融合根溫的奶油生菜光合速率模型預測
2023-10-04 22:45:27黎雪
山西農業科學 2023年3期
黎雪

摘要:綠色植物的光合作用受多方面因素的影響,因此,構建融合多種因素的光合速率模型是實現作物生長狀態高效預測的關鍵。試驗選取奶油生菜作為研究對象,設置多因素嵌套試驗,其中外界環境變量包括光量子通量密度、室溫、根溫、二氧化碳體積分數等,共獲得試驗數據648 組,利用Pearson 分析法對各因素與植物光合速率進行相關性分析,并選定根溫、葉溫、二氧化碳體積分數、光量子通量密度為建模輸入特征,在此基礎上構建基于LMBP 算法的奶油生菜光合速率模型。結果表明,根溫在0.01 水平上與植株的光合速率顯著相關。與未加入根溫的訓練結果進行對比,加入根溫后模型訓練集和測試集精度均有提高;訓練集均方根誤差為1.46×10-3 μmol/(m2·s),平均絕對誤差為1.873×10-3 μmol/(m2·s),測試集決定系數為0.995 26。基于LMBP 算法的奶油生菜光合速率模型考慮了根溫對植物光合作用的影響,可以實現光合速率的精準預測。
關鍵詞:根溫;預測模型;光合速率;相關性分析;LM 訓練法
中圖分類號:S636.2 文獻標識碼:A 文章編號:1002?2481(2023)03?0257?07
作為我國現代農業的重要組成部分,設施蔬菜栽培面積已達到我國設施園藝總面積的95% 以上,超過了全球總面積的80%[1]。隨著水培技術的發展,水培蔬菜以其環保無污染、營養均衡、品質良好等優點已成為研究熱點。在蔬菜培育過程中,葉片生長是提高其品質和產量的關鍵,而葉片生長又受光合作用的影響。光合作用是植物物質積累的核心反應,可將光能轉化為化學能,受溫、光、水、氣、肥等環境因子的影響。其中,水、肥等環境因子在設施條件下較為穩定,而考慮溫、光、氣等環境因子變化是精準預測水培蔬菜光合速率的關鍵。光照可為光合作用提供能量,光照不足會導致植物凈光合速率下降,嚴重影響農產品的產量和品質[2-4]。CO2是植物光合作用的主要原料,若濃度較低會限制作物的光合作用,浪費光熱資源,會成為設施栽培蔬菜產量和品質提高的關鍵限制因素[5-7]。氣溫不僅會通過影響與光合作用有關的酶活性來影響光合速率,還會通過影響蒸騰作用來影響光合速率[8]。
在水培蔬菜中,根系溫度也是影響作物生長和光合作用的關鍵。營養液溫度會影響作物根系對礦物質元素的吸收、光合速率的增減和干物質的積累。
根溫在綠色植物生長過程中扮演著極其重要的角色[9-10]。這些環境因素不僅影響作物光合反應,而且存在耦合作用[11]。因此,構建融合多種因素的光合速率模型是實現作物生長狀態高效預測的關鍵。
在植物光合速率預測模型的研究中,YE 等[12]在傳統光合生理模型的基礎上提出了不同類型的光響應模型,為光合模型研究奠定了良好的基礎。
然而,在這些模型中存在大量難以確定的生理參數,不能直接應用于設施環境調控中。近年來,已有學者提出了光合速率的預測模型,但早期模型中沒有考慮多種環境因素與植物光合速率之間的耦合關系[13]。為此,學者們利用多元非線性回歸方法建立了以多環境因素為輸入的光合速率預測模型,提高了光合速率預測模型的準確性和通用性[14-15],但在多維光合數據的擬合上仍存在精度不足的問題。智能算法在光合速率預測模型上的應用有效提高了模型精度,成為新的研究熱點[16-17]。然而,現有的基于智能算法的光合速率預測模型大多沒有考慮根溫引起的水培蔬菜光合能力差異。
相關研究表明,水培蔬菜根溫將直接影響水培作物的根系生長和養分吸收[18-19]。HUANG 等[20]研究認為,隨著根溫的增加,斑茅的光合速率、根鮮質量、根數、氮、磷、鉀含量降低,從而加速根系死亡,影響根系生長速度和養分積累。李潤儒等[21]研究了不同根溫對水培環境中生菜生長和礦物質含量的影響;韓亞平等[22]探討了夏季高溫下不同根溫處理對番茄葉片氣孔的影響,發現根溫的增加會導致干旱脅迫。因此,針對水培作物在設施內根溫易調控以及根溫、氣溫、CO2 濃度和光照強度等環境變化時,作物最大光合速率差異性顯著的特點,構建根溫與環境耦合的光合速率預測模型,以提升模型精度。
本研究以生菜為試驗對象,設計了多因素嵌套試驗獲取試驗數據,利用神經網絡算法建立融合根溫的光合速率預測模型,以實現不同環境下的生菜葉片凈光合速率統一預測,旨在為設施農業水培生菜的環境控制奠定理論基礎。
1材料和方法
1.1 試驗材料
試驗選取奶油生菜作為研究對象。
1.2 試驗方法
試驗對生菜進行無土栽培,水溫17~22 ℃、pH值6.0~6.5,每隔2 d 更換一次營養液,不對其噴施農藥和激素。選擇其中茁壯的100 株作為試驗樣本,將其放置于MD1400 培養箱(荷蘭sinder 公司)內培養,待生菜幼苗生長至五葉一心時,試驗在第5 葉位完成數據測量。培養箱內光源設置為紅光(波長630 nm)和藍光(波長460 nm),相對濕度設定為60%,溫度為25 ℃,CO2體積分數為400 μL/L,光周期為晝14 h/夜10 h。
試驗使用LI-6800(美國LICOR 公司)便攜式光合速率儀測定幼苗期生菜的凈光合速率,設置光量子通量密度、葉溫、根溫、二氧化碳體積分數等不同的環境參量,進行多因子嵌套試驗。其中,根溫共設置6 個梯度:13、15、17、21、25、29 ℃ ;葉溫共設置4 個梯度:15、20、25、30 ℃ ;二氧化碳體積分數共設置3 個梯度:400、800、1 200 μL/L;光量子通量密度共設置9 個梯度:0、20、50、100、300、500、550、600、700 μmol/(m2 ·s)。為確保試驗數據穩定性并減少偶然性,每組試驗在相同條件下重復3 次,并取3 次試驗的均值作為該組試驗最終結果。基于以上試驗獲得的數據,作為適宜根溫區間的多因子耦合光合速率建模所需的樣本集,為光合速率建模提供數據基礎。
1.3 相關性分析
為探索4 個變量之間的關系,對試驗數據進行相關性分析,應用Pearson 分析法計算各因子與光合速率的相關系數。
1.4 融合根溫的光合速率模型的構建
由1.3 相關性分析得出,選取根溫、葉溫、二氧化碳體積分數、光量子通量密度作為輸入量,光合速率作為輸出量,采用3 層網絡結構的LMBP 算法建立光合速率預測模型。其建模過程主要包括3 個步驟,為了避免輸入和輸出數據量綱差異對模型預測效果的影響,首先利用比例放縮法完成數據歸一化,將輸入和輸出限制在[0,1]。
建模過程包括:第一,以歸一化處理后的數組X '1 (加入根溫的樣本集)作為輸入樣本集1,樣本集1 的 輸 入 層 節 點 數 為 4,其 輸 入 信 號 為 X1'=( X ' 1,X ' 2,X ' 3,X ' 4 ),以歸一化處理后的數組 X '2 (未加入根溫)作為輸入樣本集2,樣本集2 的輸入層節點數為 3,其輸入信號為 X2'=( X1',X2',X3' ),其中,X1'、X2'、X3'和X4'分別為根溫、葉溫、二氧化碳體積分數和光量子通量密度;光合速率T '為輸出樣本集。
第二,基于光合試驗數據采用試參法對模型參數進行優化、設計,以訓練誤差最小作為參數和隱節點數的選擇依據,完成模型結構的設計。
第三,基于預試驗結果進行網絡設計,并設定模型終止條件是均方根誤差<0.002 μmol/(m2·s),最大循環次數為1 000,從而在隨機數隨機分配權值閾值的基礎上,構建訓練網絡選取試驗數據進行網絡訓練。其流程如圖1 所示。
首先利用優化的網絡參數對網絡進行初始化,輸入一組處理好的樣本數據,觸發以下過程,利用LM 訓練法對網絡進行訓練。
在輸出層節點數為1 時,利用光合速率的真實值(d)與網絡輸出值(o),可以得到輸出層的誤差信號和隱含層誤差信號。
2結果與分析
2.1 相關性分析結果
各環境參量與植物光合速率的相關性如表1所示。
由表1 可知,根溫、葉溫、二氧化碳體積分數、光量子通量密度與光合速率均呈正相關,且均達到顯著水平(P<0.01)。根溫與光合速率的相關系數為0.078,在0.01 水平上顯著相關,相關性雖然不是最大的,但是根溫對光合速率的影響是不可忽視的,根溫也同樣是構建光合速率模型過程中不可或缺的因素之一。
2.2 參數優化結果
2.2.1 隱層節點數獲取結果 BP 網絡到目前為止還沒有確定的隱層節點數獲取方法,常根據經驗確定,這樣可能會對網絡效能產生較大的影響,隱層節點數過少則網絡誤差較大,而隱層節點數過多又會延長訓練時間,甚至會出現“過擬合”現象。針對不同節點訓練網絡模型,利用試驗測得的648 組數據進行BP 算法的隱層節點數試驗,獲取模型訓練集與測試集均方根誤差,并以該誤差作為選擇隱層節點數的指標。由于網絡參數初始化是隨機的,為避免初始化的偶然性結果影響模型性能,不同隱層節點數的網絡都進行100 次初始化并利用誤差反傳算法迭代500 次獲得100 個不同的結果。同一節點數訓練100 次后,訓練集和測試集均方根誤差取100 次的平均值,并作為評價隱層節點數的最終指標。隱層節點數與均方根誤差的關系如圖2、3 所示。
由圖2、3 可知,隨著隱層節點數的不斷增加,其訓練集誤差不斷縮小,但均方根誤差在隱層節點數為15 時已經趨于平穩,當隱層節點數再增加時對訓練集誤差變化很小,并且測試集均方根誤差在隱層節點數為15 時取得最小,為了兼顧訓練集和測試集誤差,選取15 作為隱層節點數。
2.2.2 傳遞函數結果分析 BP 神經網絡在建模過程中,激活函數必須是處處可微的,且是連續可微的,故BP 網絡經常使用的是S 型的對數或正切激活函數和線性函數。為了提高網絡的性能,進一步探索最優傳遞函數組合,就不同的傳遞函數組合進行試驗。試驗在網絡其他條件相同時,獲取不同隱含層和輸出層傳遞函數組合條件下模型均方根誤差和模型誤差穩定時的訓練步數,結果如表2 所示。
從表2 可以看出,序號7 隱含層傳遞函數為logsig,并且輸出層傳遞函數為purelin 時,其訓練結果均方根誤差最小,訓練步數相對較少,故選擇logsig 和purelin 的組合作為隱含層和輸出層傳遞函數。
2.2.3 訓練方法選取結果 BP 網絡不同的訓練方法將會對網絡收斂速度、誤差等性能產生影響。為了測試不同訓練函數的性能,在網絡其他條件相同時,利用不同訓練函數對2.2.2 得到的648 組數據進行網絡訓練,直至網絡均方根誤差趨于穩定,將不同訓練函數條件下的網絡模型均方根誤差結果展示如表3 所示。從表3 可以看出,2.2.2 訓練法中trainlm 訓練函數能有效減小均方根誤差,且能降低訓練步數,訓練性能最優,故采用trainlm 訓練法進行建模。
2.2.4 學習率確定 BP 算法的學習率也是模型性能的主要影響因素之一,學習率太小會導致訓練速度太慢;而學習率太大可能會使查找的步長太大而錯過網絡的最優解,使網絡性能大大降低。為獲取最佳模型訓練學習率,在網絡其他條件相同時,利用不同學習率訓練模型,記錄模型誤差穩定時的均方根誤差與模型訓練時間,結果如表4 所示。
從表4 可以看出,序號2 的學習率為0.10 時,其均方根誤差為0.287 873 0,與序號1 相比,在均方根誤差相近的基礎上縮短了整個算法執行時間,為了同時保證訓練結果的精確性與算法執行的快速性,本研究選擇的學習率為0.10。
2.3 模型結果對比與驗證
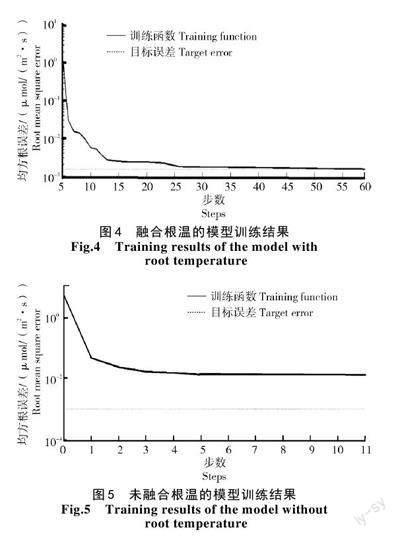
基于2.2.2 試驗獲得的樣本集,以LM 訓練法訓練網絡,目標均方根誤差為0.002,訓練最大步數為1 000 步,融合根溫對模型訓練結果如圖4、5所示。進一步對比分析可知,加入根溫的樣本集在訓練過程中均方根誤差逐漸逼近目標誤差值,并且在第45 步收斂,達到目標誤差值。此時模型訓練集均方根誤差為1.465×10-3 μmol/(m2·s),平均絕對誤差為1.873×10-3 μmol/(m2·s);未加入根溫的訓練結果在第5 步已經趨于平穩,誤差值不再縮小,并且與訓練目標值存在較大偏差。
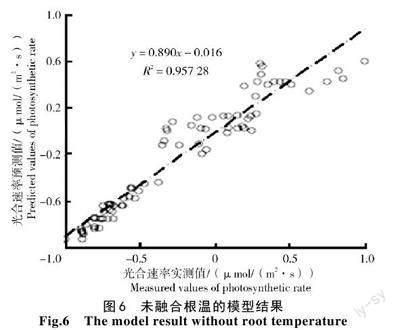
采用130 組數據組成的測試集對光合速率模型進行驗證,其光合速率的實測值和預測值之間的關系如圖6、7 所示。從圖6、7 可以看出,加入根溫后的模型斜率為1,截距為0.004 5;未加入根溫的模型斜率為0.890,截距為0.016。對比分析可知,加入根溫后的模型的預測值和實測值的擬合度更優。因此,融合根溫之后的BP 光合速率模型相較于未加入根溫的模型誤差更小,模型的預測性能更好。
3結論與討論
融合根溫后的模型更精準可能是由于水培作物根溫直接影響養分吸收,從而改變地上器官的光合作用特性[23]。蒸騰速率的增加是由于根溫的增加和根系流量的增加。它為PSⅡ反應中心提供了一個合適的環境,有助于光化學反應和電子轉移[24]。在適宜的條件下,凈光合速率大大提高。當根溫增加到一定程度時,蒸騰速率逐漸降低,葉表面氣孔關閉[25]。在這種情況下,由于作物不能進行充分的光合作用,光合速率逐漸停滯。此外,適宜的根溫有利于緩解地上器官的低溫脅迫,促進細胞分裂和葉面積的擴大,為葉綠素的產生提供必要的條件[26]。因此,以水培作物對根溫的響應為基礎,結合環境溫度、CO2 體積分數和光量子通量密度構造光合速率預測模型具有重要意義。當未考慮根溫條件時,同樣外界環境由于根溫差異,地上器官的光合能力存在差異,但模型只能預測同一光合速率,導致模型精度不足,與上述結果一致。
考慮到各環境因子對水培蔬菜光合反應的影響,利用試驗數據進行相關性分析表明,光合速率與根溫、葉溫、二氧化碳體積分數、光量子通量密度都在0.01 水平上顯著相關。在建立光合速率預測模型時需要充分考慮上述因素的變化,同時結合人工神經網絡算法,可實現水培生菜光合速率精準預測。模型預測值和光合速率真實值擬合結果表明,加入根溫因素后擬合直線斜率更接近1,斜率更接近0,且收斂誤差更低。本研究不只是針對水培奶油生菜的光合速率預測研究,而是一般性研究,也可適用于其他水培作物。為了使本模型更具有工程意義,未來還應考慮營養液配比、光配方等的影響。
參考文獻:
[1] 齊琦. 我國設施蔬菜種苗產業化繁育的現狀及發展[J]. 農業與
技術,2015,35(4):154.
QI Q. Present situation and development of industrialized breeding
of protected vegetable seedlings in China[J]. Agriculture and
Technology,2015,35(4):154.
[2] 秦玉芝,邢錚,鄒劍鋒,等. 持續弱光脅迫對馬鈴薯苗期生長和
光合特性的影響[J]. 中國農業科學,2014,47(3):537-545.
QIN Y Z,XING Z,ZOU J F,et al. Effects of sustained weak
light on seedling growth and photosynthetic characteristics of potato
seedlings[J]. Scientia Agricultura Sinica,2014,47(3):
537-545.
[3] 張玉鑫,常濤,曲亞英,等. 根區溫度對甜瓜幼苗光合特性的影
響[J]. 中國蔬菜,2009(20):50-54.
ZHANG Y X,CHANG T,QU Y Y,et al. Effects of root-zone
temperature on photosynthesis characteristics of Cucumis melo
L. seedlings[J]. China Vegetables,2009(20):50-54.
[4] 趙玉萍,鄒志榮,楊振超,等. 不同溫度和光照對溫室番茄光合
作用及果實品質的影響[J]. 西北農林科技大學學報(自然科學
版),2010,38(5):125-130.
ZHAO Y P,ZOU Z R,YANG Z C,et al. Effect of temperature
and light to tomato photosynthesis and quality in greenhouse[J].
Journal of Northwest A & F University(Natural Science Edition),
2010,38(5):125-130.
[5] 孫潛,崔世茂,宋陽,等. 內保溫日光溫室光溫性能及增施二氧
化碳對黃瓜生長發育的影響[J]. 北方園藝,2016(9):50-55.
SUN Q,CUI S M,SONG Y,et al. Impact of illumination and
temperature performance of blanket-inside solar greenhouse and
CO2 enrichment on cucumber growth and development[J].
Northern Horticulture,2016(9):50-55.
[6] HE J,AUSTIN P T,LEE S K. Effects of elevated root zone
CO2 and air temperature on photosynthetic gas exchange,nitrate
uptake,and total reduced nitrogen content in aeroponically
grown lettuce plants[J]. Journal of Experimental Botany,2010,
61(14):3959-3969.
[7] JIANG Y Q,LI T,ZHANG M,et al. WSN-based control system
of CO2 concentration in greenhouse[J]. Intelligent Automation
& Soft Computing,2015,21(3):285-294.
[8] 樊懷福,蔣衛杰,郭世榮. 低溫對番茄幼苗植株生長和葉片光
合作用的影響[J]. 江蘇農業科學,2005,33(3):89-91.
FAN H F,JIANG W J,GUO S R. Effects of low temperature
on plant growth and leaf photosynthesis of tomato seedlings[J].
Jiangsu Agricultural Sciences,2005,33(3):89-91.
[9] 宋敏麗,溫祥珍,李亞靈. 根際高溫對植物生長和代謝的影響
綜述[J]. 生態學雜志,2010,29(11):2258-2264.
SONG M L,WEN X Z,LI Y L. Effects of high rhizosphere
temperature on plant growth and metabolism:a review[J]. Chinese
Journal of Ecology,2010,29(11):2258-2264.
[10] 馮玉龍,劉恩舉,孫國斌. 根系溫度對植物的影響(Ⅰ)——根
溫對植物生長及光合作用的影響[J]. 東北林業大學學報,
1995,23(3):63-69.
FENG Y L,LIU E J,SUN G B. Influence of temperature of
root system on plant(i)—influence of root temperature on plant
growth and photosynthesis[J]. Journal of Northeast Forestry
Universiyt,1995,23(3):63-69.
[11] 李潤儒. 根區溫度對水培生菜生長與光合特性的影響[D]. 南
京:南京農業大學,2015.
LI R R. Effects of root zone temperature on growth and photosynthetic
characteristics of hydroponic lettuce[D]. Nanjing:Nanjing
Agricultural University, 2015.
[12] YE Z P,YU Q,KANG H J. Evaluation of photosynthetic electron
flow using simultaneous measurements of gas exchange
and chlorophyll fluorescence under photorespiratory conditions
[J]. Photosynthetica,2012,50(3):472-476.
[13] 吳鍇,張靜,郝稱意. 溫室番茄群體光合作用模擬模型的研究
[J]. 山西農業大學學報(自然科學版),2013,33(3):225-229.
WU K,ZHANG J,HAO C Y. Simulation of canopy photosynthesis
model of greenhouse tomato[J]. Journal of Shanxi Agricultural
University(Natural Science Edition),2013,33(3):
225-229.
[14] 李天來,顏阿丹,羅新蘭,等. 日光溫室番茄單葉凈光合速率
模型的溫度修正[J]. 農業工程學報,2010,26(9):274-279.
LI T L,YAN A D,LUO X L,et al. Temperature modified
model for single-leaf net photosynthetic rate of greenhouse tomato[
J]. Transactions of the Chinese Society of Agricultural
Engineering,2010,26(9):274-279.
[15] 胡瑾,何東健,任靜,等. 基于遺傳算法的番茄幼苗光合作用
優化調控模型[J]. 農業工程學報,2014,30(17):220-227.
HU J,HE D J,REN J,et al. Optimal regulation model of tomato
seedlings photosynthesis based on genetic algorithm[J].
Transactions of the Chinese Society of Agricultural Engineering,
2014,30(17):220-227.
[16] HU J,XIN P P,ZHANG S W,et al. Model for tomato photosynthetic
rate based on neural network with genetic algorithm
[J]. International Journal of Agricultural and Biological Engineering,
2019,12(1):179-185.
[17] WANG W,ZHANG M,LIU C,et al. Real-time monitoring of
environmental information and modeling of the photosynthetic
rate of tomato plants under greenhouse conditions[J]. Applied
Engineering in Agriculture,2013:783-792.
[18] 李思思,張紅梅,金海軍,等. 根際高溫對不同砧木黃瓜嫁接
幼苗生長、光合及生理特性的影響[J]. 中國農學通報,2018,
34(1):61-67.
LI S S,ZHANG H M,JIN H J,et al. Effects of rhizosphere
high temperature on growth,photosynthesis and biochemical indexes
of grafted cucumber seedlings on different rootstocks[J].
Chinese Agricultural Science Bulletin,2018,34(1):61-67.
[19] HE J,TAN L P,LEE S K. Root-zone temperature effects on
photosynthesis,14C-photoassimilate partitioning and growth of
temperate lettuce(Lactuca sativa cv‘. Panama) in the tropics
[J]. Photosynthetica,2009,47(1):95-103.
[20] HUANG B R,XU Q Z. Root growth and nutrient element status
of creeping bentgrass cultivars differing in heat tolerance as
influenced by supraoptimal shoot and root temperatures[J].
Journal of Plant Nutrition,2000,23(7):979-990.
[21] 李潤儒,朱月林,高垣美智子,等. 根區溫度對水培生菜生長
和礦質元素含量的影響[J]. 上海農業學報,2015,31(3):
48-52.
LI R R,ZHU Y L,GAO Y M Z Z. Effects of root zone tem ‐
perature on the growth and mineral elements content of
hydroponically-grown lettuce[J]. Acta Agriculturae Shanghai,
2015,31(3):48-52.
[22] 韓亞平,李安平,李亞靈,等. 不同根際溫度處理對番茄葉片
氣孔的影響[J]. 山西農業科學,2015,43(10):1234-1236.
HAN Y P,LI A P,LI Y L,et al. The influence of different
root-zone temperature treatment on tomato leaves stomata[J].
Journal of Shanxi Agricultural Sciences,2015,43(10):1234-
1236.
[23] HE J,LEE S K,DODD I C. Limitations to photosynthesis of
lettuce grown under tropical conditions:alleviation by root-zone
cooling[J]. Journal of Experimental Botany,2001,52:1323-
1330.
[24] WANG B,JEFFERS S N. Effects of cultural practices and
temperature on Fusarium root and crown rot of containergrown
hostas[J]. Plant Disease,2002,86(3):225-231.
[25] 范玉潔,姜慧敏,溫祥珍,等. 長期高溫與增施CO2 對番茄葉片
光合作用及淀粉含量的影響[J]. 山西農業科學,2022,50(1):
41-45.
FAN Y J,JIANG H M,WEN X Z,et al. Effects of long-term
elevated CO2 coupled high temperature on the photosynthesis
and content of starch of tomato leaves[J]. Journal of Shanxi Agricultural
Sciences,2022,50(1):41-45.
[26] 李云,石玉,張毅,等. 根區亞低溫下GA3 與不同水平養分配合
施用對黃瓜生長和養分吸收的影響[J]. 山西農業科學,2020,
48(9):1472-1476,1480.
LI Y,SHI Y,ZHANG Y,et al. Effects of GA3 and different levels
of nutrients combined application on growth and nutrient uptake
of cucumber under sub low temperature in root zone[J].
Journal of Shanxi Agricultural Sciences,2020,48(9):1472-
1476,1480.
