蘋果冷藏期間果肉褐變機制及控制研究進展
2023-09-10 01:21:49王濟瀚李富軍張新華李曉安張馨月孫雯雯
果樹學報 2023年3期
關鍵詞:機制
王濟瀚 李富軍 張新華 李曉安 張馨月 孫雯雯

摘要:果肉褐變是蘋果采后冷藏期間一種常見的生理性病害,特征各異,成因復雜,且不易檢測,嚴重影響了蘋果的貯藏和銷售。目前,細胞膜區室化的破壞被認為是果肉褐變的主要原因,許多研究就不同因素導致區室化的破壞提出了多種假設。通過參考近年來的研究成果,對蘋果果肉褐變的類型、成因和機制進行了總結;同時結合生產經驗,提出了減少蘋果冷藏期間果肉褐變的有效技術措施,展望了果肉褐變今后研究重點與方向,以期為蘋果冷藏期間果肉褐變的基礎理論研究和防控技術開發提供參考。
關鍵詞:蘋果;貯藏;褐變;機制;細胞膜區室化
中圖分類號:S661.1 文獻標志碼:A 文章編號:1009-9980(2023)03-0566-11
蘋果是世界上生產和消費最多的水果之一。中國蘋果產業協會數據表明,我國2020 年蘋果產量超過4400 萬t,居世界第一位。為保持蘋果新鮮品質并做到全時空供應,1/5~1/4 的蘋果會進入冷藏環節[1]。然而,蘋果冷藏過程中會發生一些生理病害,導致果實品質下降。其中,果肉褐變是蘋果貯藏期間常見病害之一。
蘋果冷藏期間的果肉褐變通常發生在果肉部分,有時也會波及果心或果皮,是單一或多種因素綜合作用的結果[1-2]。發生果肉褐變的蘋果不但風味和營養價值降低,而且會因為與健康果實在外觀上無明顯區別導致病害發生情況難以監控,造成巨大的經濟損失[2]。因此,對蘋果貯藏期間果肉褐變的成因、機制和控制技術進行研究具有重要的理論價值和實踐意義。
筆者在本文中綜述了近年來在蘋果冷藏期間常見的果肉褐變的分型、成因、發生機制和控制技術等方面的研究進展,以期為蘋果冷藏期間果肉褐變的理論和防控研究提供參考。
1 果肉褐變的分型
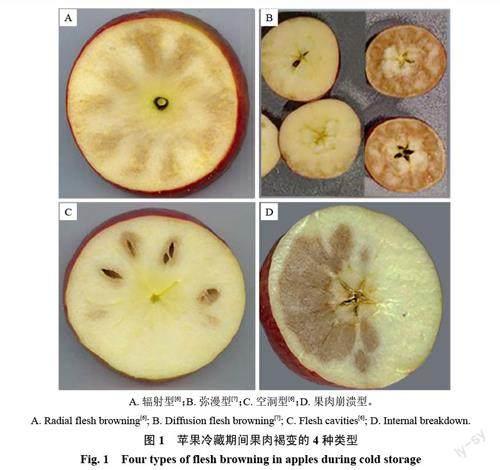
蘋果果肉褐變在冷藏期間或脫離原冷藏環境后都有發生,褐變部位的特征各有不同。質地或硬或軟,或濕潤或干燥,味道或苦或甜,有時還會形成空腔[3-5]。特征不同,成因也不同。區分不同的褐變特征,可以快速找到褐變的成因。常見蘋果冷藏期間的果肉褐變可以分為輻射型、彌漫型、空洞型和崩潰型4類(圖1)。
1.1 輻射型褐變
輻射型褐變主要表現為果實的維管束發生明顯褐變,而果皮相對不受影響(圖1-A)[6,8-9]。一般果柄端果肉褐變的嚴重程度略重于花萼端,褐變組織質地較軟,細胞壁受損嚴重[8]。此類褐變被認為是由衰老導致的維管束損傷引起,會隨著貯藏時間的增加而加劇,也會因為晚采、貯藏環境中高CO2和/或低O2濃度持續時間長以及貯藏溫度過低而增強其易感性[6,8-9]。
1.2 彌漫型褐變
彌漫型褐變的果實維管組織往往不受影響或褐變較輕,而果皮組織褐變明顯(圖1-B)[8]。彌漫型褐變被認為是一種冷害,常見于低溫生長的地區[7,9]。
當果實發育受到低溫脅迫時,果實的正常生理代謝活動會被破壞,導致果實發育滯后,以至于采后果實無法適應正常的低溫貯藏,進而導致彌漫性褐變的發生[7]。此類褐變一般在果實兩端較嚴重,中間部分較輕[6]。病害初期,僅在果皮組織附近形成小塊褐變,褐變區域有明顯界限(圖1-B左);隨著病害的加重,褐變向內發展,褐變面積變大,整個果皮組織附近全部褐變(圖1-B 右)。
1.3 空洞型褐變
空洞型褐變的特征是果肉褐變組織成點或片狀分布,在果肉組織之間形成褐變圈,并隨著癥狀的加重,褐變組織發展出透鏡狀的凹坑和空洞(圖1-C)[2,9]。空洞型褐變發生初期,果皮色澤變暗,有些品種會出現淺褐色斑點,果實內部產生塊狀褐變,果肩和果心線組織發病率較高,褐變組織的質地堅實、濕潤[2,10-11];隨后病癥迅速發展,褐變組織失水,質地變得松散、干燥,果實風味變淡,食之初甜后苦;最終,褐變組織產生凹坑和空洞,果皮出現褐斑和褶皺[3,5]。
1.4 崩潰型褐變
崩潰型褐變表現為果肉組織松散狀褐變(圖1-D)[12-13],可以細分為低溫崩潰、衰老崩潰和水心崩潰等[12]。低溫崩潰由果肉內層開始逐步向外側發展,初期褐變組織質地堅硬、濕潤,褐變顏色較淺,維管束無明顯褐變,后期發展為以果核為核心的圓球形大面積褐變,多發于元帥等品種[13],近幾年在富士、維納斯黃金等品種上也常見到。衰老崩潰多發于蘋果貯藏后期[14],常見有果肉粉棉病和橡皮病。果肉粉棉病發病初期,果肉發綿變軟,隨后變干形成易碎的粉質狀,后期果肉發生褐變,嚴重時果皮變暗、變褐或出現果皮破裂、果肉綻裂等癥狀[2];橡皮病常發于維管束和靠近果皮的組織,質地綿軟,病害前期果肉輕微褐變,隨后果皮變暗明顯,病害部位有輕微輪廓,手指按壓時容易陷下去,多發于紅玉蘋果,因此也常稱為紅玉橡皮病[13-14]。水心崩潰常見于采收時過熟的蘋果,表現為果肉的一部分成水浸狀,一般在果心處或圍繞維管束開始發展,具體發病部位及程度因品種而異,也會受到低溫、采收時間等因素的影響[12,15]。
盡管果肉褐變的類型不盡一致,但在實踐生產中往往發現2 種以上的褐變類型疊加發生,比如輻射型褐變后期往往也會發現空洞型褐變,而水心崩潰后期也往往伴隨著衰老崩潰。這種不同的導致果肉褐變的因素疊加,有時會造成區分困難,同時也增加了蘋果冷藏期間果肉褐變的風險和防控的難度。
2 影響因素
品種、采前氣候條件、栽培技術以及采后貯藏條件等,都會影響蘋果冷藏期間果肉褐變的發生。
2.1 品種
不同蘋果品種對果肉褐變的敏感性不同。一些品種由于果肉細胞排列緊密,果肉中的氣體交換較為困難,導致組織內部CO2濃度較高,易發生CO2傷害,進而造成果肉褐變,如富士、布瑞本(Breaburn)等[15]。Serra 等[16]對14 種蘋果采后褐變情況研究發現,青蘋和紅粉佳人品種不易發生果肉褐變,而嘉年華(Fiesta)和米切嘎拉(Mondial Gala)等品種易發生嚴重的果肉褐變。國內的主栽培品種中,富士、長富2 號和王林等品種對水心崩潰有較高的易感性,而金冠、國光和元帥等品種則不敏感[17-18]。
2.2 采前因素
氣候條件、土肥水管理、結果部位、作物負荷和采收期等栽培技術措施都可能影響到果實貯藏期間果肉褐變的發生。
2.2.1 氣候條件 (1)溫度:蘋果生長期溫度過低或過高都會造成果肉褐變的產生。有研究發現,在寒冷地區生長的布瑞本蘋果比在溫暖地區生長的更容易發生褐變,并推測其可能原因為寒冷的生長條件改變了細胞代謝過程,通過降低表皮和組織的擴散性/增強對高CO2和低O2的敏感性,增加了果肉褐變的風險[19]。McCormick 等[20]還發現,采前4 周環境溫度>10 ℃可以顯著降低果肉褐變的發病率,但具體機制不清楚。
(2)干旱:干旱對營養生長和果實發育具有重要影響。干旱環境下,根部對鈣等微量元素的吸收、利用和運轉受限[21],增強了對果肉褐變的易感性[22-23]。
此外,干旱導致的果實坐果率下降,也有可能通過降低果園負荷引起一系列包括果肉褐變易感性在內的果實品質差異[20]。
2.2.2 采收期 采收成熟度與果肉褐變聯系十分密切,過早或過晚采收均會增加果肉褐變的風險。其中,早采的蘋果易發生果心褐變[24],晚采的蘋果的CO2 傷害(空洞型褐變)、水心崩潰的發病率更高[24-25]。
2.2.3 礦質元素 土壤中營養元素和施肥的種類、數量、方式也直接影響果實采后貯藏質量。
(1)Ca:Ca2+能通過結合細胞膜上的磷脂和蛋白質來維持細胞膜的功能和穩定性[26],而果肉褐變與細胞膜系統的穩定性顯著相關。Ca2+含量過低會增大膜的通透性,甚至使細胞膜區室化被破壞,引起果肉褐變[23,27];但也有報道[28]指出,Ca2+含量過高會破壞細胞中Ca2+穩態機制,過多的Ca2+從細胞器或細胞膜外釋放到細胞質中,對細胞產生毒害。因此,Ca2+含量過低或過高都有可能引起果肉褐變的發生。
(2)N:N含量過高間接影響果肉褐變的發生。
采前大量施氮能促進果實的生長發育,氮鈣比增大,引起果實缺鈣[14,21]。此外,采前氮肥施入量過多,會使果實呼吸強度增大,加快果實衰老,引起衰老崩潰。
(3)Mg和K:Ca2+和Mg2+之間具有離子相似性,而K+又是Ca2+和Mg2+的拮抗劑,這導致Mg2+和K+與Ca2 +之間的互相競爭引發細胞膜功能的紊亂[29]。
Corrêa 等[30]發現,發生果肉褐變的果實具有更高的Mg/Ca、K/Ca和(K+Mg)/Ca。
2.2.4 結果部位 結果部位對果肉褐變易感性的影響主要通過光照、溫度等因素實現。內膛果由于光照不足,在貯藏期間衰老更快,更易發生果肉粉綿病;而外層果常因光照過強,易發生水心崩潰[31]。此外,結果部位對果肉褐變的影響,還與其對鈣含量分布的影響有關,如分枝根部的蘋果往往比頂端的蘋果Ca2+含量更高,長穗蘋果的Ca2+含量比短穗蘋果的Ca2+含量高[21],而Ca2+含量較低的果實通常具有更高的果肉褐變風險[32]。
2.2.5 樹體負荷 相比于溫度、礦質元素等采前因素,作物負荷對果實質量具有更大的影響。比如枝條生長旺盛、果實過度稀疏等低負荷的果樹,發生蘋果水心病和果肉褐變的風險就大[33]。這可能與低負荷增大了果實單果質量,從而降低了果實內包括Ca2+在內的微量元素濃度有關[20,34]。此外,低負荷導致的果個增大減緩了果實內氣體交換,增加了果肉內部CO2的積累,增大了空洞型褐變的風險[31]。而高負荷量,則表現出與果心褐變的正相關性[20]。
2.3 采后因素
蘋果采后貯藏環境的溫度、氣體成分和外源物質處理等都會影響到果肉褐變的發生。
2.3.1 溫度 貯藏溫度不適宜會導致果肉褐變的發生。溫度過低容易引起低溫誘導的、由外而內的果肉彌散型褐變,或者由內到外的低溫崩潰。如Saba等[5]發現,與3 ℃貯藏的蘋果相比,0.5 ℃貯藏誘導了恩派(Empire)蘋果果肉褐變的發生;王志華等[4]的研究表明,-2 ℃貯藏的蘋果褐變指數高于0 ℃貯藏的蘋果褐變指數。
2.3.2 氣體成分 空洞型果肉褐變是典型的由高CO2傷害引起的褐變癥狀。氣調貯藏的蘋果在CO2濃度過高,以及普通冷藏的蘋果在因包裝不合適及庫內通風不良造成環境中CO2濃度過高等情況下,都可能引起空洞型果肉褐變的發生。低O2環境會加劇此類褐變的形成和發展[2,8],主要表現為果實硬度下降和果皮下陷的加速以及褐變組織擴大和空洞形成的加劇,且常伴隨輕微發酵味和腐爛現象的發生[14]。此外,貯藏溫度過高也會使呼吸強度提高從而引起CO2積累,導致空洞型果肉褐變的出現[3],或者因加快果實成熟引起貯藏后期的衰老崩潰。
Castroa 等[35]研究發現,紅粉佳人在5 kPa CO2+1.5 kPa O2條件下冷藏2 個月后,果肉褐變發病率顯著高于1 或3 kPa CO2+1.5 kPa O2中冷藏的蘋果,而在0.5 kPa CO2+1.5 kPa O2環境中沒有觀察到果肉褐變;同時發現,在1.5 kPa CO2條件下,將O2由1 kPa提升到19 kPa 時,果肉褐變發病率可由5%降低至1%。Argenta等[24,36]研究發現,在0.5 kPa CO2+1.5 kPaO2貯藏條件下,富士表現出彌漫型褐變的特征;在2.5 kPa CO2+1.5 kPa O2貯藏條件下,3 種不同品系的富士蘋果均會發生果肉褐變,且能同時觀察到空洞型褐變和彌漫型褐變2 種果肉褐變類型。
值得注意的是,貯藏環境中的高CO2含量一般不被認為是導致蘋果彌漫型和輻射型果肉褐變的主要因素,而是褐變發生中的一個疊加因素[7]。
2.3.3 外源物質處理 研究表明,1-甲基環丙烯(1-methylcyclopropene, 1-MCP)、鈣和二苯胺(diphenylamine,DPA)處理都可能影響到蘋果貯藏期間的果肉褐變。
(1)1-MCP:多篇文獻均報道了1-MCP 可以顯著延緩蘋果等多種水果采后衰老進程,降低了果肉褐變、水心褐變等的發生[37-39],但也有1-MCP使用不當增加果肉褐變的報道。如Koushesh 等[5]研究發現,1-MCP處理后0.5 ℃貯藏的恩派蘋果,果肉褐變的發病率和褐變嚴重程度均顯著高于3 ℃貯藏的,并提出這與低溫冷害關系密切;DeEll 等[40]也發現,1-MCP 處理的恩派蘋果在2.5 kPa O2+2 kPa CO2或1.5 kPa O2+1.2 kPa CO2條件下,果肉褐變的發病率顯著增加,并認為這可能與1-MCP處理增強了果實對CO2的敏感性有關。此外,1-MCP處理也被發現誘導了晚采的恩派貨架期間果肉的褐變[41]。由此推測,1-MCP處理后對果肉褐變影響效果的差異,可能與蘋果品種、冷藏溫度和氣體成分的控制等的差異有關。
(2)鈣:許多報道表明,采后一定濃度的鈣處理有利于蘋果保持良好品質。一方面,鈣可以作為直接或間接的自由基清除劑,維持細胞膜結構的正常功能;另一方面,鈣也可以提高果實的抗壞血酸(ascorbic acid,AsA)含量[42]。AsA 作為一種重要的抗氧化劑,在抑制果肉褐變方面發揮關鍵作用。如Zhao 等[43]研究發現,首紅經5%的CaCl2浸泡10 min處理后,顯著維持了AsA等抗氧化物質的含量。
(3)DPA:DPA對低溫崩潰和CO2傷害導致的果肉褐變有明顯的抑制作用。有報道表明,DPA可以降低蘋果對CO2的易感性;在配合低溫處理使用時,DPA也能明顯降低果心褐變率[44]。但由于DPA使用后的廢液對環境和人體存在潛在的危害,英國、德國等部分國家已禁止DPA的使用。
3 蘋果果肉褐變的機制
目前,主流理論認為,細胞膜區室化功能喪失造成的酶促褐變是導致蘋果冷藏期間果肉褐變的主要機制。正常情況下,多酚氧化酶(polyphenol oxidase,PPO)位于細胞質中,酚類物質位于液泡中,由細胞的膜結構將兩者分隔開;但當細胞膜結構被破壞或通透性發生改變時,蘋果中的酚類物質泄露,并在O2的參與下被PPO催化形成醌類物質,而后醌類物質聚合生成棕色色素,導致果肉褐變的發生[45-46]。
下文總結了可能導致蘋果冷藏期間果肉細胞膜區室化功能損失的機制。
3.1 Ca2+濃度改變 Ca2+可以通過與細胞膜表面的脂質和蛋白質結合來維持細胞膜結構的功能和穩定性。據報道,維持膜結構的功能需要超過0.1 mmol·L-1的游離Ca2+,而原生質體自身Ca2+維持在一個極低的水平(0.1~0.2 μmol·L-1),無法發揮類似的作用[23]。因此,當原生質體外Ca2+減少或Ca2+與質膜的結合被抑制時,會導致細胞膜結構被破壞,使細胞膜區室化的功能喪失,隨后酚類物質泄露并與多酚氧化酶反應生成醌類物質,最終導致果肉褐變。
一般認為,Ca2+減少導致的區室化功能喪失的可能機制有2種。一種機制是K+、Mg2+等與Ca2+競爭結合質膜上Ca2+結合位點,但結合后的K+、Mg2+等卻不能替代Ca2+發揮維持細胞膜功能的作用,促使細胞膜通透性增加,最終區室化被破壞(圖2)[23]。Freitas等[23]、Corrêa 等[30]和Shoffe 等[47]的研究均證實高K+/Ca2+比或Mg2+/Ca2+比增加了果肉褐變的發病風險。
另一種機制是細胞壁Ca2+結合位點的增加,使細胞膜外側有效Ca2+濃度減少,不足以維持細胞膜的功能,導致區室化被破壞。如Freitas等[23]研究發現,在蘋果貯藏過程中,果膠甲酯酶(pectin methylesterases,PMEs)基因表達上調和PME活性增加并在細胞壁上創造了更多的Ca2+結合位點,降低了細胞膜外側游離Ca2+的水平,膜透性增大,導致果肉褐變發病率升高。
此外,鈣含量低會減弱果實耐CO2能力,加速了由高CO2引起的果肉褐變的發生[22],也使果實更易發生水心病、苦痘病等病害以及貯藏期間的果肉崩潰[48]。劉野等[49]提出,貯藏環境中的CO2進入細胞后形成的CO32-,可以與細胞質膜上水溶性的Ca2+結合,使質體外有效Ca2+減少,導致細胞透性增大,并指出這是CO2傷害引起果肉褐變的一個可能原因。
但也有報道提出,Ca2+濃度過高也可能造成蘋果冷藏期間的果肉褐變,原因可能在于Ca2+濃度過高導致的鈣穩態機制受損,使過多的Ca2+從細胞器或細胞膜外釋放到細胞質中,從而對細胞產生了毒害作用[50-51]。
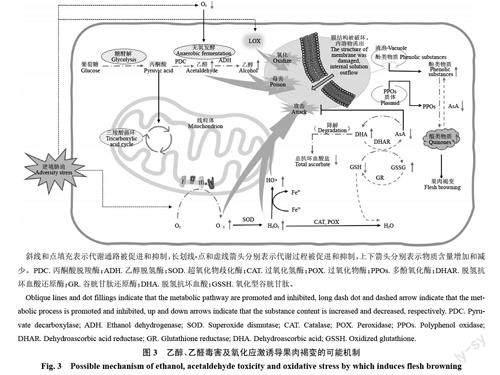
3.2 乙醇和乙醛毒害
在低溫、低O2和高CO2等逆境條件下,果蔬呼吸強度降低,有氧代謝會部分轉為無氧代謝。在此過程中,乙醇、乙醛等發酵代謝產物大量積累并對細胞膜結構產生毒害作用,導致細胞膜區室化功能喪失,果肉褐變風險增加(圖3)。分子水平上的研究表明,高CO2與低O2條件可通過下調果實PcMDHAR和PcAPX 表達,降低氨基酸-谷胱甘肽循環功能,減少AsA 含量,積累大量乙醇與乙醛,增加細胞膜透性,最終導致果肉褐變加重[52];Shoffe 等[47]研究也發現,冷害的嚴重程度與乙醇和乙醛的積累有關;Liu等[53]研究表明,發生水心褐變的果實中的乙醇和乙醛的濃度分別增加了710.26%和100.54%。
3.3 自由基及其清除體系失衡
正常環境下,果蔬組織內活性氧(reactive oxygenspecies, ROS)等自由基的產生與清除處于動態平衡,少量的ROS 不會對組織產生傷害,且充當了一種重要的穩態信號傳導實體[54]。但當果蔬處于逆境脅迫時,組織內ROS的異常積累超出了系統的清除能力,導致多種蛋白質和酶的結構與功能受到破壞,膜脂中的不飽和脂肪酸因ROS、脂氧合酶(lipoxygenase,LOX)等的攻擊發生過氧化反應,膜通透性增大,最終導致果肉褐變(圖3)[55]。
此外,許多非酶類物質,如AsA和谷胱甘肽(reducedglutathione,GSH)等可以參與ROS的清除,因而也在防止果肉褐變方面發揮著重要作用[45,56- 57]。
AsA還是一種高效的酚酶抑制劑,具有很強的抗褐變能力;GSH 可以通過與醌類物質相互作用,產生無色物質,抑制褐變的發生。但是,這些抗氧化物的活性很可能會在逆境脅迫下遭到破壞(圖3)。
3.4 能量虧缺
生物膜完整性的維持與足夠的細胞能量狀態密切相關,充足的三磷酸腺苷(adenosine triphosphate,ATP)可以穩定細胞膜的結構;較高的ATP含量也可以保持細胞內較高的ROS清除能力,減輕ROS對膜脂的過氧化作用,從而減少果肉褐變的發生[4,58]。當果蔬處于逆境脅迫下(如低溫、高CO2等),有氧呼吸的三羧酸循環(tricarboxylic acid cycle,TCA)減弱后,電子傳遞鏈偶聯的ATP減少,無法提供正常能量以維持細胞膜的結構和功能(圖4)[4,15];同時,乙醇、乙醛和ROS含量增加破壞了細胞膜結構完整性(圖3),進一步導致了ATP 合成受阻最終造成細胞膜區室化快速喪失,褐變加重[27]。王志華等[4]發現,低溫減少了ATP 和二磷酸腺苷(adenosine diphosphate,ADP)含量,導致細胞供能不足,從而加重了果肉褐變,且溫度越低,能量虧缺越嚴重,褐變指數越高。
Hatoum 等[15]研究發現,當將CO2 從0.7 kPa 升高到3.7 kPa 時,褐變發病率隨之升高,并認為這是高濃度CO2將呼吸代謝從三羧酸循環轉移到發酵過程,使ATP生成減少,導致維持細胞膜的能量不足,最終引起褐變。Wang 等[58]也發現,ATP 合成酶、NADH脫氫酶和無機焦磷酸化酶基因表達下調和磷脂酶D基因表達上調所導致的能量虧缺和細胞膜穩定性下降,是果肉褐變增加的一個重要原因。
綜上所述,能量虧缺和氧自由基傷害導致的膜通透性改變,可能是蘋果貯藏期間果肉褐變的主要原因,Ca2+等因為參與了細胞膜的穩定性,因而也對果實褐變有一定的影響,而無氧呼吸可以促進褐變的進程。
4 采后防止果肉褐變的措施
蘋果的品種,采前的氣候條件、采收期、土肥水管理、疏花疏果以及修剪等栽培技術措施都對蘋果貯藏期間的果肉褐變有著明顯的影響(見2.2 所述)。因此,選擇不易褐變的蘋果品種、適期采收、防止過度干旱以及合理負荷等都在一定程度上降低了果實貯藏期間果肉褐變的風險。
4.1 預冷
研究發現,采后及時預冷可以降低多種果蔬在貯藏過程中果肉褐變的發病率[59]。筆者在實踐生產中也發現,蘋果采后及時預冷,能有效減緩果實冷藏期間的果肉褐變,推測其機制可能是預冷可以快速降低果實的呼吸作用。這不僅減緩了果實成熟,推遲或減輕冷害的發生,還減少了呼吸產生的CO2過度積累帶來傷害的風險。
4.2 貯藏溫度
對蘋果應用低溫貯藏或氣調貯藏時,應綜合考慮果實的成熟度、采收期、品種特性等多種因素,以防止冷害造成的果肉褐變。對于富士等晚熟品種而言,適宜貯藏的溫度為(-1~1)℃(機械冷藏)或(-0.5~0.5)℃(氣調貯藏)。
4.3 氣體成分
用于氣調貯藏的蘋果需注意庫內氣體濃度,以防止O2濃度過低或CO2濃度過高導致空洞型褐變、輻射型褐變的發生。尤其是對于環境CO2敏感的富士和維納斯黃金,長期貯藏的這2 種蘋果理想氣體指標為CO2不超過1%,O2在5%左右。
4.4 外源物質
1-MCP 是目前國內外在富士等蘋果品種上常用的保鮮處理技術措施。1-MCP的使用方法和效果取決于蘋果的品種、貯藏時間及貯藏溫度等因素[60]。對富士蘋果而言,1-MCP的常用體積分數為0.5~1.0 μL · L- 1,一般在常溫(20~25 ℃)或低溫(0~2 ℃)條件下處理12~24 h,就可以有效減輕果肉褐變,同時提高果實其他貯藏品質[36,61]。
5 展望
目前,果肉褐變機制的研究仍集中于細胞膜區室化功能的喪失。然而,細胞膜衰老是果蔬采后成熟衰老過程中不可避免的自然事件。因此,啟動細胞膜衰老的關鍵因子和關鍵步驟是什么,貯藏環境中哪些因素可以導致細胞膜衰老加速或者有利于保持細胞膜結構的穩定性,值得研究。特別是對于不同類型的果肉褐變,它們之間細胞膜結構和功能變化有什么差異;為什么會出現輻射型、彌散型、空洞型和果肉崩潰型等明顯不同的褐變形式;這些不同褐變類型的發生,分別對應著哪些誘導果肉褐變的因素;它們之間有無共性的問題,存不存在互相影響的情況等,需要深入研究。只有深入研究和了解細胞膜衰老的機制、區分出不同褐變類型的誘導因素,才能更好地、有針對性地采取果肉褐變的防治措施。
對于有特殊需求,比如為追求貯藏期或者風味品質而特意早采或者晚采的蘋果,以及需要特定貯藏環境或特定采后處理需求的蘋果等,研究這些特定的農業技術措施或采前采后處理措施對果肉褐變發生規律的影響,探討其影響機制,尋找有針對性的防控技術措施,也具有很大的實際應用價值。
針對發生果肉褐變的蘋果外觀無明顯差異、僅靠肉眼無法準確判斷是否發病及發病程度的情況,開發更加靈敏的基于聲波、光波等原理的無損檢測技術及分檢設備,將有助于在蘋果貯藏和出庫、上市期間發生褐變果實的篩選分級,減少病果流入市場帶來的經濟和信譽損失。
根據生理狀態、采收期和貯藏環境等的差異和果肉褐變發生的機制,篩選鑒定果肉褐變相關的特定生理成分(標志物),研究建立基于標志物的蘋果果肉褐變發生的數學模型和預測預警技術,實現在蘋果入庫初期即對蘋果貯藏期間果肉褐變發生情況進行預測,或者根據生物標志物對地區或果園整體果實冷藏期間果肉褐變發生的概率提出預測預警,都有助于貯藏企業提前采取有針對性的預防或控制措施,減輕因果肉褐變帶來的風險和經濟損失。
參考文獻References:
[1] 李永新,王超,王麗巖,茹磊,陳敬鑫. 蘋果采后貯藏褐變研究
進展[J]. 渤海大學學報(自然科學版),2020,41(3):202-210.
LI Yongxin,WANG Chao,WANG Liyan,RU Lei,CHEN
Jingxin. Research progress on postharvest storage browning of
apple[J]. Journal of Bohai University (Natural Science Edition),
2020,41(3):202-210.
[2] 石建新,閆曉芳. 蘋果貯藏期間常見果肉褐變類型及防治[J].
落葉果樹,1995,27(3):35-36.
SHI Jianxin,YAN Xiaofang. Types and control of common flesh
browning of apple during storage[J]. Deciduous Fruits,1995,27
(3):35-36.
[3] 王杰,里程輝,王穎達. 蘋果貯藏期二氧化碳傷害及貯藏建
議[J]. 北方果樹,2019(3):50-51.
WANG Jie,LI Chenghui,WANG Yingda. Carbon dioxide damage
during apple storage and storage suggestions[J]. Northern
Fruits,2019(3):50-51.
[4] 王志華,賈朝爽,王文輝,佟偉,姜云斌. 低溫貯藏對‘金紅蘋果
能量代謝和品質的影響[J]. 園藝學報,2020,47(12):2277-2289.
WANG Zhihua,JIA Chaoshuang,WANG Wenhui,TONG Wei,
JIANG Yunbin. Effects of low temperature storage on energy
metabolism,related physiology and quality in‘Jinhongapple
fruit[J]. Acta Horticulturae Sinica,2020,47(12):2277-2289.
[5] SABAMK,WATKINS C B. Flesh browning development of‘Empireapple
during a shelf life period after 1-methylcyclopropene
(1-MCP) treatment and controlled atmosphere storage[J/OL]. Scientia
Horticulturae,2020,261:108938. DOI:10.1016/j.scienta.
2019.108938.
[6] JAMES H,JOBLING J. The flesh browning disorder of‘Pink
Ladyapples[J]. New York Fruit Quarterly,2008,16(2):23-28.
[7] TONG C B S,CHANG H Y,BOLDT J K,et al. Diffuse flesh
browning in‘Honeycrispapple fruit is associated with low
temperatures during fruit growth[J]. HortScience,2016,51(10):
1256-1264.
[8] JAMES H J,JOBLING J J. Contrasting the structure and morphology
of the radial and diffuse flesh browning disorders and
CO2 injury of‘Cripps Pinkapples[J]. Postharvest Biology and
Technology,2009,53(1/2):36-42.
[9] JAMES H,JOBLING J,TANNER D. Investigating structural
and physiological differences between radial and diffuse types
of flesh browning in cripps pink apples[J]. Acta Horticulturae,
2008,768:77-84.
[10] 李喜宏,宋壯興,田勇,馮小元,曹恩義. 新紅星蘋果貯藏前期
高CO2處理的極限值[J]. 中國果樹,1993(1):6-9.
LI Xihong,SONG Zhuangxing,TIAN Yong,FENG Xiaoyuan,
CAO Enyi. Limit value of high CO2 treatment in early storage of
Xinhongxing apple[J]. China Fruits,1993(1):6-9.
[11] 曲怡寧. 蘋果果實貯藏期二氧化碳傷害特性觀察與研究[D].
楊凌:西北農林科技大學,2019.
QU Yining. Observation and study on CO2 damage characteristics
of apple fruit during storage[D]. Yangling:Northwest A & F
University,2019.
[12] 福田博之,章祖涵. 蘋果果肉褐變的發生機制及其分類法[J].
煙臺果樹,1982(3):51-57.
FUTIAN Bozhi,ZHANG Zuhan. Mechanism and classification
of apple flesh browning[J]. Yantai Fruits,1982(3):51-57.
[13] 董啟鳳,楊克欽,肖永年. 蘋果的果內崩潰[J]. 遼寧果樹,1986
(1):49-52.
DONG Qifeng,YANG Keqin,XIAO Yongnian. Intra fruit collapse
of apple[J]. Liaoning Fruits,1986(1):49-52.
[14] 劉興華,陳維信. 果品蔬菜貯藏運銷學[M]. 北京:中國農業出
版社,2002.
LIU Xinghua,CHENWeixin. Fruit & vegetable storage and transportation
science[M]. Beijing:China Agriculture Press,2002.
[15] HATOUM D,BUTS K,HERTOG M,GEERAERD A,
SCHENK A,VERCAMMEN J,NICOLAI B. Effects of preand
postharvest factors on browning in braeburn[J]. Horticultural
Science,2018,41:19-26.
[16] SERRA S,ANTHONY B,BOSCOLO S F,MASIAA,MUSACCHI
S. Determination of post-harvest biochemical composition,
enzymatic activities,and oxidative browning in 14 apple cultivars[
J]. Foods,2021,10(1):186.
[17] DU M J,LIU Z T,ZHANG X T,LI H D,LIU Z Y,LI X H,
SONG J X,JIA X Y,WANG L Y. Effect of pulsed controlled atmosphere
with CO2 on the quality of watercored apple during
storage[J/OL]. Scientia Horticulturae,2021,278:109854. DOI:
10.1016/j.scienta.2020.109854.
[18] 姜云斌,王志華,賈朝爽. 蘋果水心病的發病機理研究進展[J].
中國果樹,2022(1):8-14.
JIANG Yunbin,WANG Zhihua,JIA Chaoshuang. Research
progress on pathogenesis of watercore in apple[J]. China Fruits,
2022(1):8-14.
[19] LAU O L. Effect of growing season,harvest maturity,waxing,
low O2 and elevated CO2 on flesh browning disorders in‘Braeburnapples[
J]. Postharvest Biology and Technology,1998,14
(2):131-141.
[20] MCCORMICK R J,BIEGERT K,STREIF J. Occurrence of
physiological browning disorders in stored‘Braeburnapples
as influenced by orchard and weather conditions[J/OL]. Postharvest
Biology and Technology,2021,177:111534. DOI:10.1016/
j.postharvbio.2021.111534.
[21] SAURE M C. Calcium translocation to fleshy fruit:its mechanism
and endogenous control[J]. Scientia Horticulturae,2005,
105(1):65-89.
[22] 王志華,姜云斌,賈朝爽,王文輝,劉偉. 河北承德寒富蘋果貯
藏期果肉褐變原因分析與建議[J]. 果樹實用技術與信息,2020
(6):43-45.
WANG Zhihua,JIANG Yunbin,JIA Chaoshuang,WANG Wenhui,
LIU Wei. Analysis and suggestions on flesh browning of
‘Hanfuapple in Chengde,Hebei province during storage[J]. Practical
Technology and Information of Fruit Trees,2020(6):43-45.
[23] DE FREITAS S T,AMARANTE C V T D,LABAVITCH J M,
MITCHAMA,E J. Cellular approach to understand bitter pit development
in apple fruit[J]. Postharvest Biology and Technology,
2010,57(1):6-13.
[24] ARGENTA L C,DO AMARANTE C V T,BETINELLI K S,
BRANCHER T L,NESI C N,VIEIRA M J. Comparison of
fruit attributes of‘Fujiapple strains at harvest and after storage[
J/OL]. Scientia Horticulturae,2020,272:109585. DOI:
10.1016/j.scienta.2020.109585.
[25] KWEON H J,KANG I K,KIM M J,LEE J,MOON Y S,CHOI
C,CHOI D G,WATKINS C B. Fruit maturity,controlled atmosphere
delays and storage temperature affect fruit quality and incidence
of storage disorders of‘Fujiapples[J]. Scientia Horticulturae,
2013,157:60-64.
[26] HIRSCHI K D. The calcium conundrum. Both versatile nutrient
and specific signal[J]. Plant Physiology,2004,136(1):2438-2442.
[27] 王雷. 蘋果果實不同O2/CO2 簡易氣調貯藏CO2 傷害特性研
究[D]. 楊凌:西北農林科技大學,2020.
WANG Lei. Study on the injury characteristics of CO2 in different
O2/CO2 storage of apple fruits[D]. Yangling:Northwest A &
F University,2020.
[28] WOOD R,PROSKE M,DE FREITAS S D,SCHEER C,
V?GELE T,NEUWALD D A. Seasonal variation in calcium
and ascorbic acid content at harvest related to internal browning
in‘Braeburnapple during controlled atmosphere storage[J].
Scientia Horticulturae,2022,297:110943.
[29] PRETI R,TAROLA A M. Study of polyphenols,antioxidant capacity
and minerals for the valorisation of ancient apple cultivars
from Northeast Italy[J]. European Food Research and Technology,
2021,247(1):273-283.
[30] CORR?A T R,STEFFENS C A,AMARANTE C V T D,
MIQUELOTO A,BRACKMANN A,ERNANI P R. Multivariate
analysis of mineral content associated with flesh browning
disorder in‘Fujiapples produced in Southern Brazil[J]. Bragantia,
2017,76(2):327-334.
[31] FERGUSON I,VOLZ R,WOOLF A. Preharvest factors affecting
physiological disorders of fruit[J]. Postharvest Biology and
Technology,1999,15(3):255-262.
[32] KALCSITS L,MATTHEIS J,GIORDANI L,REID M,MULLIN
K. Fruit canopy positioning affects fruit calcium and potassium
concentrations,disorder incidence,and fruit quality for
‘Honeycrisp apple[J]. Canadian Journal of Plant Science,
2019,99(5):761-771.
[33] KANAYAMAY,KOCHETOV A. Abiotic stress biology in horticultural
plants[M]. Tokyo:Springer,2015:127-145.
[34] MUSACCHI S,SERRA S. Apple fruit quality:Overview on preharvest
factors[J]. Scientia Horticulturae,2018,234:409-430.
[35] DE CASTRO E,BARRETT D M,JOBLING J,MITCHAM E J.
Biochemical factors associated with a CO2-induced flesh browning
disorder of Pink Lady apples[J]. Postharvest Biology and
Technology,2008,48(2):182-191.
[36] ARGENTA L,FAN X T,MATTHEIS J. Delaying establishment
of controlled atmosphere or CO2 exposure reduces‘Fujiapple
CO2 injury without excessive fruit quality loss[J]. Postharvest Bi-
ology and Technology,2000,20(3):221-229.
[37] LEE J,KANG I K,NOCK J F,WATKINS C B. Effects of preharvest
and postharvest applications of 1- methylcyclopropene
on fruit quality and physiological disorders of‘Fujiapples during
storage at warm and cold temperatures[J]. HortScience,
2019,54(8):1375-1383.
[38] MIN D D,DONG L L,SHU P,CUI X X,ZHANG X H,LI F J.
The application of carbon dioxide and 1-methylcyclopropene to
maintain fruit quality of‘Niuxinpersimmon during storage[J].
Scientia Horticulturae,2018,229:201-206.
[39] DIAS C,RIBEIRO T,ROBDRIGUES A C,FERRANTE A,
VASCONCELOS A W,PINTADO M. Improving the ripening
process after 1-MCP application:Implications and strategies[J].
Trends in Food Science & Technology,2021,113:382-396.
[40] DEELL J R,LUM G B. Effects of low oxygen and 1-methylcyclopropene
on storage disorders of‘Empireapples[J]. Hort-
Science,2017,52(9):1265-1270.
[41] JUNG S K,CHOI H S. Browning of early and late- harvested
‘Empireapples affected by cold storage and 1-MCP[J]. Agronomy,
2020,10(7):1050.
[42] NXUMALO K A,MATSUANE C,MASARIRAMBI M T. Calcium-
related post- harvest physiological disorders of fruits and
vegetables in eswatini:A review[J]. Current Journal of Applied
Science and Technology,2019,33(6):1-10.
[43] ZHAO Y,WANG C. Effect of calcium chloride in combination
with salicylic acid on post- harvest freshness of apples[J]. Food
Science and Biotechnology,2015,24(3):1139-1146.
[44] 劉麗丹,吳仲珍,李育生,張璐. 二苯胺控制采后果蔬生理病
害的機制[J]. 保鮮與加工,2014,14(1):53-56.
LIU Lidan,WU Zhongzhen,LI Yusheng,ZHANG Lu. The
mechanism of diphenylamine on controlling physiological diseases
of fruits and vegetables during storage[J]. Storage & Process,
2014,14(1):53-56.
[45] GLAGOLEVAAY,SHOEVAO Y,KHLESTKINA E K. Melanin
pigment in plants:Current knowledge and future perspectives[J].
Frontiers in Plant Science,2020,11:770.
[46] TANG T T,XIE X F,REN X,WANG W J,TANG X M,
ZHANG J,WANG Z D. A difference of enzymatic browning unrelated
to PPO from physiology,targeted metabolomics and
gene expression analysis in Fuji apples[J/OL]. Postharvest Biology
and Technology,2020,170:111323. DOI:10.1016/j.postharvbio.
2020.111323.
[47] AL SHOFFE Y,NOCK J F,BAUGHER T A,MARINI R P.
WATKINS C B. Bitter pit and soft scald development during
storage of unconditioned and conditioned‘Honeycrispapples
in relation to mineral contents and harvest indices[J/OL]. Postharvest
Biology and Technology,2020,160:111044. DOI:
10.1016/j.postharvbio.2019.111044.
[48] 王田利. 鈣對蘋果生長的影響及預防缺鈣措施[J]. 西北園藝
(果樹),2020(5):31-32.
WANG Litian. Effect of calcium on apple growth and measures
to prevent calcium deficiency[J]. Northwest Horticulture,2020
(5):31-32.
[49] 劉野,胡小松,張飛. 二氧化碳導致鴨梨褐變與細胞內鈣的關
系[J]. 食品科學,2011,32(13):62-65.
LIU Ye,HU Xiaosong,ZHANG Fei. Relationship between carbon
dioxide-induced browning and intracellular calcium in Yali
pears (Pyrus bretschneideri Rehd.) [J]. Food Science,2011,32
(13):62-65.
[50] DE FREITAS S T,PAREEK S. Postharvest physiological disorders
in fruits and vegetables[M]. Boca Raton:CRC Press,2019:
127-162.
[51] SCHMIDT R R,WEITS D A,FEULNER C F J,VAN DONGEN
J T. Oxygen sensing and integrative stress signaling in
plants[J]. Plant Physiology,2018,176(2):1131-1142.
[52] DEUCHANDE T,CARVALHO S M P,GIN?-BORDONABA
J,VASCONCELOS M W,LARRIGAUDI?RE C. Transcriptional
and biochemical regulation of internal browning disorder in
‘Rochapear as affected by O2 and CO2 concentrations[J]. Postharvest
Biology and Technology,2017,132:15-22.
[53] LIU X,FAN H M,LIU D H,LIU J,SHEN Y,ZHANG J,WEI
J,WANG C L. Transcriptome and metabolome analyses provide
insights into the watercore disorder on Akibae pear fruit[J]. International
Journal of Molecular Sciences,2021,22(9):4911.
[54] OLSON K R,GAO Y,DELEON E R,ARIF M,ARIF F,ARORA
N,STRAUB K D. Catalase as a sulfide-sulfur oxido-reductase:
An ancient (and modern?) regulator of reactive sulfur species
(RSS)[J]. Redox Biology,2017,12:325-339.
[55] CHEN C,JIANG A L,LIU C H,WAGSTAFF C,ZHAO Q,
ZHANG Y,HU W. Hydrogen sulfide inhibits the browning of
fresh- cut apple by regulating the antioxidant,energy and lipid
metabolism[J/OL]. Postharvest Biology and Technology,2021,
175:111487. DOI:10.1016/j.postharvbio.2021.111487.
[56] LEE J,CHENG L L,RUDELL D R,FOCK J F,WATKINS C B.
Antioxidant metabolism in stem and calyx end tissues in relation
to flesh browning development during storage of 1-methylcyclopropene
treated‘Empireapples[J]. Postharvest Biology and
Technology,2019,149:66-73.
[57] LI Z L,LI B R,LI M Q,FU X D,ZHAO X M,MIN D D,LI F
J,LI X A,ZHANG X H. Hot air pretreatment alleviates browning
of fresh-cut pitaya fruit by regulating phenylpropanoid pathway
and ascorbate-glutathione cycle[J]. Postharvest Biology and
Technology,2022,190:111954.
[58] WANG J W,JIANG Y G,LI G D,L? M,ZHOU X,ZHOU Q,
FU W W,ZHANG L,CHEN Y F,JI U J. Effect of low temperature
storage on energy and lipid metabolisms accompanying
peel browning of‘Nanguopears during shelf life[J]. Postharvest
Biology and Technology,2018,139:75-81.
[59] DUAN Y,WANG G B,FAWOLE O A,VERBOVEN P,
ZHANG X R,WU D,OPARA U L,NICOLAI B,CHEN K S.
Postharvest precooling of fruit and vegetables:A review[J].
Trends in Food Science & Technology,2020,100:278-291.
[60] ZHANG X H,LI F J,ZHAI H,SHU H R. Advances in the mechanism
of 1-MCP inhibiting ethylene action and its application in
apple storage[J]. International Agricultural Engineering Journal,
2011,20(2):135-140.
[61] LEE J,JEONG M C,KU K H. Chemical,physical,and sensory
properties of 1- MCP- treated Fuji apple (Malus domestica
Borkh.) fruits after long- term cold storage[J]. Applied Biological
Chemistry,2017,60(4):363-374.
猜你喜歡
四川勞動保障(2021年9期)2022-01-18 05:11:08
文苑(2018年21期)2018-11-09 01:23:06
當代陜西(2018年9期)2018-08-29 01:21:00
當代陜西(2017年12期)2018-01-19 01:42:33
暨南學報(哲學社會科學版)(2016年9期)2017-01-15 13:52:00
中國衛生(2016年9期)2016-11-12 13:28:08
中國衛生(2015年9期)2015-11-10 03:11:12
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國衛生(2014年3期)2014-11-12 13:18:12
中國火炬(2014年4期)2014-07-24 14:22:19
