動物蛋白源免疫調節肽的研究進展
2023-09-10 05:20:08梁泳儀白衛東肖更生曾曉房
食品工業科技 2023年18期
梁泳儀,王 宏,白衛東,肖更生,曾曉房
(仲愷農業工程學院,廣東省嶺南特色食品科學與技術重點實驗室,農業農村部嶺南特色食品綠色加工與智能制造重點實驗室,輕工食品學院,廣東廣州 510225)
免疫調節肽是一類具有免疫調節作用的生物活性肽,其作用包括但不限于促進/抑制淋巴細胞分化成熟、調節巨噬細胞的吞噬功能、轉移免疫信息、調節細胞分泌因子等[1-2]。免疫調節肽在某些情形下可以刺激先天和適應性免疫系統的細胞,增強免疫反應,為宿主提供抵抗感染的能力[3],而也有報道表明它們可以抑制免疫反應,從而抑制過敏和炎癥[4]。
研究表明,天然來源的免疫調節肽安全可靠、有效且成本較低[5]。20 世紀80 年代以來,免疫調節肽逐漸成為熱門研究,主要集中在提取工藝、結構鑒定和作用機理等。免疫調節肽的來源較為廣泛,涵蓋了動物源以及植物源等。其中以動物蛋白源的免疫調節肽占據了重要的地位,因為這類免疫調節肽具有極高的可食用安全性、良好的營養等特性以及廣闊的發展前景。到目前為止,研究人員已從動物源蛋白中鑒定出許多具有免疫活性的肽段,如雞胸骨軟骨中的P3-2-4肽段,鮑魚內臟中的AVPP 肽段等[6]。目前用于制備動物蛋白源免疫調節肽的原材料主要來自禽蛋、海洋動物、乳蛋白、哺乳動物、昆蟲以及其他動物等。此外,免疫調節多肽可能還具有其它活性,如:抗炎、抑菌、抗癌等[7]。
研究者在動物蛋白源免疫調節肽的提取純化、鑒定和表征、結構和功能等方面開展了大量研究,其免疫機制的相關研究相對分散、仍不明確[8]。因此,本文對動物蛋白源免疫調節肽的構效關系、免疫作用機制和不同動物蛋白源免疫調節肽的研究進展進行了綜述。
1 免疫調節肽的構效關系及作用機制
1.1 免疫調節肽的構效關系
免疫調節肽的活性與其結構有著密不可分的關系,其中,氨基酸組成、末端氨基酸、鏈長、總分子質量、排列順序、疏水性和空間結構等都會影響其生物活性[9]。
疏水性氨基酸能增強多肽的免疫調節活性,可能是通過增強了肽和細胞膜之間的相互作用來調節下游信號通路[10-14]。多數免疫調節肽含有疏水性氨基酸,如Li 等[13]從家蠶蛹蛋白水解物中分離出免疫調節肽DHAV,在這個多肽氨基酸序列中,一半的氨基酸是疏水性氨基酸(Ala 和Val)。何小慶[12]從波紋巴菲蛤中篩選得到的二肽TL 中含有疏水性氨基酸色氨酸(Trp)和亮氨酸(Leu)。此外,Cao 等[15]解析雞胸骨軟骨II 型膠原中的免疫調節肽P3-2-4結構發現,氨基酸組成中帶有較多的谷氨酸、精氨酸,亦對免疫調節活性起著重要的作用。在肽的N 端或C 端區域內富含精氨酸的多肽,還可以與T 細胞膜進行受體識別,進而使T 細胞產生淋巴因子[16],起到免疫調節的作用。
帶電荷的肽類似于趨化因子,可以結合免疫細胞的受體,激活免疫反應[17]。研究發現,肽段在相同pH 條件下,陰離子肽和陽離子肽對不同組分電荷的影響不同。負電荷部分對誘導型一氧化氮合酶(iNOS)活性的促進作用與動物蛋白水解產物相同,而正電荷部分對iNOS 的免疫調節作用較小[18-19]。Abachi-Hokmabadinazhad[18]在pH 為3 的實驗條件下,利用電滲析-超濾法(EDUF)分離出大西洋鯖魚免疫調節肽(pH3 EDUF),發現該多肽的負電荷部分具有最顯著的免疫調節作用。此外,Lebetwa 等[20]發現酪蛋白肽(CPPIII)的負電荷部分還具有抗病毒活性。而正電荷部分可通過與陰離子細菌膜的靜電相互作用,導致細菌死亡[21],進而對免疫系統有調節作用。
分子質量是與免疫調節活性相關的重要參數之一[22]。大多數免疫調節肽的分子質量小于2 ku[23-26],主要原因是分子量較小的肽(2~10 個殘基)溶解性更好,并能與酶活性位點有效協調,還能夠通過腸道屏障,并直接影響體循環中免疫細胞[27]。例如,Narayanasamy 等[24]從蟹腿肌肉中分離的低分子量肽,序列為LGLGAAVL(713.456 Da),可抑制COX-2 的表達,從而得知該多肽具有潛在的免疫調節活性。牦牛骨水解物中含有高比例的低分子量免疫調節肽YBHs(<2000 Da),可使血清總蛋白和白蛋白濃度升高,證明YBHs 能減緩環磷酰胺(CTX)誘導免疫抑制小鼠的免疫功能下降[26]。
隨著生物信息學的發展,根據上述多肽的結構特征預測其是否具有免疫調節活性與其在生物體內吸收、分布、代謝、排泄等生理性質[28]。如利用PeptideCutter、ProtParam、Blast 和BIOPEP-UWM 等分析平臺對已知蛋白序列進行模擬酶切[2,29]。綜上,大部分具有免疫調節作用的動物蛋白源肽由2~10 個氨基酸組成,且具有疏水性。雖然不同來源的免疫調節肽的結構或活性都存在差異,但是仍然為新型免疫調節肽的發掘與活性研究提供了指導意義。
1.2 免疫調節肽的作用機制
1.2.1 免疫調節肽對先天性免疫的影響 大量研究表明,先天免疫是非特異性的,主要通過機體的天然屏障、生理學防御、細胞或炎癥因子提供第一道防御[30]。而巨噬細胞、樹突狀細胞和自然殺傷細胞是為先天免疫系統發揮作用的主要細胞群[31]。
巨噬細胞由血液中循環的單核細胞祖細胞經血管壁遷移到組織后分化而成[32],主要作用是吞噬病原體和死亡或垂死的細胞。不同的細胞因子還可激活巨噬細胞,使其分化為M1 和M2 兩種表型[33]。研究發現,醋蛋中的三種免疫組分(SVE、FVE 和VVE)可誘導人單核細胞(U-937)分化成M1 表型的巨噬細胞[34]。Gu 等[35]從巖鯛中合成了多肽BO18,BO18能增強巨噬細胞的呼吸爆發能力。從青蛤提取的多肽SCSP 能增強巨噬細胞的吞噬能力[36]。Szmyt等[37]人工合成的蛋黃肽可激活巨噬細胞并使其分泌TNF-α。不僅如此,免疫調節肽還能促使巨噬細胞遷移,如Hu 等[38]從水蛭中分離得到了序列為EAGSAKELEGDPVAG 的寡肽,發現水蛭肽激活c-Junn-末端激酶(JNK)抑制劑和p38 絲裂原活化蛋白激酶(p38 MAPK)抑制劑,從而降低了巨噬細胞的抗遷移能力。馮挺等[39]提取的蜂毒肽可增強單核巨噬細胞的吞噬功能,以實現對腫瘤細胞的殺傷作用。但當免疫反應過度激活時,馬蠅唾液腺中的HA 可以抑制幾個單核-巨噬細胞表面標記物(CD115,IRF-8,pu1 和EGR-1)的mRNA 水平,從而降低巨噬細胞的過度激活[40]。
樹突狀(DC)細胞是機體功能最強的專職抗原遞呈細胞,通過激活初始T 淋巴細胞調控并維持免疫應答的中心環節[41-42]。研究發現,多肽LL-37 可在體外誘導DC,進而顯著上調吞噬細胞的吞噬能力,并釋放相關免疫因子[43]。Kim 等[44]發現了中華稻蝗的免疫組分(命名為Ocs-P),Ocs-P 能誘導DC 的表型改變,刺激輔助T 細胞(Th 細胞)和細胞毒性T 細胞(Tc 細胞)產生。
自然殺傷細胞又稱為NK 細胞,是機體重要的先天免疫細胞。NK 細胞是具有抗腫瘤、抗病毒和免疫調節作用的免疫細胞,但是在某些情況下參與超敏反應和自身免疫性疾病的發生[45]。從牦牛骨中提取的多肽YBHs 可通過增加NK 細胞活力來調節小鼠的細胞免疫功能[26]。不僅如此,王敏等[46]研究鮑魚水解肽(AHP)對小鼠NK 細胞的影響,發現陰性對照的NK 細胞活性為32.92%±5.81%,低中高劑量的NK 細胞活性分別為(38.58%±6.33%、42.34%±6.47%和45.69%±7.38%),證明AHP 能顯著提高NK 細胞活性,使NK 細胞對T 淋巴細胞的應答性增強,從而提高機體的免疫功能。
1.2.2 免疫調節肽對適應性免疫的影響 當先天性免疫中的細胞無法迅速應對病原體侵襲時,適應性免疫系統將被激活[47]。適應性免疫,可分為細胞免疫和抗體介導的體液免疫[48-49]。巨噬細胞分泌細胞因子進行免疫調節,屬于體液免疫的一種。當細胞受到刺激或損傷時,細胞會釋放細胞因子,從而影響細胞生長、活化、炎癥、免疫、組織修復或組織纖維化[34]。免疫調節肽不僅可以通過產生促炎細胞因子激活巨噬細胞,如青蛤中的多肽SCSP[36];同時還通過釋放抗炎細胞因子抑制巨噬細胞的過度激活,如馬蠅中的HA[40]。具體來說,Li 等[14]發現的牡蠣多肽DNSIAMESMK(P1)和LLQLGSGR(P2)和He 等[50]發現的鴨蛋卵清酶解物PH,均能誘導巨噬細胞產生一氧化氮(NO)、腫瘤壞死因子α(tumor necrosis factorα,TNF-α)和白細胞介素-6(interleukin-6,IL-6)。類似的,牡蠣寡肽[51]可誘導RAW264.7 巨噬細胞TNF-α和IL-6 的產生,在免疫反應中發揮關鍵作用。Adams等[52]發現發酵乳中某β-酪蛋白源的多肽可誘導巨噬細胞產生IL-8 和IL-1β。歐洲鰻鱺中的EP[53]可增加巨噬細胞中iNOS 的mRNA 水平、蛋白水平,進而使iNOS 產生NO。Sutthiwanjampa 等[4]發現在文蛤酶解組分BVH 濃度分別為0.5、1、5 mg/mL 的情況下,可刺激包括TNF-α、IL-1β、IL-10、IL-12 在內的其他細胞因子mRNA 表達。但是過度的免疫反應會導致炎癥。免疫細胞不受控制地產生促炎細胞因子(如NO、PGE2、TNF-α[54]、IL-1β和IL-6[25,55])會導致組織損傷和免疫功能喪失[19]。Wang 等[56]從白鱗魚鯊魚中分離復合肽(SCP),得知SCP 抑制血清中TNF-α、IL-6、IL-8 和NO 的水平,在脂多糖(LPS)刺激的小鼠中表現出明顯的抗炎活性。
同時巨噬細胞或DC 細胞將抗原傳遞給B 淋巴細胞與T 淋巴細胞,可進入細胞免疫階段[49]。B 細胞和T 細胞均在適應性免疫中發揮著重要的作用。B 細胞主要分布在淋巴結和脾臟中。當B 細胞受到特異性抗原刺激時,通過同型轉換機制能夠分化成為漿細胞,漿細胞可產生抗體,主要包括免疫球蛋白A(IgA)、IgG 和IgE[32]。Li 團隊[57]發現阿拉斯加鱈魚肽APP 可增強IgA+B 細胞的最終分化,促進漿細胞分泌,增加IgA 含量,從而提高機體免疫。然而,IgE 的過量會導致過敏反應。IgE 與兩個主要高親和力受體(FcεRI 和FcεRII)結合,這種相互作用導致了致敏狀態,使肥大細胞會釋放化學介質[32],從而刺激記憶B 和T 細胞反應,放大2 型輔助型T 細胞(Th2)和IgE 反應[58-59]。Th2 細胞會分泌IL-4、IL-5 和IL-13[32]。而口服酪蛋白磷酸肽(P-CPP)[60]可減輕IgE 介導的過敏反應,發揮免疫調節作用。
大多數T 細胞可根據其表面分化抗原簇(CD)分子的表達分為CD4+或CD8+兩個亞群[61]。CD4+T 細胞的功能主要是傳遞抗原信息,并輔助其他免疫細胞進行免疫活動[62]。CD8+T 細胞可以殺死被病毒感染的細胞或有可能導致癌癥的突變細胞[61]。由蜂毒、蛇蛉中提取的免疫調節肽可刺激CD4+T 細胞和CD8+T 細胞產生γ干擾素(IFN-γ)[39,44]。
1.2.3 免疫調節肽參與免疫調節作用的其他作用機理 免疫調節肽不僅能夠通過細胞免疫和體液免疫來平衡免疫功能,還通過胞內外信號轉導途徑來完成免疫反應[2]。研究表明,免疫調節肽發揮作用主要與核因子激活的B 細胞的κ-輕鏈增強(NF-κB)通路和絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)通路有關[19]。He 等[50]發現鴨蛋卵清蛋白酶解物PH 能通過與Toll 樣受體(TLR)2 和TLR4 相互作用,激活NF-κB 和MAPK 通路;Sutthiwanjampa團隊[4]發現文蛤水解組分BVH 通過刺激巨噬細胞活化,使磷脂酰肌醇-3 激酶(PI3K)激活蛋白激酶B(Akt)參與調節MAPKs(p38、ERK 和JNK),進而刺激各種細胞因子(TNF-α、iNOS 等)的產生;Hall等[63]發現蟋蟀蛋白水解物CPHD 通過抑制NF-κB mRNA 水平的激活發揮抗炎作用。此外,Yao 等[53]發現歐洲鰻鱺中的多肽EP 可誘導IκB(NF-κB 的抑制劑)磷酸化,從而導致巨噬細胞中p65、p38、JNK和ERK 磷酸化增加。
Feng 等[64]在鳥類中樞體液免疫器官法氏囊(BF)中分離到了一種多肽(BSP-I),通過增強凋亡誘導因子Bax 蛋白表達,增加抗腫瘤蛋白p53 的表達導致細胞凋亡,誘導細胞產生IFN-γ和IL-4,從而抑制腫瘤細胞增殖。Ji 等[65]純化了從以色列暗褐中分離到蝎毒肽Smp76,發現其激活干擾素調節因子3(IRF3)磷酸化,上調β干擾素(IFN-β)的表達,增強I 型干擾素(IFN)反應從而抑制病毒感染。此外,馮挺等[39]發現蜂毒肽能激活磷脂酶A2(PLA2),且肽段與PLA2 與腫瘤細胞相互作用后,可水解腫瘤細胞膜Band-3 蛋白,即具有抗癌作用。
綜上所述,免疫調節肽可通過多種路徑發揮作用(圖1)。由于機體的整體免疫調節能力是多種因素綜合作用的結果[23],不同動物蛋白來源的免疫調節肽往往表現出不同的作用,如激活免疫細胞的功能,促進各種細胞因子的分泌等。這些免疫調節肽究竟是如何發揮作用的分子機制,其構效關系值得進一步探索。
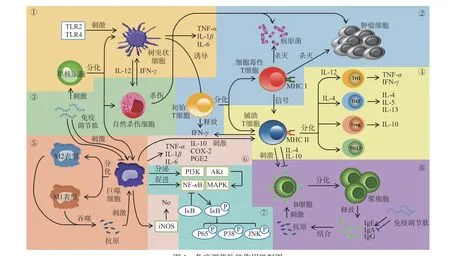
圖1 免疫調節肽的作用機制圖Fig.1 Mechanism of immunomodulatory peptides
2 不同動物蛋白源的免疫調節肽
2.1 禽及禽蛋類來源的免疫調節肽
禽類多肽具有免疫調節作用,可能有助于刺激機體免疫反應和預防微生物感染[66],在食品工業和醫學治療中發揮重要作用。Yu 等[67]發現5%雞肌肉酶解肽可使大鼠脾臟有較高比例的巨噬細胞、樹突狀細胞及較低比例的抗原呈遞細胞,進而表現對幼齡健康大鼠有免疫調節作用。此外,研究人員發現羽毛、骨頭等家禽的副產物中也可以得到相應的免疫調節肽,不僅可以提高這些副產物的附加值,還可以有效緩解副產物的環境污染。Sukaboon 團隊[22]從雞羽毛粉水解物中鑒定了一種序列為SNPSVAGVR(885.97 u)的多肽,該多肽降低了脂多糖誘導的RAW 264.7 細胞中促炎細胞因子iNOS、TNF-α、COX-2和IL-6 的基因表達。Cao 等[15]從雞胸骨軟骨II 型膠原(CII)中分離免疫調節肽,發現P3-2-4組分(RGQ LGPM)促進淋巴細胞增殖活性最高(85.54%),與人白細胞抗原DRB1(HLA-DRB1)分子的結合能力最好,也是一種有較高免疫狀態的多肽。
禽蛋類免疫調節肽具有巨大的研究潛力[15]。雞蛋可以降低44%~61%的大鼠結腸損傷,對預防和治療腸道損傷具有一定的治療價值[68]。不僅如此,雞蛋中免疫調節肽還具有抗癌作用,可誘導癌細胞凋亡,預防DNA 損傷,降低癌細胞的侵襲能力,并在各種癌細胞株中表現出細胞毒性和抗誘變活性[9]。而且,雞蛋多肽可以調節細胞因子(TNF-α、IL-1β、IL-6、IL-8、IL-10 和IL-12)的產生,并影響多種細胞系中炎癥介質(NO、PGE2、iNOS、COX-2)的產生[69]。就蛋類的不同部位而言,大部分卵黃多肽,都可促進細胞因子和NO 的分泌,因此可以作為治療免疫紊亂的天然活性物質[69]。Szmyt 等[37]利用優化后的大腸桿菌人工合成了蛋黃肽復合物,產率約為1.1 mg/L,其免疫活性可與天然蛋黃肽復合物相媲美。該多肽通過刺激TNF-α和IL-10 的產生和誘導NO 表達而具有免疫調節活性。此外,由卵轉鐵蛋白衍生的Ile-Arg-Trp 和Ile-Gln-Trp 可以通過調節結腸內微生物組成來改善腸道健康,緩解結腸炎癥[70]。卵轉鐵蛋白衍生肽在食品工業中作為抗微生物補充劑,在醫藥工業中作為抗癌、抗病毒和金屬補充劑具有很大的潛力。現已有大量研究證實蛋源多肽具有調節免疫的功能,相比之下對禽類的免疫活性組分的研究較少;因此研究禽類免疫調節肽的結構與活性之間的關系顯得尤為重要。
2.2 海洋動物來源的免疫調節肽
海洋動物有著豐富的蛋白質(10%~47% w/w)[6],通過酶解和細菌發酵生成的多肽具有多種功能和生物活性,包括抗氧化、降血壓、增強免疫力、抗炎和神經保護作用等[8,25]。目前為止,研究人員對鱈魚、海藻、牡蠣和貽貝類等來源的免疫調節肽研究較多,這些免疫調節肽均顯示出廣泛的調節免疫功能。阿拉斯加鱈魚肽(APP)中純化得到兩個具有較高免疫活性的肽段,分別是Gly-Val-Ile-Lys 和Ala-Cys-Asn-Gly-Arg,可保護環磷酰胺(Cy)引起的腸道屏障破壞[57]。Yao 等[53]從歐洲鰻鱺中分離得到了誘導NO 活性最強的Fr-1-1,可以激活巨噬細胞,進而提高免疫力。Narayanasamy 等[24]從蟹腿肌肉中分離了一種低分子量肽CL。用MTT 法測定在CL 刺激下LPS 誘導RAW264.7 細胞的細胞活力,在CL 濃度為50 μg/mL 時,最大細胞活力為81.2%±0.98%,證明CL 可刺激巨噬細胞的激活。Sutthiwanjampa等[4]研究文蛤酶解組分BVH 的免疫調節活性,發現在BVH 濃度在0.5~5 mg/mL 范圍內,可通過激活PI3K/Akt 和NF-κB 通路刺激RAW 264.7 細胞,從而刺激NO 以及其他細胞因子的產生,發揮免疫調節活性。青蛤水解產物中純化得到的免疫調節肽SCSP 可以通過激活NF-κB 信號通路刺激巨噬細胞活性[8]。Rathnapala 等[70]研究發現加州海螺的毒素肽可促進T 淋巴細胞增殖和Th1、Th2、Th17 和Treg細胞的分化。與此同時,海產品在加工過程中會產生大量的副產物,如魚皮、魚骨、魚頭和內臟。這些副產物通常會丟棄或降級生產飼料。因此利用低成本的海洋動物副產物開發免疫調節肽等活性成分,用于功能食品、保健品以醫藥和化妝品,可提高附加值[8],在功能性食品和營養食品行業具有潛在的應用前景[21]。Chataigner 等[71]利用海洋副產物作為原料進行酶解。研究發現,酶解液降低了促炎細胞因子IL-6、IL-1β和TNF-α的mRNA 表達量,因此該酶解液可作為一種潛在預防和抵消神經炎癥的保健品來源。類似地,鮑魚內臟蛋白肽AVPP 可以從細胞免疫、體液免疫、非特異性免疫、免疫器官等多方面調節小鼠免疫功能[46,72]。海洋動物擁有獨特的生活環境,蛋白含量豐富,且蛋白質氨基酸組成不同于陸生動物,目前已從蛤類等甲殼類軟體動物、海洋魚類等海洋動物蛋白中提取得到免疫調節肽,但是仍然有很多具有潛力的海洋動物及其加工過程中副產物資源未得到充分開發利用,值得進一步挖掘。
2.3 乳蛋白來源的免疫調節肽
乳是一種優質蛋白質來源,乳蛋白來源的物質大都具有免疫調節能力,包括牛αs1-酪蛋白、牛αs2-酪蛋白、牛β-酪蛋白、乳清蛋白及其多肽[73],能促進淋巴細胞增殖、激活巨噬細胞、刺激抗體合成和細胞因子表達[74]。Zab?ocka 等[75]從羊或牛的初乳中提取了富含脯氨酸的多肽復合物(PRP),該多肽可誘導細胞因子的分泌和外周血白細胞的增殖。而由驢乳蛋白提取的免疫調節肽能夠促進脾淋巴細胞增殖,增強巨噬細胞的吞噬作用以及促進免疫球蛋白的合成,從而增強免疫系統對病原菌的抵抗能力[76]。
乳蛋白包括乳清蛋白和酪蛋白。研究人員用胃蛋白酶和胰酶體外消化獲得初乳乳清蛋白水解物(CWP),處理MDA-MB-231 細胞,該細胞早期凋亡率由10.82%提高至21.85%,血液單核細胞吞噬活性顯著增強了20%,該結果證實了CWP 可增強免疫細胞增殖和活化,有治療癌癥等慢性疾病方面的潛力[77]。酪蛋白的初級結構含有若干具有生物活性的氨基酸序列[78]。Lebetwa 等[60]通過給OVA 致敏小鼠灌食含0.05%酪蛋白水解物P-CPP 的飼料,6 周后發現P-CPP 可以使血清中總IgE 抗體水平降低12%(P<0.05),使脾臟中T 卵泡輔助細胞(Tfh)的數量由7.1%±0.5%增加至9.7%±0.6%,這表明CPP 的磷酸化可以緩解過敏原特異性IgE 調節的致敏反應。
酪蛋白來源的多肽還可以通過微生物發酵獲得[78]。Adams 等[52]發現利用LAB 菌株發酵酪蛋白而產生的多肽可以誘導IL-10 和NO 產生。類似地,用保加利亞乳桿菌PrtB92059水解酪蛋白發現其水解產物(包含β-CN)具有免疫調節活性[79]。而費氏丙酸桿菌CIRM-BIA 129 發酵乳制品產生的多肽可以保護免疫調節蛋白不被消化蛋白分解,以調節腸道免疫系統[80]。由奶牛和牦牛奶制成的乳山奶酪和奶扎奶酪可鑒定出309 和225 個肽段,其中45%的肽段具有免疫調節特性[81]。乳蛋白是最早用于制備免疫調節肽重要的蛋白來源之一,即便已經開展了大量的科學研究,但在產業發展上仍然任重而道遠。
2.4 哺乳動物來源的免疫調節肽
豬、牛、羊等常見哺乳動物的肉制品大都直接食用。近年來一些藥用哺乳動物來源的免疫活性肽得到研究人員關注,如養殖鹿源蛋白。汪濤[82]從鹿皮中通過液質聯用(HPLC-MS)鑒定出8 個具有較高免疫活性的氨基酸序列。而張凱月則從鹿胎中經分離鑒定出9 種鹿胎寡肽[83],為進一步研究鹿來源多肽的免疫機制奠定基礎。此外,肉制品加工過程中的副產物通過制備高附加值的免疫調節肽等活性物質,不僅提高了產品的利用率,同時也改善了環境。Gao等[26]利用牦牛骨水解物(YBHs)進行灌胃小鼠后,發現其白細胞、免疫球蛋白(lgA、lgG)、細胞因子(IL-1β、IL-6 和TNF-α)濃度及免疫細胞(脾臟淋巴細胞、NK 細胞)均有升高,表明YBHs 可通過增加先天免疫和適應性免疫來預防或改善免疫抑制,并可能對腸道菌群具有潛在的益生作用。現階段為了生產哺乳動物源的免疫調節肽,通常選擇其肌肉作為蛋白質來源。實際生產過程中利用皮膚、骨骼和血液等加工副產品作為免疫調節肽的制備來源,提高產品附加值,將具有更大的經濟效益和研究價值。
2.5 昆蟲類來源的免疫調節肽
昆蟲多肽是昆蟲蛋白經誘導產生的具有多種生物活性的小分子物質[84],部分具有免疫活性。例如,家蠶體內富含甘氨酸的免疫調節肽(BMGlvA2),BMGlvA2 可以有效減少小鼠脾的損傷、壞死以及脾臟內淋巴細胞和巨噬細胞的數量,還可以顯著降低炎癥小鼠體內的炎性細胞因子IL-1β、IL-6 和TNF-α的表達,從而進行免疫調節[85]。孫瑞坤[86]從方格星蟲蛋白中鑒定了2 種具有免疫調節作用的氨基酸序列,分別為Phe-Lys(FK)和Phe-Gln(FQ),這兩種多肽均可增強小鼠細胞免疫功能、體液免疫功能、巨噬細胞功能和NK 細胞活性。蜂蛹多肽處理小鼠巨噬細胞RAW264.7,可通過激活MAPK 信號通路從而增強其免疫活性[87]。Ji 等[65]從以色列暗褐蝎中分離到蝎毒肽Smp76,該多肽對登革熱病毒(DENV)和塞卡病毒(ZIKV)有明顯的抑制作用,具有獨特的機制,涉及I 型干擾素反應,可以作為一種增強宿主固有免疫的替代治療。考慮到安全性,這些昆蟲來源的免疫調節肽仍需要更深入的實驗進行驗證。
2.6 其他動物來源的免疫調節肽
在兩棲動物中,研究員發現從青蛙皮膚提取的多肽具有抗菌和免疫調節活性[88]。在林蛙油肽OR 中發現了具有免疫增強功能的多肽,主要體現在OR 可顯著增強肝巨噬細胞的吞噬能力和自然殺傷細胞活性;然而OR 對淋巴細胞增殖、腹膜巨噬細胞吞噬能力和免疫器官指標影響較小[89]。此外,王潔[90]發現林蛙骨肉小肽可以促進RAW264.7 細胞分泌TNF-α和IL-2,進而具有增強免疫的作用。盡管如此,這些新型免疫調節肽仍需要通過臨床試驗進行驗證,以期為拓展動物資源高值化利用及開發新型免疫調節劑提供理論基礎。
從動物蛋白水解物中分離鑒定出的免疫調節肽如表1 所示。
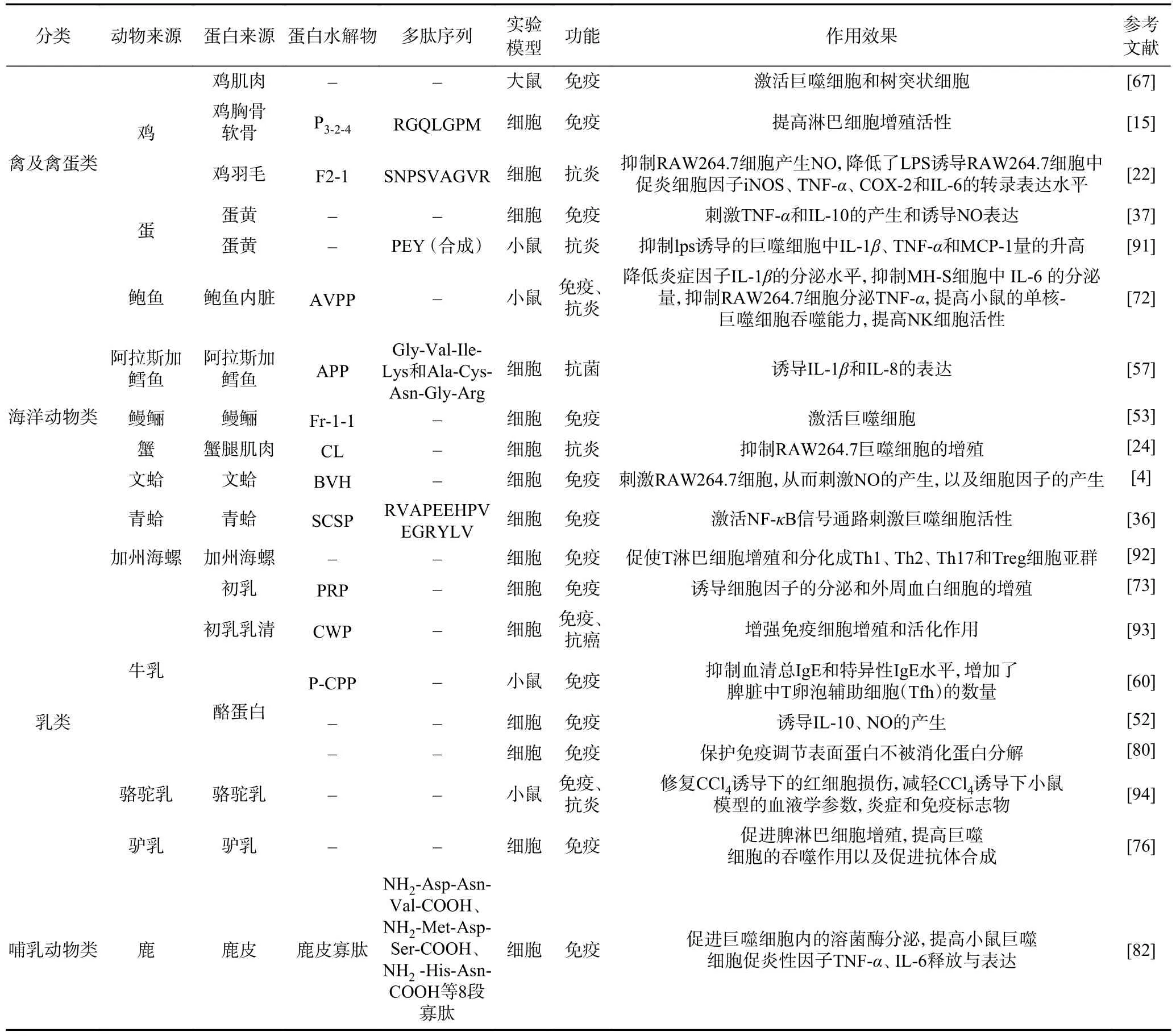
表1 動物蛋白源免疫調節肽的來源、功能、機制列表Table 1 Source, function and mechanism of food protein-derived immunomodulatory peptides
3 展望
由于具有安全、高效、成本較低等優勢,動物蛋白源免疫調節肽受到廣大科研人員的關注。免疫調節肽可以通過與代謝物的直接相互作用、改變酶的活性或基因表達、改變溶解度和促進微生物細胞膜穿孔等多種方式發揮其生物活性;但動物蛋白源免疫調節肽的開發仍然有較大的空間。目前制備動物蛋白源免疫調節肽的主流方法是酶解法,因其酶切位點的隨機性和不確定性,導致工業化生產分離純化免疫調節肽的成本增加;免疫調節肽在分子結構與生物功能關系的研究上仍處于基礎階段,明確免疫調節肽的關鍵作用位點和機制依舊任重道遠;此外,從動物蛋白中提取的免疫調節肽,其免疫活性大多在細胞或動物模型中得以證實,缺乏更多臨床研究的依據,從而阻礙了人類營養健康產品開發的進程。
綜上,未來動物蛋白源免疫調節肽的研究可從以下幾點出發:a.在傳統提取工藝的基礎上創新、優化純化免疫調節肽的技術;b.繼續深入探究這些免疫調節肽的分子作用機制,分析其構效關系,同時開發簡便有效的免疫活性評價方法,這可以為未來免疫調節肽的研究提供便利;c.在臨床研究上,須對免疫調節肽進行活性檢測和安全性評估,進一步明確其吸收、遷移、靶向和胃腸道穩定性等相關問題,以便更好地用于功能食品、醫藥領域。
