高溫脅迫下陸地棉GhCIPK6轉(zhuǎn)基因后代生理生化分析
2023-10-11 02:02:06郭金成張庭軍何良榮
新疆農(nóng)業(yè)科學(xué) 2023年9期
關(guān)鍵詞:植物
王 輝,郭金成,宋 佳,張庭軍,何良榮
(1.塔里木大學(xué)農(nóng)學(xué)院,新疆阿拉爾 843300;2.新疆生產(chǎn)建設(shè)兵團(tuán)第六師農(nóng)業(yè)科學(xué)研究所,新疆五家渠 831300)
0 引 言
【研究意義】高溫脅迫造成作物的經(jīng)濟(jì)損失越來越嚴(yán)重[1]。預(yù)計全球氣溫每10年將上升0.2℃[2]。植物的生長和發(fā)育過程受到高溫(HT)脅迫的嚴(yán)重影響[3,4]。高溫會導(dǎo)致作物的光合作用等生理活動受到嚴(yán)重影響,并影響植株的正常生長和發(fā)育,是作物減產(chǎn)的主要影響因素[5]。【前人研究進(jìn)展】在高溫脅迫下,一般先受到破壞的是細(xì)胞質(zhì)膜的結(jié)構(gòu)和功能[6]。高溫影響細(xì)胞膜的穩(wěn)定性,氣體交換(光合作用、光呼吸作用、蒸騰作用、氣孔導(dǎo)度)及相關(guān)酶的活性[7]。植物細(xì)胞在高溫逆境下利用多種方式產(chǎn)生活性氧造成氧化脅迫,但一定程度的高溫脅迫往往會促使產(chǎn)生較高水平的耐熱性,但當(dāng)高溫脅迫超出一定范圍后,活性氧生成速率加快,與此同時非酶性清除劑含量不斷減少、活性氧酶性消除系統(tǒng)活性也在不斷降低,這樣就導(dǎo)致大量活性氧不能被有效清除,進(jìn)而造成氧化傷害[8]。【本研究切入點】植物體內(nèi)的存在的一類微量植物激素,如脫落酸(Abscisic acid,ABA)、生長素(Auxin,IAA),水楊酸(Salicylic acid,SA)、茉莉酸(Jasmonic acid,JA)等。植物受到逆境脅迫時,體內(nèi)的激素發(fā)生變化,通過影響植物的生理生化活動來產(chǎn)生抵御脅迫的能力[9]。需開展關(guān)于植物對熱脅迫的響應(yīng)和防御方面的研究,了解陸地棉在生理生化水平上對高溫脅迫的響應(yīng)機理和抵御機制。【擬解決的關(guān)鍵問題】選擇轉(zhuǎn)基因材料超表達(dá)和、RNAi和野生型材料,測定3種材料測定不同時間、不同環(huán)境下生理生化指標(biāo)變化,為研究陸地棉抗高溫分子機理提供理論基礎(chǔ)。
1 材料與方法
1.1 材 料
材料為陸地棉品系YZ1,由華中農(nóng)業(yè)大學(xué)育種實驗室構(gòu)建GHCIPK6超表達(dá)和RNAi載體,遺傳轉(zhuǎn)化棉花品系YZ1,分別得到超表達(dá)和干涉的轉(zhuǎn)基因純系。RN代表正常溫度RNA干涉材料,RH代表高溫RNA干涉材料;WN代表正常溫度野生型材料,WH代表高溫野生型材料;ON代表正常溫度超表達(dá)材料,OH代表高溫超表達(dá)材料。
1.2 方 法
1.2.1 試驗設(shè)計
將棉花材料每株系種植90株以上,培養(yǎng)室中培養(yǎng),培養(yǎng)條件設(shè)定為相對濕度RH=80%,光周期14 h/10 h 白天/夜晚,光密度500 μmol/(m2·S),全程充分供水。待植株長大到第6周,將棉花植株轉(zhuǎn)入生長箱中。對照組生長條件保持不變,處理組42℃熱脅迫24 h,在26℃恢復(fù)24 h,處理及對照組分別于4、12、24和48 h取樣,每個時間點分2個生物學(xué)重復(fù),每個重復(fù)5株幼苗。
1.2.2 測定指標(biāo)
1.2.2.1 光合特性
使用美國LI-COR公司生產(chǎn)的Li-6400XT便攜式光合作用測量系統(tǒng)測定正常溫度和高溫下不同時間段棉花幼苗的凈光合速率(Pn)、氣孔導(dǎo)度(Gs)、蒸騰速率(Tr)、細(xì)胞間CO2濃度(Ci),使用SPAD測定儀(SPAD-502PLUS)測定葉綠素相對含量。
1.2.2.2 抗性相關(guān)
丙二醛(MDA)含量測定采用硫代巴比妥酸(TBA)比色法[10],過氧化物酶(POD)活性測定采用愈創(chuàng)木酚顯色法,可溶性蛋白含量的測定采用考馬斯亮藍(lán)染色法,脯氨酸(Pro)的測定方法采用茚三酮顯色法[11],超氧化物歧化酶(SOD)活性測定采用氮藍(lán)四唑(NBT)比色法[12]。
1.2.2.3 植物激素
激素測定所用的儀器為液質(zhì)聯(lián)用系統(tǒng)4000 Q-TRAR LC-MS(Applied Biosystems,USA)
1.3 數(shù)據(jù)處理
數(shù)據(jù)分析用WPS office Excel軟件,采用統(tǒng)計軟件DPS進(jìn)行單因素方差分析及數(shù)據(jù)顯著性檢驗和SPSS 21分析數(shù)據(jù)。
采用主成分分析對試驗中各個時期的14個相關(guān)生理指標(biāo)平均值進(jìn)行綜合評價。根據(jù)各主成分的得分,及各主成分的方差貢獻(xiàn)率為權(quán)數(shù),對所提取的得分進(jìn)行加權(quán)求和,得到各材料高溫下的綜合得分。
2 結(jié)果與分析
2.1 高溫脅迫下陸地棉幼苗葉片比較
研究表明,4 h時,在正常溫度26℃下,3種材料幼苗葉片正常生長,無任何變化,在高溫42℃下,超表達(dá)第1片真葉和第2片真葉明顯下垂枯萎,第3片真葉也出現(xiàn)下垂,RNAi和野生型第1片真葉和第2片真葉下垂不明顯,第3片真葉無變化。在48 h時,取下對照溫度和高溫的葉片,進(jìn)行比較,超表達(dá)葉片已經(jīng)發(fā)黃,葉片上已出現(xiàn)大量斑點,野生型葉片發(fā)黃程度次之,RNAi葉片只輕微發(fā)黃。圖1

注:A、B分別代表4 h正長溫度和高溫幼苗,C代表48 h幼苗葉片
2.2 高溫脅迫下陸地棉光合特性
2.2.1 高溫脅迫下陸地棉凈光合速率(Pn)變化
研究表明,光合作用是棉花生長發(fā)育必要條件,也是增加產(chǎn)量的必要因素,而凈光合速率是光合作用的最直接體現(xiàn)。在4 h,野生型進(jìn)行光合速率快,而超表達(dá)材料較慢;在12 h,三者均在正常溫度下進(jìn)行光合速率快,高溫下較慢;在24 h,三者在正常溫度下進(jìn)行光合速率大于三者高溫下,其中超表達(dá)高溫光合速率能力最低,在48 h,野生型正常溫度和高溫下不顯著,RNAi在高溫下與正常溫度下達(dá)到顯著,超表達(dá)在正常溫度和高溫下光合速率能力低。超表達(dá)相對其他兩種材料,光合速率較慢。圖2
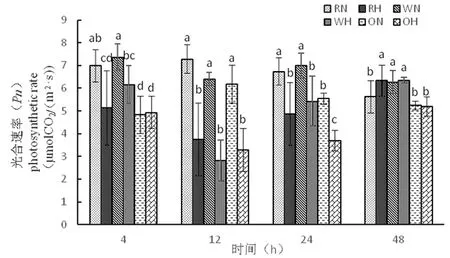
注:不同小寫字母表示差異顯著(P<0.05),下同
2.2.2 高溫脅迫下陸地棉氣孔導(dǎo)度(Gs)的變化
研究表明,氣孔的保衛(wèi)細(xì)胞受到外界環(huán)境的刺激能迅速做出反應(yīng),在高溫脅迫下,植物氣孔的保衛(wèi)細(xì)胞產(chǎn)生應(yīng)激反應(yīng)打開氣孔,CO2進(jìn)入植物的量增多,從而導(dǎo)致葉片光合速率增加。超表達(dá)材料在整個時期氣孔導(dǎo)度能力很低,在12 h時,3種材料氣孔導(dǎo)度在高溫下達(dá)到最低,氣孔關(guān)閉,隨著脅迫時間延長,氣孔導(dǎo)度逐漸增大,在脅迫超過一定時間,氣孔打開,光合速率增加,光合作用增強。超表達(dá)材料相對其他兩種材料,氣孔導(dǎo)度減弱,超表達(dá)材料進(jìn)行光合作用能力差。圖3
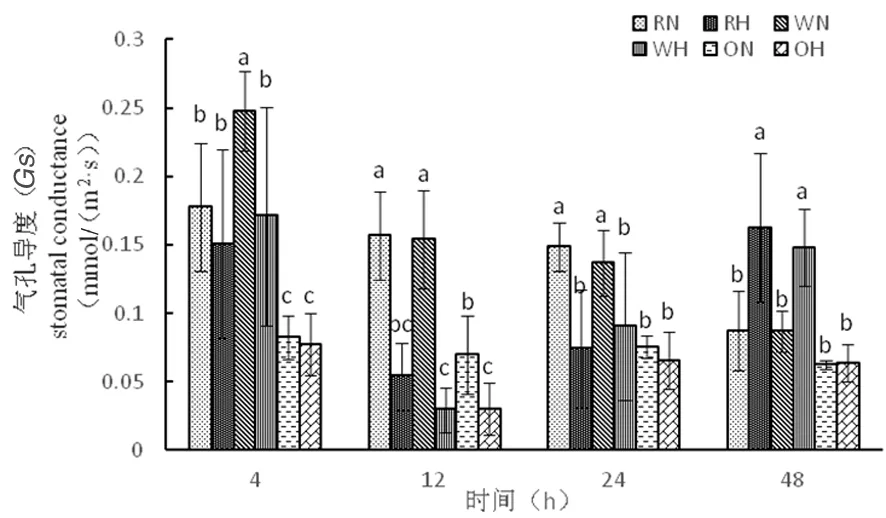
圖3 高溫脅迫下氣孔導(dǎo)度的變化
2.2.3 高溫脅迫下陸地棉蒸騰速率(Tr)的變化
研究表明,蒸騰作用能夠?qū)ν饨绛h(huán)境做出反應(yīng)來進(jìn)行自我保護(hù),在一定溫度下,蒸騰作用可降低葉片溫度,降低水分散失,增強對礦物質(zhì)的吸收。在正常溫度下12 h時,RNAi株系的蒸騰速率為1.33 mmolH2O/(m2·s),野生型株系蒸騰速率為1.14 mmolH2O/(m2·s),而高溫下,RNAi蒸騰速率為0.64 mmolH2O/(m2·s),野生型株系蒸騰速率為0.35 mmolH2O/(m2·s);48 h時,高溫處理下各材料蒸騰速率加快,RNAi為1.64 mmolH2O/(m2·s),野生型為1.42 mmolH2O/(m2·s),正常溫度下蒸騰速率減慢,RNAi為0.84 mmolH2O/(m2·s),野生型為0.68 mmolH2O/(m2·s)。高溫處理下,隨著時間延長,蒸騰速率增加,可以降低葉片溫度,減少水分散失,增強對礦物質(zhì)的吸收。超表達(dá)株系從4 h到48 h先降低在增加,在整個時期蒸騰速率緩慢。圖4
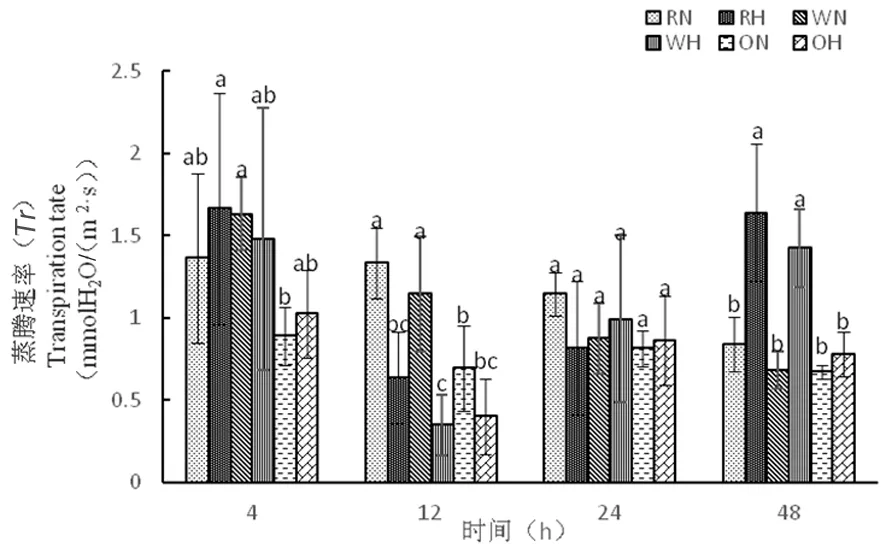
圖4 高溫脅迫下蒸騰速率的變化
2.2.4 高溫脅迫下陸地棉細(xì)胞間CO2濃度(Ci)的變化
研究表明,從4 h到12 h時,RH由303 μmol/mol下降到298 μmol/mol,WH由280 μmol/mol下降到262 μmol/mol,RNAi干擾株系和野生型株系胞間CO2濃度在高溫下并沒有急劇下降,氣孔導(dǎo)度下降,凈光合速率下降的主要因素并不是氣孔閉合,而超表達(dá)株系(4 h到12 h時)在高溫下急劇下降,OH由248 μmol/mol下降到199 μmol/mol,高溫下超表達(dá)株系敏感性強。圖5
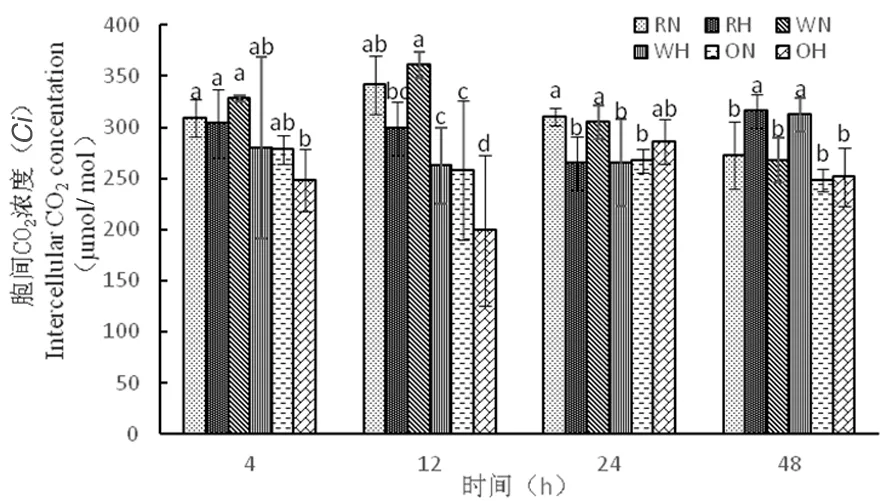
圖5 高溫脅迫下胞間CO2濃度的變化
2.2.5 高溫脅迫下陸地棉葉綠素相對含量(SPAD)的變化
研究表明,隨著時間延長,3種株系的葉綠素含量變化較小,其中RH葉綠素含量為30.5、30.1、30.3、34.1,WH葉綠素含量為29.5、30.3、30.3、32.9,OH葉綠素含量為28.2、27.8、27.6、29.3,在高溫脅迫下,SPAD值不會受溫度影響仍能保持高含量,從而進(jìn)行光合作用。圖6
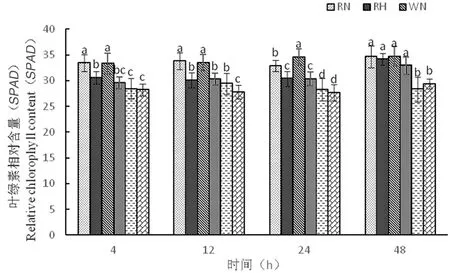
圖6 高溫脅迫下葉綠素相對含量的變化
2.3 高溫脅迫下陸地棉抗性
2.3.1 高溫脅迫下陸地棉丙二醛(MDA)的變化
研究表明,在正常溫度下,3種株系隨著時間延長,丙二醛含量呈先增加再降低后增加。在高溫下,隨著時間延長,RNAi干擾株系中丙二醛含量呈先增加后下降,野生型株系中丙二醛含量呈逐漸緩慢增加,而超表達(dá)株系丙二醛含量逐漸增加,其中48 h時達(dá)到最大為0.06 μmol/g,超表達(dá)株系在高溫脅迫下對膜造成的傷害大,對棉花細(xì)胞活力起到抑制作用。圖7
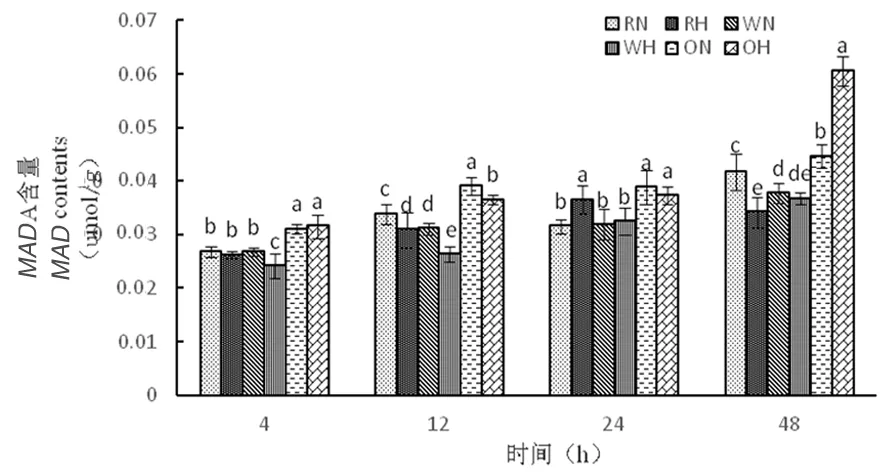
圖7 高溫脅迫下MDA含量的變化
2.3.2 高溫脅迫下陸地棉氧化物酶(SOD)、超氧化物歧化酶(POD)的變化
研究表明,在正常溫度下,RNAi干擾株系和野生型株系SOD活性相對較高,超表達(dá)株系中SOD活性先降低后增加。在高溫下,隨著時間延長,RNAi干擾株系中SOD活性呈先增加后下降,野生型株系和超表達(dá)株系中SOD活性呈先降低再上升后降低,超表達(dá)株系在整個時期都相對較低,分別為3.1、1.8、2.9、2.3 μ/mg,在正常溫度下,RNAi干擾株系和野生型株系中SOD活性對于清除自由基能力較強保護(hù)膜質(zhì)過氧化,在高溫下,超表達(dá)株系中SOD活性對于清除自由基能力較差,未能減輕細(xì)胞膜的損壞。圖8

圖8 高溫脅迫下SOD活性的變化
在正常溫度下,3種株系POD活性隨時間延長逐漸增加,其中超表達(dá)株系POD活性在整個時期含量較高。在高溫下,RNAi干擾株系和野生型株系隨著時間延長,SOD活性趨于平衡,而在超表達(dá)株系在整個時期都達(dá)到最大值,分別為222、239、199、220 μ/(mg·FW),超表達(dá)株系通過提高POD活性來應(yīng)對熱脅迫。受到高溫脅迫,在整個時期超表達(dá)株系SOD活性較低,而POD活性較高,在應(yīng)對高溫環(huán)境SOD、POD表現(xiàn)出不同的變化,發(fā)揮自身功能,應(yīng)對高溫脅迫表現(xiàn)出不同的作用。圖9
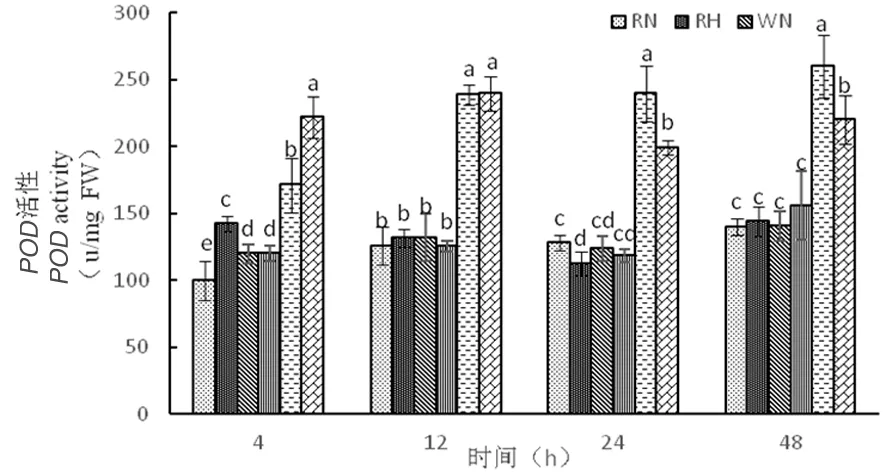
圖9 高溫脅迫下POD活性的變化
2.3.3 高溫脅迫下陸地棉可溶性蛋白的變化
研究表明,3種株系在正常溫度下可溶性蛋白含量相對較低,在48 h達(dá)到了最低,RN為0.71 mg/g,WN為0.88 mg/g,ON為1.38 mg/g。而在高溫下,隨著時間延長,RNAi干擾株系中可溶性蛋白含量呈先降低后增加,分別為5.87、3.95、2.87、4.49 mg/g,野生型株系可溶性蛋白含量先增加再降低后增加,分別為5.02、7.28、2.40、6.70 mg/g,超表達(dá)株系中可溶性蛋白含量呈先增加后降低,分別為4.99、5.15、6.30、5.68 mg/g,在高溫誘導(dǎo)下,可溶性蛋白含量較高。圖10

圖10 高溫脅迫下可溶性蛋白含量的變化
2.3.4 高溫脅迫下陸地棉脯氨酸(Pro)的變化
研究表明,在正常溫度下,3種株系的脯氨酸含量隨著時間延長逐漸增加,而在高溫下,野生型株系和超表達(dá)株系脯氨酸含量從4~12 h緩慢上升,12~48 h下降,RNAi干擾株系的脯氨酸含量隨著時間逐漸降低,超表達(dá)株系在整個時期脯氨酸含量相對較高,分別為34.32、36.39、34.91、30.97 μg/g,熱脅迫對超表達(dá)株系造成嚴(yán)重傷害,從而通過脯氨酸含量的積累應(yīng)對逆境。圖11
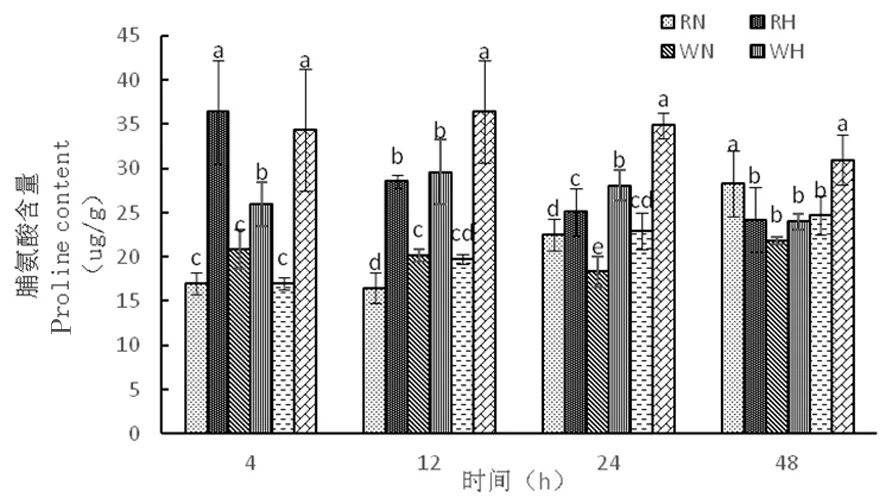
圖11 高溫脅迫下脯氨酸含量的變化
2.4 高溫脅迫下陸地棉植物激素
2.4.1 高溫脅迫下陸地棉生長素(IAA)的變化
研究表明,在正常溫度下,RNAi干擾株系和野生型株系隨著時間延長生長素含量趨于平衡,超表達(dá)株系中生長素含量逐漸上升。在高溫下,RNAi干擾株系中生長素含量呈先下降后上升,變化為7.59、7.02、9.20、10.31 ng/g,野生型株系W中生長素含量呈先上升后下降,變化為6.77、7.06、10.53、7.86 ng/g,超表達(dá)株系OE中生長素含量逐漸升高,變化為6.83、8.19、9.28、9.70 ng/g。隨著時間延長,生長素不受高溫影響,能夠抵御高溫環(huán)境。RNAi干擾株系和超表達(dá)株系隨著時間延長體內(nèi)生長素含量逐漸增加。圖12
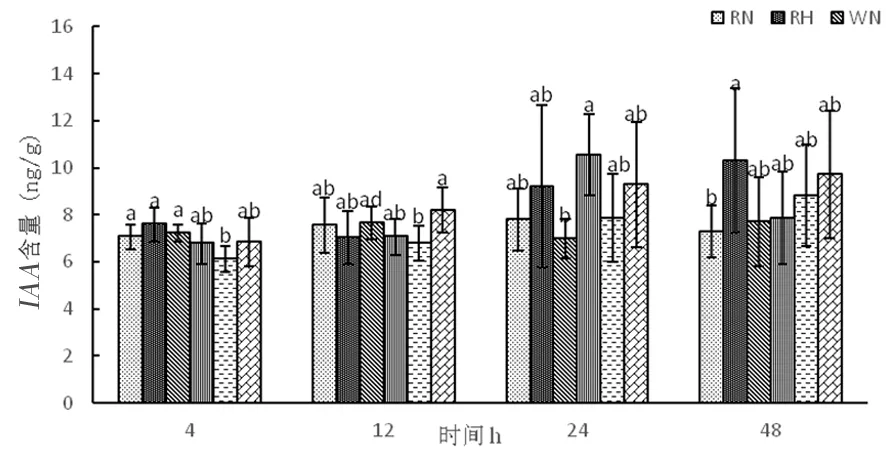
圖12 高溫脅迫下生長素含量的變化
2.4.2 高溫脅迫下陸地棉脫落酸(ABA)的變化
研究表明,正常溫度下,3種株系在整個時期脫落酸含量呈現(xiàn)先增加在下降。高溫下,超表達(dá)株系和野生型株系隨著時間延長,脫落酸含量先增加再下降后上升,野生型株系上升的幅度較大,超表達(dá)株系則上升幅度較小,而RNAi干擾株系脫落酸含量先增加后下降。超表達(dá)株系在整個時期脫落酸含量相對較低,常溫處理4個時間段分別為84.80、148.68、94.45、126.64 ng/g,而高溫處理時分別為165.71、195.34、147.47、162.46。高溫脅迫時,超表達(dá)株系OE中的脫落酸因未能迅速做出反應(yīng)來提高植物抗性。圖13
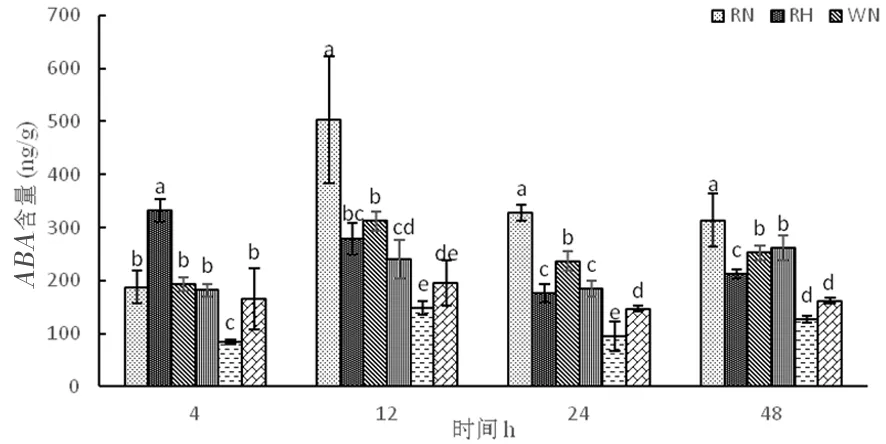
圖13 高溫脅迫下脫落酸含量的變化
2.4.3 高溫脅迫下陸地棉茉莉酸(JA)的變化
研究表明,正常溫度下隨著時間延長,野生型株系中茉莉酸含量呈先增加后下降,RNAi干擾株系中茉莉酸含量逐漸增加,超表達(dá)株系茉莉酸含量先增加再下降后上升。高溫下,3種株系隨著時間延長體內(nèi)茉莉酸含量呈現(xiàn)先增加后下降。圖14
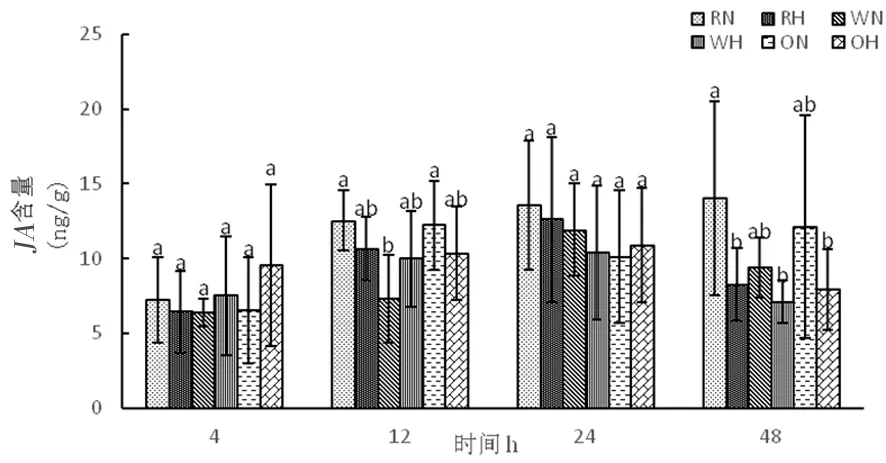
圖14 高溫脅迫下茉莉酸含量的變化
2.4.4 高溫脅迫下陸地棉水楊酸(SA)的變化
研究表明,正常溫度下,3種株系隨著時間延長水楊酸含量呈先上升再下降后上升,在整個時期,超表達(dá)株系OE中水楊酸變化幅度較為明顯,分別為22.56、94.16、31.54、37.60 ng/g,其中在12 h達(dá)到最大值。高溫下,RNAi干擾株系和野生型株系中水楊酸含量呈先下降再上升后下降,RNAi分別為19.10、12.41、53.33、23.24 ng/g,野生型W分別為18.50、13.91、37.93、19.10 ng/g,超表達(dá)株系高溫脅迫中水楊酸含量呈先上升后下降,分別為33.19、40.51、48.42、20.21 ng/g。高溫脅迫前期,超表達(dá)株系可以通過增加水楊酸含量來實現(xiàn)對植物抗逆保護(hù),但48 h下降為20.21 ng/g,保護(hù)作用下降。圖15
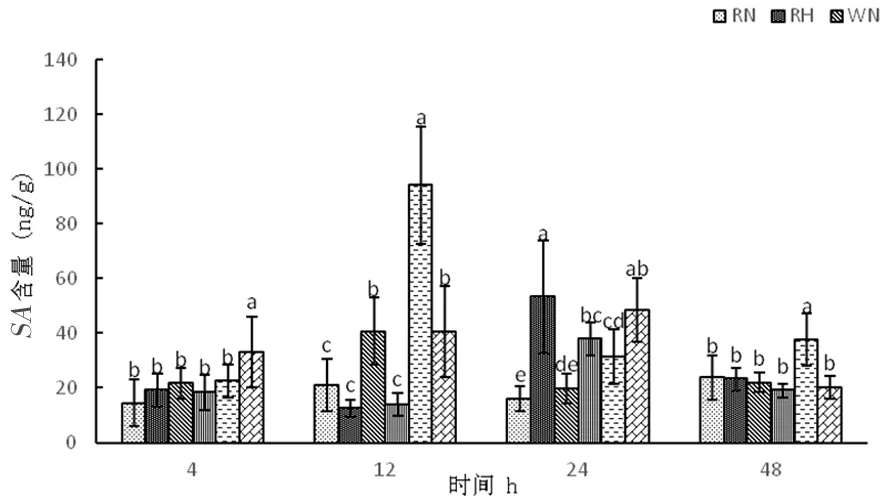
圖15 高溫脅迫下水楊酸含量的變化
2.5 高溫脅迫下陸地棉綜合評價
研究表明,得到各植物種測定指標(biāo)成分的特征值、貢獻(xiàn)率和累計貢獻(xiàn)率,得到4個主成分可以解釋全部方差的84.587%,提取前4個主成分可以代表14個高溫下指標(biāo)信息的84.94%。在第1主成分分析中,光合作用指標(biāo)的貢獻(xiàn)率較大,其中蒸騰速率最大,過氧化物酶最低;第2主成分分析中,生長素的貢獻(xiàn)率較大,水楊酸次之;在第3主成分分析中,丙二醛含量的貢獻(xiàn)率最大;第4主成分分析中,脯氨酸含量的貢獻(xiàn)率最大。蒸騰速率可視為反映植物抗高溫性能的最重要生理生化指標(biāo),其次是生長素含量、丙二醛含量和脯氨酸含量。表1
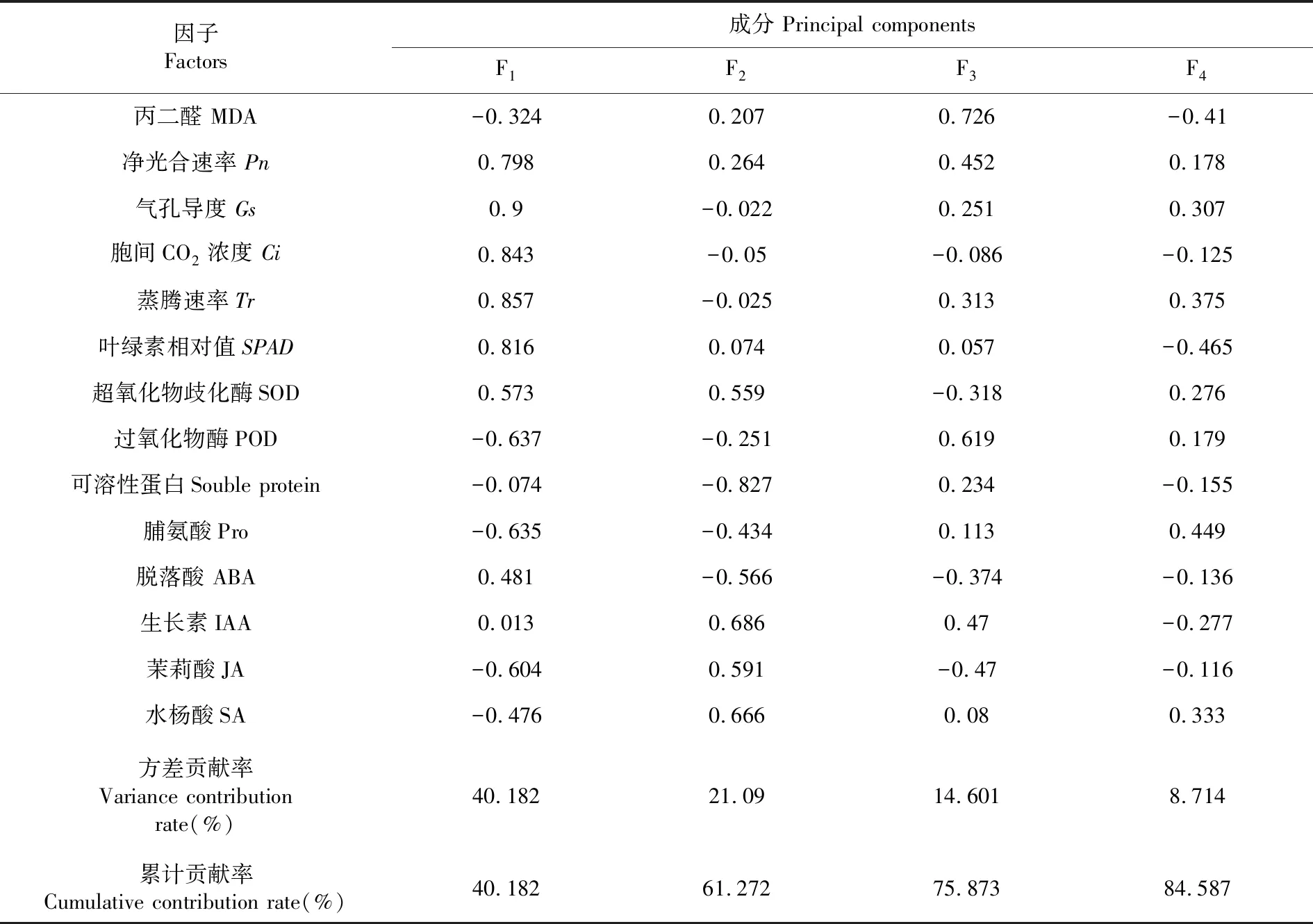
表1 主成分因子載荷矩陣及主成分特征根
排序排在前3的是RNAi中的48 h,野生型的48 h和4 h,3種材料抗高溫能力由強到弱為:RNAi,野生型,超表達(dá),RNAi綜合得分為0.201,野生型綜合得分為0.135,超表達(dá)綜合得分為-0.337。表2

表2 主成分得分及綜合評價
3 討 論
3.1 高溫脅迫對陸地棉光合特性的影響
氣孔是植物與環(huán)境進(jìn)行物質(zhì)交換的通道,氣孔通過調(diào)節(jié)其開閉狀態(tài),控制著CO2進(jìn)入植物體內(nèi)及水分從體內(nèi)的散失[13]。可溶性蛋白具有滲透調(diào)節(jié)的作用,還能夠在一定程度防止細(xì)胞質(zhì)脫水[14]。脯氨酸具有調(diào)節(jié)滲透及保護(hù)細(xì)胞結(jié)構(gòu)穩(wěn)定的作用[15]。脫落酸具有促進(jìn)植物休眠、抑制生長、促進(jìn)氣孔關(guān)閉和增加抗逆的作用[16]。茉莉酸作為一種重要的新型植物內(nèi)源激素,通過信號傳遞來調(diào)控植物防御反應(yīng)和抗逆發(fā)揮著重要作用[17]。水楊酸作為植物產(chǎn)生系統(tǒng)獲得性抗性的內(nèi)源信號, 當(dāng)植物受到非生物因素影響時, 植物體局部水楊酸含量增加,促進(jìn)抗逆蛋白合成、提高滲透調(diào)節(jié)能力和抗氧化能力,并伴有一些防衛(wèi)相關(guān)蛋白的表達(dá)[18]。光合作用是一切生物直接或間接的能量來源,是植物進(jìn)行物質(zhì)轉(zhuǎn)換和能量代謝的關(guān)鍵[19]。引起植物凈光合速率下降的原因主要分為氣孔因素和非氣孔因素,氣孔限制因素引起的Pn下降表現(xiàn)為Gs、Ci、Tr的下降,非氣孔限制因素引起的Pn下降表現(xiàn)為Tr、Gs下降的同時Ci升高[20,21]。研究中隨著時間延長凈光合速率Pn呈先下降后上升的同時氣孔導(dǎo)度Gs、蒸騰速率Tr、細(xì)胞間CO2濃度Ci均也呈先下降后上升,Pn的變化是由氣孔限制因素所導(dǎo)致的,與竇飛飛[22]結(jié)論一致。
3.2 高溫脅迫對陸地棉光抗性相關(guān)的影響
在高溫下,RNAi氧化物酶呈先上升后下降,而超表達(dá)和野生型材料先下降再上升后下降,隨著時間延長,體內(nèi)的氧化物酶都在下降,可能導(dǎo)致酶活性下降,也有相關(guān)研究在馬鈴薯和大白菜中發(fā)現(xiàn)同樣的規(guī)律[23,24]。超氧化歧化酶活性與氧化物酶活性不同,RNAi和野生型材料在高溫下大致趨于平衡,可能由于超氧化歧化酶活性不受環(huán)境影響,在高溫下對植物進(jìn)行保護(hù),而超表達(dá)材料超氧化歧化酶活性異常顯著,敏感性材料會大量產(chǎn)生超氧化歧化酶。3種材料隨著時間延長,體內(nèi)丙二醛含量逐漸增加,其中超表達(dá)材料在48 h丙二醛含量達(dá)到最大值,且不同材料耐熱性存在顯著差異,與王倩[25]研究結(jié)論相似。脯氨酸具有阻止細(xì)胞膜透性的作用,在高溫脅迫下,隨著時間延長,脯氨酸含量逐漸降低,與前人研究結(jié)果相反,隨著時間延長,脯氨酸調(diào)節(jié)細(xì)胞滲透的功能下降,抗熱脅迫能力減弱。在溫度脅迫下,植物體內(nèi)通過提高可溶性蛋白含量來增強本身的抗逆性[26],研究3種材料在高溫下,可溶性蛋白含量在整個時期相對較高,可溶性蛋白遇到高溫誘導(dǎo)產(chǎn)生熱激蛋白來增加植物的的抗逆性。
3.3 高溫脅迫對陸地棉植物激素的影響
植物內(nèi)源激素在植物應(yīng)對環(huán)境變化時起重要作用,內(nèi)源激素水平、合成和分布會受溫度的影響而發(fā)生改變,多種植物內(nèi)源激素參與植物應(yīng)對逆境脅迫時的信號轉(zhuǎn)導(dǎo)[27]。在高溫下,RNAi和超表達(dá)材料生長素含量相對呈上升趨勢,提升植物耐受性,與王日明[28]研究結(jié)果相似,野生型材料生長素含量隨著時間延長呈現(xiàn)先上升后下降,說明野生型抗熱性差。楊長琴等[29]研究結(jié)果顯示高溫脅迫下棉花葉片中ABA含量隨著時間的延長呈先升后降的趨勢,研究在高溫下,超表達(dá)和野生型材料隨著時間延長脫落酸含量呈先上升再下降后上升,RNAi材料隨著時間延長,脫落酸含量先上升后下降,植物不一定總是通過提高ABA水平來抵抗高溫脅迫。隨著高溫脅迫時間的延長,3種材料茉莉酸含量先升后下降,與宋悅[30]研究結(jié)果相似。RNAi和野生型材料在高溫下,隨著時間延長水楊酸含量呈先下降再上升后下降,超表達(dá)隨著時間延長先上升后下降,可能由于前期水楊酸促進(jìn)抗逆蛋白合成來增加抗逆性,但隨著時間延長,植物需求水楊酸逐漸增大,致使水楊酸含量逐漸降低。
3.4 高溫脅迫下陸地棉綜合評價
植物響應(yīng)高溫是由體內(nèi)各種機制共同制約環(huán),僅憑一個指標(biāo)來評價植物耐熱性是有很大的局限性[31]。研究結(jié)果表明,RNAi耐熱性最強,其次野生型,超表達(dá)耐熱性最差。
4 結(jié) 論
3種材料抗高溫能力由強到弱為RNAi,野生型材料,超表達(dá)材料。在高溫下可通過提高蒸騰速率來減弱逆境脅迫的危害,提高生長素含量來積極調(diào)控植物生長,提高細(xì)胞的滲透調(diào)節(jié)能力來減輕細(xì)胞遭受的損傷。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
