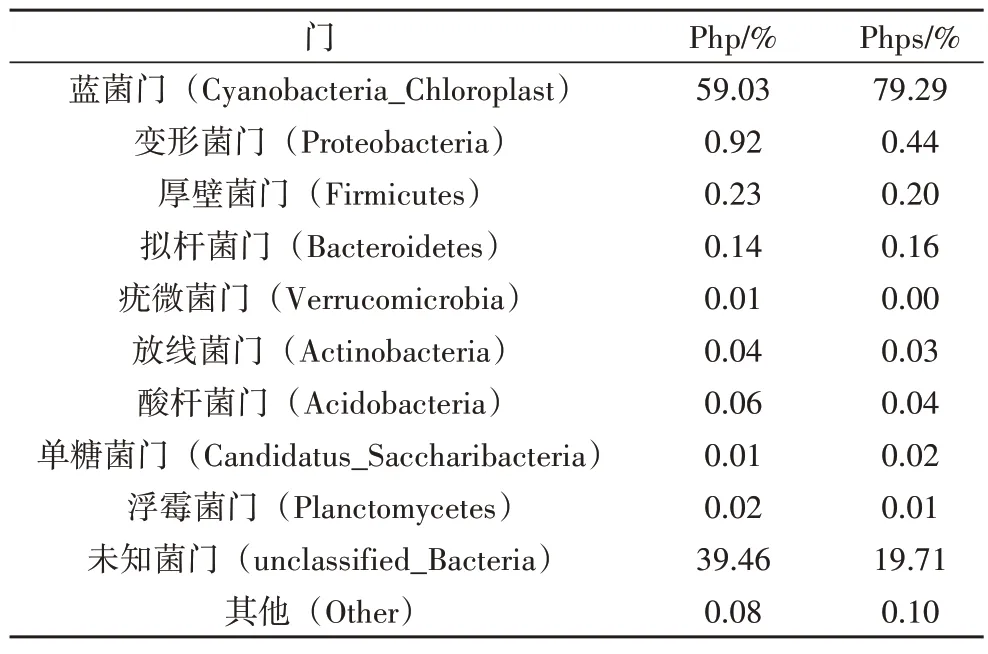
表3 門水平下芒草植株及種子細菌相對豐度表
2.2.2 屬水平下陳化磷石膏上生長芒草植株及種子細菌相對豐度
在屬水平下,共檢測出148 個屬的細菌,在芒草植株中鏈型植物(Streptophyta 59%)占比最大,其次是未知菌屬細菌(unclassified_Bacteria 39.5%);鞘氨醇單胞菌屬(Sphingomonas 0.07%)為植株菌群內部的重要細菌群落,在芒草種子(phps)中占比最大的同樣為鏈型植物(Streptophyta 79.2%),其次為未知菌屬(unclassified_Bacteria 19.8%)。如表4、圖5 所示,在植株到種子過程中,其內部鏈型植物、未知菌豐度逐漸增大,但鞘氨醇單胞菌屬、鏈霉菌屬等細菌豐度相對減少。鏈型植物占比較高的原因可能為芒草植株及種子內部葉綠體信息沒有被排除,導致測序結果中鏈型植物數據占了絕大部分。芒草植株之所以能夠在磷石膏上生長,原因可能是鞘氨醇單胞菌屬、鏈霉菌屬細菌對植物的生長也起著一定的促進作用。鞘氨醇單胞菌屬細菌常被研究者用于修復重金屬污染土壤[9]。
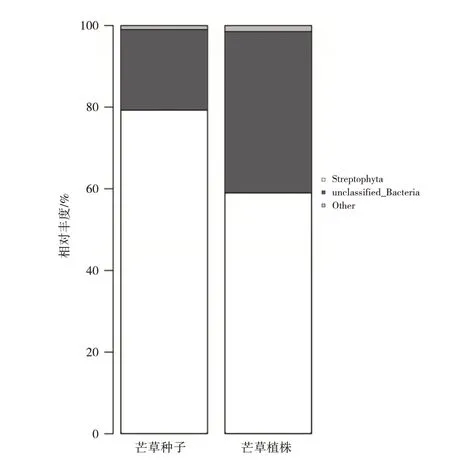
圖5 屬水平下芒草植株及種子細菌相對豐度圖
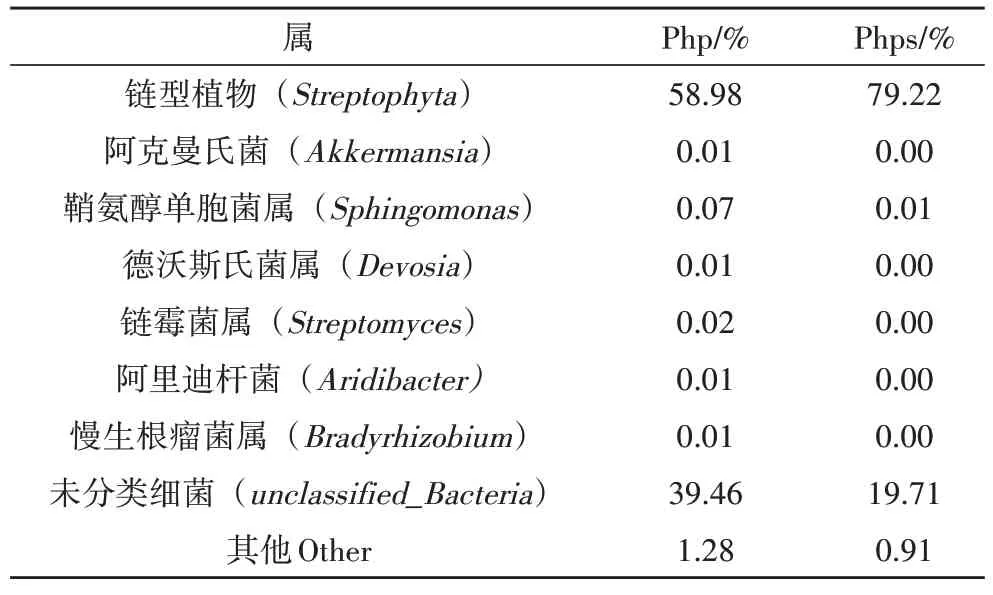
表4 屬水平下芒草植株及種子細菌相對豐度表
2.3 16s功能預測分析
如圖6 所示,在php 與phps 中,其功能性強的前五類均為:信號轉導組氨酸激酶、糖基轉移酶、由CheY 樣受體域和翼螺旋DNA 結合結構域組成的響應調節劑、藍藻中保守的未表征蛋白質、未表征的低復雜度蛋白質,且種子內部功能豐度值高于植株。其中植株與種子中功能性最強的均為信號轉導組氨酸激酶。組氨酸激酶可以作為細菌細胞中的重要信號感知分子,細菌可以利用組氨酸激酶來感受胞外環境的變化,作為一種自身具有激酶、磷酸轉移酶和磷酸酶活性的多功能酶,組氨酸激酶能夠作為信號分子的細胞受體,且具有跨細胞膜信號轉導的功能。這種轉導信號在細菌、酵母菌和高等植物中都有發現[10-11]。

圖6 COG功能預測熱圖
3 討論與結論
本研究以磷石膏上芒草植株和種子為研究對象,利用高通量測序技術對微生物多樣性進行細致研究,結果顯示:在門水平上,其共同的優勢菌門是藍菌門。在屬水平上,鞘氨醇單胞菌屬細菌(Sphingomonas)、鏈霉菌屬細菌(Streptomyces)為優勢菌屬,且php 中兩種優勢細菌相對豐度高于phps。這兩種細菌在植物生長及抗逆性方面都起著較為重要的作用。黃媛林等對一株紅樹林鏈霉菌所產抑菌活性化合物的分離及其生物合成基因簇的研究表明,鏈霉菌的次生代謝產物具有多種抗性的功效,且多被用作生物防治劑[12]。何曉紅等對鞘氨醇單胞菌的各項功能的綜述研究表明,鞘氨醇單胞菌能夠耐受惡劣環境及貧瘠營養環境,且能夠促進植物生長與抵抗多種植物病原菌[13]。
目前,鏈霉菌屬與鞘氨醇單胞菌屬已廣泛用于重金屬污染的土壤植物與植物促生菌的聯合修復研究[14],微生物環境修復仍然是目前研究的熱點問題之一。然而現階段對于磷石膏的處理利用及其內部元素對于環境污染的系統研究還比較缺乏,這些都將成為未來磷石膏堆渣污染研究的重要課題。芒草植株能夠在磷石膏堆渣上面生存的原因可能是由于鏈霉菌與鞘氨醇單胞菌的存在,這兩株細菌具備開發成為磷石膏堆渣治理與利用的植物促生菌的潛在價值,值得進一步開發與研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
