靈武長棗葉片阿拉伯半乳糖蛋白分布特征
2023-10-10 08:49:00陶珊珊章英才劉迎雪
干旱地區農業研究 2023年5期
關鍵詞:植物
陶珊珊,王 靜,章英才,白 琳,劉迎雪
(寧夏大學生命科學學院,寧夏 銀川 750021)
阿拉伯半乳糖蛋白(AGPs)為一類廣泛分布于植物各器官細胞中富含羥脯氨酸且高度糖基化的糖蛋白(HRGPs)家族,主要由阿拉伯糖和半乳糖構成多糖支鏈和蛋白質核心鏈組成結構復雜的大分子[1-3]。通過分子生物學等技術對植物細胞中非常重要的糖蛋白AGPs研究表明,AGPs在生殖發育過程,包括花粉發育、花粉管生長、自交不親和、大孢子母細胞形成,果實的成熟和衰老過程,以及根和莖等營養器官發生與發育等方面起著重要作用[2-6]。研究結果表明,寧夏枸杞多糖及果實的細胞壁多糖是阿拉伯半乳聚糖AGPs糖蛋白,是枸杞子中具有免疫調節功能的藥用有效成分[2-5];番茄LeAGP1過量表達導致植株的株高下降,分支增多,種子數量及大小減少,表明此AGPs參與了營養生長和生殖發育[7];過量表達黃瓜 (Cucumissativus)CsAGP1基因的煙草植株株高增加,說明AGPs促進了莖的生長[8];擬南芥AtAGP30對種子萌發和根的發育起重要的調控作用[9],AtFLA1促進了根的發育和細胞分化[10],AtFLA11、AtFLA12參與了次生細胞壁發育[11-13],與木質素形成同源的基因AtXYP1、AtXYP2,該類AGPs與維管束發育密切相關[14],AtAGP31參與維管束發育與逆境應答[15-16],AtAGP19基因在細胞分裂與生長、葉的發育以及生殖發育過程中起著重要的作用[17]。基于AGPs相關基因的表達模式證實了多種類型的AGPs基因在不同組織部位特異性表達,在植物營養器官和繁殖器官發育中起非常重要的作用,因此,探明AGPs在不同組織部位分布的特征尤其重要[2-3]。
AGPs在植物各器官中都有存在,利用AGPs的特異性抗體,成為研究不同植物中不同部位AGPs分布與特性的重要手段,在枸杞果實AGPs免疫定位[2-5]、低溫引起雛菊(Bellisperennis)胚珠和花藥發育過程中AGPs分布的變化[18]、以及利用AGPs單克隆抗體JIM8、JIM13、MAC207和LM2對擬南芥有性生殖過程中AGPs進行定位方面取得了較多的研究成果[2-3,19]。在營養器官方面,JIM4識別的AGPs與胡蘿卜根早期維管組織的發育相關,JIM14識別的AGPs與根次生加厚的篩管有關[20],而JIM13識別的AGPs定位在幼嫩的木質部和根冠[21],LM2、LM14識別的AGPs定位于野生型大麥根毛的細胞壁和細胞質中[22],由此說明,不同的單克隆抗體識別的AGPs在根組織發育中有其特定的分布,但對植物葉發育中AGPs分布的研究較少。利用針對性抗體對AGPs進行定位的免疫學研究方法,由于其較高的特異性,成為不同植物中不同部位AGPs分布研究的良好途徑[2-3]。
葉是植物進行光合作用和蒸騰作用的主要器官,也是植物對環境變化比較敏感的器官,其AGPs的分布直接影響到其功能。研究表明,棗葉中有豐富的黃酮類[23-24]、三萜類[25]、糖類和皂苷等[26],不同發育時期靈武長棗葉營養成分含量與果實營養成分和果實品質間存在明顯的相關性[26];AGPs參與了靈武長棗果實維管束發育過程的形態建成、細胞分裂和體積的增大,并為果實發育提供營養支持及保護[2,27],因此,葉AGPs分布有可能對果實品質的形成發揮著重要作用。
靈武長棗(ZiziphusjujubaMill cv. Lingwuchangzao)是鼠李科棗屬的鮮食棗品種,原產于寧夏靈武市,果實具有優良的藥用品質和食用品質。近年來,在靈武長棗果實研究方面取得了非常多的研究成果[2-3,27],而有關靈武長棗葉AGPs糖蛋白的分布,以及對果實品質的影響尚不明了。因此,本研究以果實不同發育時期靈武長棗葉為材料,應用免疫熒光定位技術,較系統研究不同時期葉中AGPs糖蛋白的分布特征,為進一步研究葉AGPs對靈武長棗生長發育及其在果實品質影響方面的功能奠定基礎。
1 材料與方法
1.1 試驗材料
以位于寧夏靈武市的寧夏紅棗工程技術研究中心試驗基地6年生靈武長棗的葉為供試材料,采用隨機設計,3次重復,每次重復選擇生長發育良好、樹勢適中、長勢相似、栽培管理水平一致的5~10株植株。在果實膨大前期(7月10日)、快速膨大期(8月9日)、著色期(9月8日)、完熟期(9月28日)分別設計4次試驗,每次試驗時間均設定于上午9∶00—11∶00進行,從試驗植株樹冠的東、西、南、北4個方位以及上、中、下、里、外各個方向選擇棗吊中部果柄附近的葉作為試驗對象[2-3,27]。
AGPs單克隆抗體JIM13購自美國喬治亞大學,堿性磷酸酶標記的山羊抗小鼠IgG二抗購自Sigma-Aldrich公司。
1.2 試驗方法
參照Bao等[5]、王瑩瑩[28]及林曼娜[29]的方法,略有改動。
1.2.1 石蠟切片制作 選取各不同發育時期的葉,用鋒利的刀片將葉片分割成含有3條主脈的約0.6 cm×1.0 cm的小塊,立即放入含有3.7%(v/v)甲醛的2F4固定液中,蓋緊瓶蓋后注射器抽氣至樣品下沉瓶底,室溫固定4 h。更換固定液3次,每次30 min。然后用PBS磷酸緩沖液(10 mmol·L-1,pH=7.0)清洗3次,每次15 min。樣品在4℃下用0.05% Toluidine blue染色15 min,在4℃下依次用30%、50%、70%、90%的乙醇梯度脫水,每次60 min,100%乙醇脫水3~4次,60 min·次-1。樣品用乙醇∶Steedman’s wax=1∶1在37℃恒溫箱中滲透過夜,次日更換乙醇∶Steedman’s wax=1∶3混合液,在37℃滲透2.5 h,再用純Steedman’s wax在37℃滲透3次,每次2 h。將樣品用純Steedman’s wax在牛皮紙盒中包埋至凝固。后期蠟塊的固著、整修、切片、貼片和展片等步驟參考王靜,章英才等的方法[2-3,27]。
1.2.2 糖蛋白免疫熒光定位 切片脫蠟、復水、PBS淋洗浸泡、封閉液處理等參考王靜,章英才等的方法[2-3,27]。然后用PBS(含1% BSA)以1∶100比例稀釋后的AGPs單克隆抗體JIM13于4 ℃下孵育過夜,次日用PBS淋洗浸泡3次,每次10 min,以去除多余抗體,再用PBS(含1% BSA)以1∶300比例稀釋后的堿性磷酸酶標記的山羊抗小鼠IgG二抗36℃黑暗條件下孵育1 h。標記后,PBS淋洗浸泡除去未標記的二抗,0.01% Toluidine blue染色去除植物本身的自發熒光,切片經PBS淋洗浸泡后封片。切片在OLYMPUS IX73熒光顯微鏡(Olympus,日本)和LEICA STELLARIS 5激光共聚焦顯微鏡(Leica,德國)下觀察并拍照。
2 結果與分析
2.1 膨大前期葉AGPs分布特征
膨大前期葉AGPs的分布特征見圖1。
(1)表皮:果實膨大前期葉上、下表皮均由一層排列緊密的細胞組成。上表皮細胞較大,近長方形,垂直于葉片方向的細胞壁較薄,抗體所識別的抗原熒光AGPs分布較少,而表皮細胞外切向壁較厚,抗體所識別的抗原熒光AGPs分布較多,形成了較厚的角質層;下表皮細胞較小,方形,抗體所識別的抗原熒光AGPs分布情況與上表皮相似,下表皮細胞外切向壁AGPs分布較多,同樣形成了較厚的角質層;氣孔僅分布于下表皮,孔下室較大,保衛細胞中分布著少量AGPs(圖1A~E)。
(2) 葉肉:葉肉由6~7層細胞組成,靠近上表皮的葉肉組織由3層垂直于上表皮的長柱狀柵欄組織細胞組成、排列很緊密,細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs;近下表皮的葉肉組織主要由 3~4層短柱狀的柵欄組織細胞組成,垂直于下表皮,排列也較緊密,細胞壁和細胞內部也密集分布著抗體所識別的抗原熒光AGPs(圖1A~E)。由此看來,葉肉是葉AGPs分布的主要部位,靈武長棗葉為典型的等面葉,具有較典型的旱生結構特征。
(3)葉脈:靈武長棗葉具有3條發達的主脈,中間主脈維管束的直徑較大,兩側主脈維管束直徑略小(圖1A,B,D,F)。主脈維管束由近軸面的木質部、形成層和遠軸面的韌皮部組成。主脈木質部發達,主要包括多列導管和薄壁組織,每列均由6~7個放射狀排列的導管組成,近形成層的導管直徑相對較大,在木質部導管和薄壁組織細胞的細胞壁上密集分布著大量抗體所識別的抗原熒光AGPs,在近形成層的木質部中AGPs尤其豐富;木質部和韌皮部之間的形成層由多層扁平狀細胞組成,細胞中分布著大量抗體所識別的抗原熒光AGPs;韌皮部由多層排列緊密的較小細胞組成,細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs。維管束外由一圈卵圓形的分泌細胞形成維管束鞘的結構,細胞中沒有抗體所識別的抗原熒光AGPs分布(圖1A,B,D,F)。
主脈的上表皮之下為2~3層大小不等的厚角組織,細胞壁分布著較多抗體所識別的抗原熒光AGPs,緊鄰其下為 4~5層圓形或卵圓形較大的薄壁細胞,細胞壁上也分布著較多抗體所識別的抗原熒光AGPs,其中分布有1~2個分泌道,內部沒有AGPs熒光分布;下表皮之上為3~4 層形態較小的厚角組織,細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs,緊鄰其上為6~7層圓形或卵圓形較大的薄壁細胞,細胞壁和細胞內部抗體所識別的抗原熒光AGPs略少,其中分布有3~4個分泌道,內部沒有AGPs熒光分布(圖1A,B,D,F)。
在上下表皮間的葉肉中間部位,分布著比較小的側脈和細脈,大部分葉脈被橫切,有時可見葉脈的縱切,葉脈維管束的結構較簡單(圖1C,E)。外面是一圈含分泌物的薄壁細胞組成的維管束鞘,細胞中沒有抗體所識別的抗原熒光AGPs分布,內部木質部和韌皮部細胞的細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs(圖1C,E)。
2.2 快速膨大期葉AGPs分布特征
(1)表皮:果實快速膨大期上表皮垂直于葉片方向的細胞壁較薄,抗體所識別的抗原熒光AGPs分布較少,表皮細胞外切向壁較厚,抗體所識別的抗原熒光AGPs分布較多,形成了較厚的角質層;下表皮抗體所識別的抗原熒光AGPs分布情況與上表皮相似,細胞外切向壁AGPs分布較多,同樣形成了較厚的角質層,氣孔保衛細胞中分布著少量AGPs(圖2A~F)。
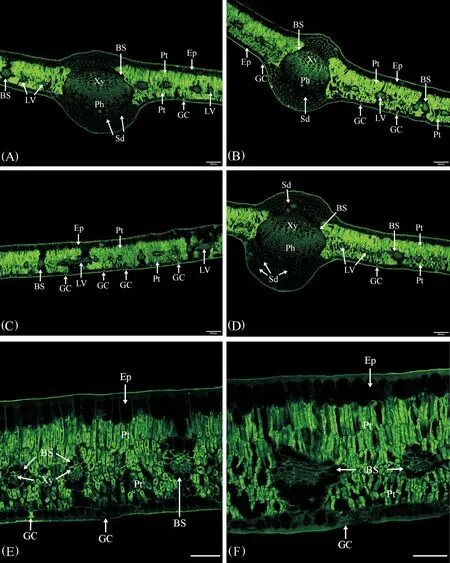
注:A~D為熒光顯微鏡圖片,Bar=50 μm; E和F為激光共聚焦顯微鏡圖片,Bar=50 μm。A、B:葉兩側主脈結構及葉AGPs分布;C、E、F:葉結構及AGPs分布;D:葉中間主脈結構及AGPs分布。Note:A~D are the pictures of fluorescence microscope, Bar=100 μm; E and F are the pictures of CLSM, Bar=50 μm. A, B:Bilateral main veins structure and AGPs distribution of leaf;C, E, F:Leaf blade structure and AGPs distribution;D:Middle main vein structure and AGPs distribution of leaf.圖2 快速膨大期葉AGPs免疫熒光分布Fig.2 Immunofluorescence distribution of AGPs in leaf during the rapid enlargement period
(2)葉肉:葉肉組織的細胞組成、細胞排列情況與果實膨大前期葉基本相似,葉肉柵欄組織細胞的細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs,AGPs分布特征也與膨大前期葉相同,葉肉柵欄組織是葉AGPs分布的主要部位(圖2A~F)。
(3)葉脈:中間和兩側主脈維管束在木質部導管和薄壁組織細胞的細胞壁上分布著大量抗體所識別的抗原熒光AGPs,形成層和韌皮部細胞壁和細胞內部分布著較多抗體所識別的抗原熒光AGPs,相比果實膨大前期有所減少(圖2A,B,D)。與果實膨大前期相似,維管束外由分泌細胞形成的維管束鞘細胞中基本沒有抗體所識別的抗原熒光AGPs分布(圖2A,B,D)。
主脈的上、下表皮之內的厚角組織細胞壁和細胞內部,以及緊鄰其內的圓形或卵圓形較大薄壁細胞的細胞壁和細胞內部均分布著抗體所識別的抗原綠色熒光AGPs,但相比膨大前期葉略有減少(圖2A,B,D);上、下表皮厚角組織內部薄壁細胞中分布的分泌道均沒有AGPs熒光(圖2A,B,D)。
上、下表皮間葉肉中間部位分布的側脈和細脈,不論是橫切還是縱切,維管束的結構均較簡單(圖2C,E,F)。與膨大前期葉相似,外面含分泌物的維管束鞘薄壁細胞中沒有抗體所識別的抗原熒光AGPs,內部木質部和韌皮部細胞的細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs(圖2C,E,F)。
2.3 著色期葉AGPs分布特征
(1)表皮:果實著色期葉上、下表皮細胞壁上抗體所識別的抗原熒光AGPs分布情況與快速膨大期相似,表皮細胞外切向壁較厚,抗體所識別的抗原熒光AGPs分布較多,形成了較厚的角質層(圖3A~F)。
(2)葉肉:與之前時期不同,果實著色期葉的葉肉組織細胞排列更加密集,即便是鄰近下表皮處的柵欄組織細胞排列也非常緊密,而且葉肉柵欄組織細胞的細胞壁和細胞內部均密集分布著大量抗體所識別的抗原熒光AGPs(圖3A~F)。
(3)葉脈:主脈維管束在木質部導管和薄壁組織細胞的細胞壁上密集分布著大量抗體所識別的抗原熒光AGPs,在近形成層的木質部中AGPs尤其豐富;形成層和韌皮部細胞壁和細胞內部分布著較多抗體所識別的抗原熒光AGPs(圖3A,B)。主脈的上下表皮之下的厚角組織、薄壁細胞及分布的分泌道AGPs分布特征與膨大前期葉相似(圖3A,B)。側脈和細脈維管束的結構均較簡單,內部木質部和韌皮部細胞的細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs(圖3C~F)。主脈、側脈和細脈維管束外的維管束鞘細胞中基本沒有抗體所識別的抗原熒光AGPs分布(圖3C~F)。
2.4 完熟期葉AGPs分布特征
(1)表皮:果實完熟期葉上、下表皮細胞壁上抗體所識別的抗原熒光AGPs分布情況與之前時期相似,表皮細胞外切向壁抗體所識別的抗原熒光AGPs分布較多,形成了較厚的角質層(圖4A~F)。
(2)葉肉:果實完熟期葉上、下表皮內的葉肉柵欄組織細胞與著色期相比排列更加緊密,柵欄組織細胞的細胞壁和細胞內部均密集分布著大量抗體所識別的抗原熒光AGPs(圖4A~F)。
(3)葉脈:與著色期相似,各主脈維管束在木質部導管和薄壁組織細胞的細胞壁上密集分布著大量抗體所識別的抗原熒光AGPs,在近形成層的木質部中AGPs同樣豐富;而韌皮部細胞壁和細胞內部分布著較多抗體所識別的抗原熒光AGPs(圖4A~B)。主脈厚角組織、薄壁細胞及分泌道AGPs分布特征與著色期相似(圖4A~B)。側脈和細脈維管束木質部和韌皮部細胞的細胞壁和細胞內部均密集分布著抗體所識別的抗原熒光AGPs(圖4C~F)。各葉脈維管束鞘細胞中均沒有抗體所識別的抗原熒光AGPs分布(圖4A~F)。
3 討論與結論
1)阿拉伯半乳聚糖蛋白(AGPs)是一類高度糖基化的蛋白質[2-3],作為細胞壁-質膜連續體的結構元素而存在,AGPs可能與植物適應逆境條件有關[18]。許多研究表明富羥脯氨酸糖蛋白(HRGPs)作為植物細胞壁的一種結構蛋白,在植物抗冷害機制中發揮著重要作用[2-3],HRGPs重要成員阿拉伯聚半乳糖蛋白AGPs與植物冷害存在重要相互關系,被證明在脅迫下參與信號傳導和細胞壁代謝[30]。棉花根部組織中的一個AGP基因GhAGP31在冷脅迫下表達量顯著上升[31],表明AGPs參與了植物的抗寒過程。另有研究表明,低溫可能導致“生理性干旱”脅迫的產生[32],低溫不僅使夏威夷椰子葉片葉肉細胞Ca2+水平及細胞超微結構等內部結構發生變化、光合作用及蒸騰作用顯著降低[33],而且AGPs的分布和表達也發生了變化。Yan等[34]采用免疫熒光標記技術研究了人工低溫脅迫下多種AGPs在香蕉幼苗葉片中的分布及變化規律;王瑩瑩[28]以香蕉葉片為供試材料,利用9種識別AGPs的單克隆抗體,采用免疫熒光技術研究了自然低溫脅迫下兩種香蕉葉片抗原的分布及差異表達,探討了AGPs在不同冷敏感程度香蕉葉片中的差異變化,不同抗體在兩種香蕉葉片中所識別的抗原位置及含量變化趨勢不同,揭示了AGPs與植物冷害的關系,認為香蕉中的AGPs基因MaFLA2-1、MaFLA17-3、MaFLA2-2的表達量可能與香蕉的抗冷性密切相關,表明這些 AGPs抗原也可能參與了香蕉的抗冷過程,揭示了AGPs與植物冷害的關系,豐富了植物抗冷機理,為香蕉抗冷育種工作提供參考。近年來發現,AGPs在植物應對生物及鹽脅迫等非生物脅迫過程中也扮演著重要角色[35-36],Lamport等[36]利用鹽脅迫分析AGPs功能認為,阿拉伯半乳聚糖蛋白(AGPs)通過未知機制參與細胞擴張,因此AGPs含量和細胞擴張速率可能存在相關性,鹽脅迫使煙草BY-2細胞中AGPs大量上調,AGPs通過高度多孔的果膠網絡擴散的速度大大提高。由此看來,AGPs在植物應對低溫、鹽堿脅迫導致的生理干旱等非生物脅迫過程中發揮了重要作用。
不僅如此,低溫等非生物脅迫還可能影響AGPs在植物體中的分布[30]。Leszczuk等[18]在低溫對雛菊(Bellisperennis)胚珠和花藥發育過程中AGPs分布變化影響研究中,利用識別AGPs碳水化合物鏈的特異性抗體(JIM13、JIM15、MAC207)免疫細胞化學研究表明,在雛菊發育的第一階段,AGPs在雌性和雄性生殖結構中的分布發生了明顯變化。通常情況下,AGPs具有特定持久的時空分布方式,AGPs抗原表位可見于每個形成階段的大孢子母細胞壁、大孢子和胚囊中,而在脅迫條件下的發育過程中,AGPs的定位發生改變,AGPs在胚囊壁中完全消失;在雄性部位發育的過程中,AGPs在正常情況下存在于絨氈層、小孢子母細胞和小孢子中,為了響應低溫發育,AGPs定位于小孢子的公共壁和成熟花粉粒中,此外,AGPs在殘余的絨氈層細胞中積累[18]。
寧夏是我國西北地區干旱半干旱荒漠與草原的過渡地帶,全區3/4以上屬干旱區。水分是限制該地區植物生長、產量和分布的主要因素之一,在所有的非生物限制因子中占首要地位[37]。葉與干旱環境的關系最密切,反應最敏感,環境不僅影響葉的外部形態,同時也影響著葉的內部構造和生理活動[38],其中葉結構特征最能體現植物對干旱環境的適應性[39]。靈武長棗是具有寧夏地方特色的藥食同源鮮果品種,抗寒、耐鹽堿、耐高溫和干旱[40]。作為干旱鹽堿地和沙荒地種植的先鋒樹種,靈武長棗葉演化出了具有較粗的葉脈,發達的機械組織,葉肉組織排列緊密,全部由多層柵欄組織組成,表皮有較厚的角質層,氣孔通常只分布在葉的下表皮等多種適應性結構,逐漸形成多種抗旱、耐旱的形態結構及對應的AGPs分布特征,在葉表皮細胞外壁、柵欄組織細胞、維管束木質部和韌皮部、機械組織等分布了大量的AGPs,反映了干旱條件下葉結構的適應性與各部位AGPs分布的相關性。從干旱缺水到營養器官和生殖器官的熱脅迫,AGPs被認為有助于植物應對生物和非生物脅迫的反應[41]。
2)AGPs糖蛋白在植物營養生長與生殖生長和發育過程中發揮著重要作用[2-3,42],但對植物葉發育中AGPs的研究較少。王瑩瑩[28]發現,抗體JIM4、JIM8、JIM14、JIM15和LM2等所識別的AGPs抗原在香蕉葉表皮、保衛細胞均有分布,既具有保護作用,又抵御了低溫脅迫。Domon等[43]發現,芒草(Miscanthus)在低溫脅迫下細胞壁中(1→3),(1→4)-β-D-glucan含量均表現上升,植物細胞壁成分的改變與植物抗寒性有著密切關系。植物在生長過程中受到生物脅迫或者非生物脅迫后,細胞壁的某些特性會發生極大改變,這些脅迫會改變細胞壁組分的含量及結構,從而改變細胞壁的機械性能,這種細胞壁機械性能的改變可以認為是植物對外界環境脅迫的一種響應[2-3,28]。本研究中,不同時期葉表皮的細胞壁及外壁中分布的AGPs,在干旱下誘發了表皮細胞壁的增厚和外壁中較厚角質的產生,減少了植物體內水分的過分蒸騰流失,使葉肉細胞更有效地利用水分,更好地發揮了保護及抵御干旱的功能。
旱生植物的葉肉向著提高光合效能、合成更多有機物以抵抗干旱的方面發展[39],靈武長棗葉高度發達的柵欄組織既避免了干旱地區強烈光照對葉肉細胞的損傷,又可以有效利用衍射光進行光合作用,即柵欄組織越厚、柵欄組織細胞越小且排列越緊密,則植物利用光能的效率越高[38]。干旱脅迫下,馬尾松針葉通過增加綠色折疊薄壁組織厚度維持較高的光合能力[44]。研究表明,低溫脅迫下,香蕉葉片組織細胞內的一些可溶物含量也在發生著改變,可溶性總糖、還原糖、甘油及可溶性蛋白質的含量均隨著溫度的降低而升高,并且淀粉與糖之間的轉化與植物的抗冷性有著密切關系[45]。煙草AGPs從引導組織向花粉管轉移,為花粉管的生長提供營養支持[46]。本研究表明,靈武長棗4個時期葉柵欄組織細胞的細胞壁和細胞內部都分布有大量JIM13抗體所識別的抗原,基本沒有明顯的變化,葉肉始終是葉AGPs分布的主要部位。葉肉柵欄組織細胞是植物進行光合作用的主要場所,柵欄組織細胞中大量的AGPs可能既參與了葉營養物質的合成,又作為營養物質來源,為植物的生長發育和后期果實營養物質的積累提供了保障和營養支持[2]。
3)細胞壁由纖維素、半纖維素、果膠質及AGPs等細胞壁結構蛋白共同構成[2,47],參與了植物細胞的生長發育、抗寒和抗旱等多種生命過程[2,28]。本研究表明,在靈武長棗4個時期葉主脈和側脈維管束木質部和韌皮部所有細胞的細胞壁和細胞內部都分布有JIM13抗體所識別的抗原,基本沒有明顯的變化。AGPs可能參與了木質部和韌皮部纖維素等細胞壁物質的沉積,以及木質部導管的分化和細胞壁的進一步增厚等過程[2-3,27,48]。靈武長棗葉脈維管組織中分布的AGPs參與了葉脈維管束發育過程的形態建成,為干旱條件下水分和養分的高效運輸提供了保障[2,27]。另外,王瑩瑩[28]發現多種不同抗體識別的維管束中AGPs抗原主要分布在香蕉葉脈的厚壁細胞和維管束鞘、木質部、韌皮部,而本研究結果略有不同,不論靈武長棗葉的主脈和側脈,在維管束鞘細胞中基本上都沒有AGPs的分布,可能與不同植物種類的葉結構差異有關,有可能還與維管束鞘中所含不同成分相關。AGPs不僅是細胞壁多糖的組分,而且可能參與了靈武長棗葉維管束鞘細胞中分泌物的形成和積累,既提高了細胞液濃度,增大滲透勢,又利于保水抗旱。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
