不同造林密度馬尾松人工林分化特征及其對生產力的影響*
2023-11-05 12:59:10顏培棟楊章旗黃綏理周永斌零天旺
林業科學 2023年10期
顏培棟 李 鵬 楊章旗 黃綏理 周永斌 零天旺
(1. 廣西壯族自治區林業科學研究院 國家林業和草原局馬尾松工程技術研究中心 廣西馬尾松工程技術研究中心 廣西優良用材林資源培育重點實驗室 南寧 530002;2. 廣西橫州市鎮龍林場 橫州 530327)
林木個體大小分化對森林生態系統的空間結構、群落演替、種間(內)關系以及林分結構和生產力水平均具有顯著影響(Ali,2019;Levicket al.,2015),尤其在人工純林生態系統中,如何減少林分內個體分化大小(Tschiederet al.,2012),對培育中大徑材林分(譚健暉,2021)、提高林分生產力水平尤為重要。林木個體大小分化在空間結構上受光照、土壤水分和無機養分等供給能力的影響,林木對生長資源的獲取和利用效率存在差別,進而發生生態位分化引起個體大小差異(Boydenet al.,2008;Schumeet al.,2004)。在時間上,林分生長早期階段種間和種內競爭較弱(Oheimbet al.,2011),隨著林齡增加,林木樹冠開始互相接觸遮蔽,根系依靠自身趨水和趨肥特性(Zhanget al.,2006),向相鄰個體水肥條件較好的地方延伸,個體間競爭加劇,該過程中處于優勢地位的個體占有更多生長資源,相比劣勢個體更容易在林分中脫穎而出(Binkleyet al.,2010),從而導致林分內個體大小不斷分化。當林分內個體競爭達到一定水平時,林分會發生自然稀疏(Duanet al.,2019),劣勢被壓木枯死,為優勢木提供生長空間和養分,尤其是高密度種植時,因獲取資源的非對稱性競爭發生年齡更早(Stankovaet al.,2017;諶紅輝等,2004)。林木個體大小分化直接導致林木徑階結構改變,林木徑階結構分布特征指林木個體按徑階大小的分布狀況,是反映林分結構的重要指標(Yanget al.,2002),在人工林純林發育過程中,探討林木個體大小分化及其徑階分布特征對提高林分生產力和培育中大徑材林木具有重要意義。
馬尾松(Pinus massoniana)是我國亞熱帶地區特有的鄉土樹種和南方荒山造林先鋒樹種,也是重要的工業原料林樹種,具有速生、豐產、適生性強、用途廣等優點(孟祥江等,2018)。目前,關于馬尾松人工林經營技術方面已開展施肥效應(胡炳堂等,2000)和密度調控(王建軍等,2019)等研究,并取得了顯著成果;然而在馬尾松人工林經營過程中仍存在密集種植、采伐過早等問題,既不利于大徑材培育,也造成林分生產力的浪費(李婷婷等,2015)。如何改進經營方法,培育中大徑材馬尾松人工林,提高林分生產力,尤其是在充分了解不同造林密度馬尾松人工林生長發育過程的基礎上,對馬尾松人工林個體分化和徑階分布特征及其對林分生產力影響的探討尤為重要。鑒于此,本研究基于南亞熱帶地區不同造林密度樣地長期監測結果,分析4 種林分密度馬尾松人工林個體大小分化、徑階分布特征及其對林分生產力的影響,闡明林齡、造林密度和自然稀疏過程對林木個體大小分化和生產力的影響規律,以期為確定適宜經營密度、培育馬尾松林中大徑材、提高林分生產力提供依據。
1 試驗地概況與研究方法
1.1 試驗地概況
試驗林位于鎮龍林場那歪分場4 林班10 小班,地處廣西橫州市境內(109°08′—109°19′E,23°02′—23°08′N),屬南亞熱帶季風氣候區,降雨充沛,年均降雨量1 477 mm,年均蒸發量1 056 mm,年均溫度21.5 ℃,極端低溫-1 ℃,極端高溫39.2 ℃,年均日照時數1 758.9 h,無霜期336 天。試驗地海拔450 m,坡度25°,坡向西南216°,中上坡位;土壤為紫色砂頁巖發育而成的赤紅壤,平均土層厚60 cm;林下灌草主要有山雞椒(Litsea cubeba)、華南毛柃(Eurya ciliata)、桃金娘(Rhodomyrtus tomentosa)、山烏桕(Sapium discolor)、三叉苦(Euodia lepta)、鴨腳木(Schefflera octophylla)、黃毛榕(Ficus esquiroliana)、 大青(Clerodendrum cyrtophyllum)、棕葉蘆(Thysanolaena maxima)、烏毛蕨(Blechnum orientale)和五節芒(Miscanthus floridulu)等。
1.2 樣地設置與調查
試驗林于1997 年5 月采用馬尾松優良家系(桂MVF443)半年生營養杯苗(平均苗高25 cm、地徑0.26 cm)造林,隨機區組設計,設置3 個區組(即3 次重復),2 500(A)、3 333(B)、4 500(C)、6 000 株·hm-2(D)4 種造林密度,株行距分別為2 m×2 m、1.5 m×2 m、1.5 m×1.5 m、1 m×1.67 m。每個小區面積30 m×30 m,造林后前2 年進行常規割草撫育,其后采用近自然經營措施。每種密度選擇坡度、坡向、土壤等立地因子基本一致的3 塊20 m×20 m 標準地(樣地間隔>50 m),共布設12 塊固定樣地(范志偉等,2012)。每年測定林木樹高(tree height,H)和胸徑(diameter at breast height,DBH)等生長指標,并記錄林木存活狀況。2006—2020 年連續15 年,每年年底進行定期測定,樣地基本概況如表1 所示。
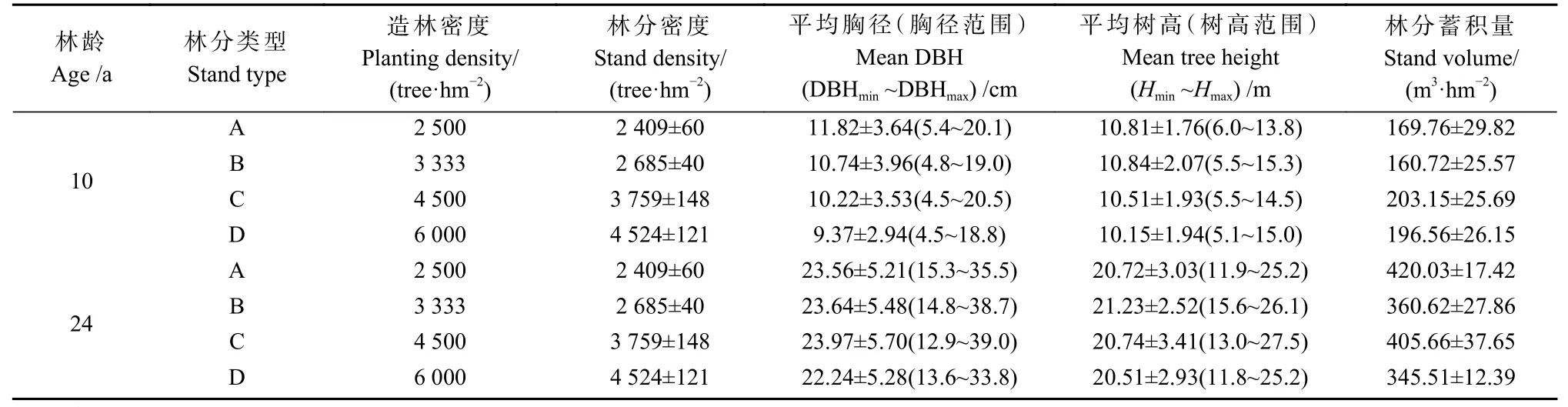
表1 樣地基本概況①Tab. 1 Basic characteristics of the sampling plots
1.3 數據分析與計算方法
1.3.1 林分生產力計算 根據林分每木檢尺結果,馬尾松人工林活立木單株材積采用二元材積公式V=7.142 654 37×10-4×DBH1.867008H0.9014632計算,其中:V為單株材積,m3;DBH 為胸徑,cm;H為樹高,m。單位面積活立木總材積為活立木單株材積累加除以樣地面積。林分生產力采用林分活立木總材積的平均年生長量(stock volume annual increment,VAI,m3·hm-2a-1)表示。
1.3.2 林木個體分化程度表征 參考楊桂娟等(2019)方法,采用基尼(Gini)系數表征林分內林木個體大小分化程度。Gini 系數最早由Lorenz 在1905 年提出(Lorenz,1905),用于社會科學中分析人群收入的差異程度,后被引入表征植物種群內個體大小的差異程度,并被廣泛應用。本研究中Gini 系數采用分組計算法,其計算公式為:
式中:n為樣地內活立木株數;Yi為活立木單株斷面積。
1.3.3 林分自然稀疏效應 林分自然稀疏程度(selfthinning rate)采用林木當年死亡數量除以總株數乘以100%表示。為確定林分自然稀疏過程中死亡單株對林分整體水平的影響,采用每年林分自然稀疏的個體平均斷面積與林分死亡發生前林分整體平均斷面積(mean basal area,BA)的比值(BAdeath: BAtotal)反映,BAdeath: BAtotal>1 表示死亡單株平均大小大于林分整體水平,BAdeath: BAtotal<1 表示死亡單株平均大小小于林分整體水平。
1.3.4 徑階分布劃分 按徑階將馬尾松分為大徑材(胸徑在26 cm 徑階及以上)、中徑材(胸徑在18~24 cm徑階之間)和小徑材(胸徑在16 cm 徑階及以下),以2 cm 為1 個徑階。
采用Excel 2016 和SPSS 20.0 軟件對數據進行統計分析,所用數據均以均值±標準差表示。運用重復測量方差分析(repeated measure ANOVE)檢驗造林密度和林齡對馬尾松人工林個體大小分化、徑階分布特征和林分生產力影響的差異,差異顯著性多重比較采用Tukey’s 檢驗(n=3),顯著性水平α=0.05。利用R 語言的Random Forest 程序包進行隨機森林分析,判定林齡、造林密度和自然稀疏過程對林木個體大小分化和林分生產力的影響。應用Smart PLS 軟件構建偏最小二乘法-結構方程進行Gini 系數和林分生產力的影響路徑分析。
2 結果與分析
2.1 馬尾松人工林個體大小分化特征
Gini 系數隨林齡增加顯著減小(F=20.598,P<0.001),個體間分化程度隨造林密度增加顯著增大(F=6.172,P<0.05),林齡與造林密度的交互作用對Gini 系數無顯著影響(圖1)。不同造林密度下Gini 系數與林齡間呈極顯著線性負相關(P<0.001)。
由圖2 可知,林分自然稀疏過程中,隨林齡增加活立木株數顯著減少(F=324.168,P<0.001),由造林時的2 500~6 000 株·hm-2降至10 年生時的2 344~4 790 株·hm-2,并逐漸趨于一致,到24 年生時為588~1 396 株·hm-2。造林密度越大,林分內存活株數減小越快,減少數量越多,隨自然稀疏進程持續趨于一致,到17 年生后不同造林密度活立木株數間無顯著差異,林齡與造林密度的交互作用對活立木株數具有顯著影響(F=8.596,P<0.001)。不同造林密度下活立木株數與林齡呈極顯著的指數函數變化趨勢(P<0.001)。隨林齡增加林分自然稀疏率存在顯著差異(F=4.389,P<0.01),造林后頭9 年,A(2 500 株·hm-2)密度自然稀疏率最小為3.65%,其次為C(4 500 株·hm-2)密度(16.20%),而B(3 333 株·hm-2)和D 密度(6 000 株·hm-2)自然稀疏率較大,分別為19.44%和24.60%,從整體來看,林分自然稀疏率較大的林齡為13~14 年和20 年生時,表明在這2 個時間點林分內個體競爭程度較大,競爭劇烈。不同造林密度間林分自然稀疏率存在顯著差異,高密度自然稀疏率大于低密度,而林齡與造林密度的交互作用對林分自然稀疏率無顯著影響。
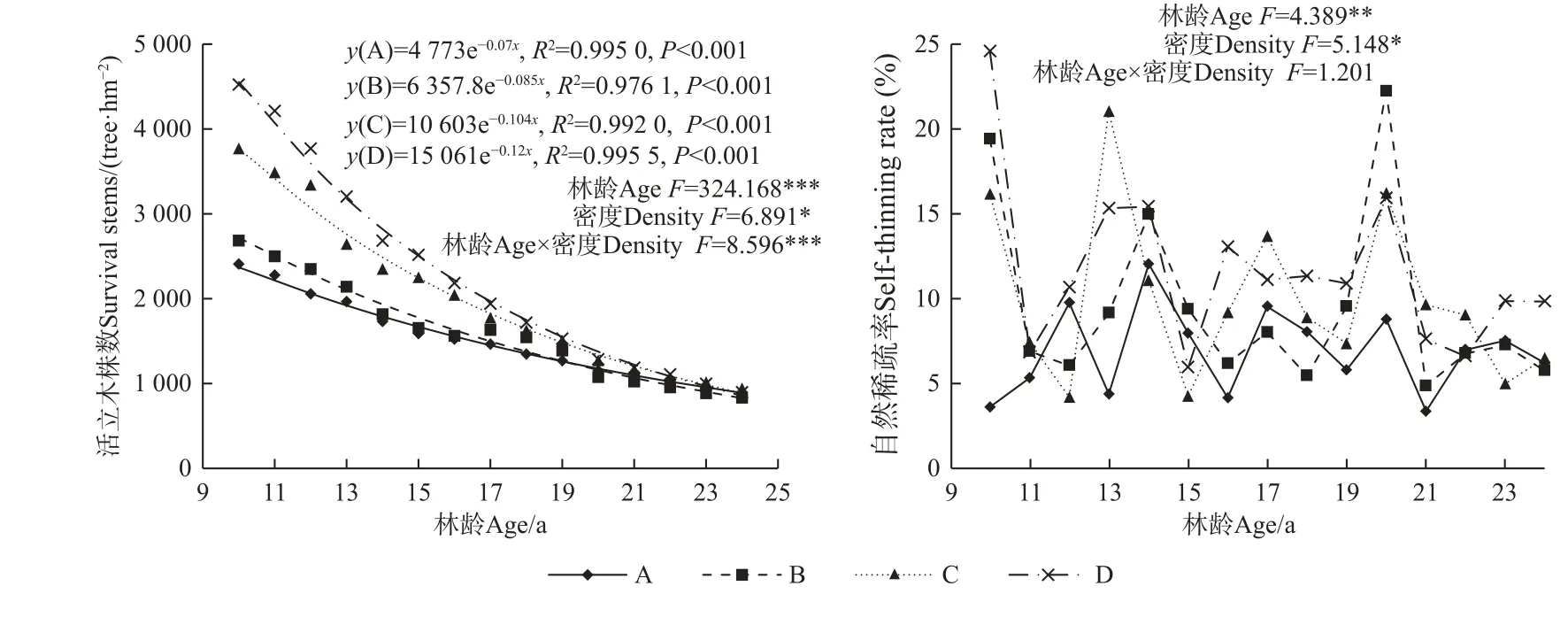
圖2 馬尾松人工林活立木數量和自然稀疏率的變化趨勢Fig. 2 Dynamics of surviving stems and self-thinning rate of P. massoniana plantations
活立木株數與Gini 系數間呈對數函數分布(P<0.001),且隨活立木株數增加Gini 系數增大,并逐漸趨于水平,而自然稀疏率與Gini 系數間無顯著相關(圖3)。
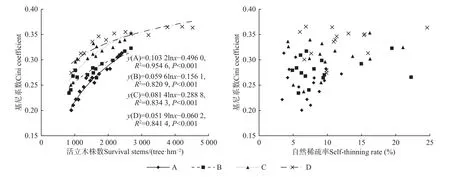
圖3 馬尾松人工林活立木數量和自然稀疏率與Gini 系數的關系Fig. 3 The relationship between the surviving stems and self-thinning rate and Gini coefficient of P. massoniana plantations
林分自然稀疏過程中,林分死亡個體斷面積與林分平均斷面積的比值(BAdeath: BAtotal)隨林齡增加顯著增大(F=2.668,P<0.05)(圖4),整體呈上升趨勢,且在19 年生后均有出現>1 的情況,表明林分自然稀疏過程中,隨林齡增加死亡單株平均大小大于林分整體水平,死亡單株對林分整體的影響隨林齡增加而加大。不同造林密度下BAdeath: BAtotal具有顯著差異(F=3.018,P<0.01),隨造林密度增加BAdeath: BAtotal增大,林齡和造林密度的交互作用對BAdeath: BAtotal無顯著影響。不同造林密度下BAdeath: BAtotal與林齡間均呈冪函數分布特征(P<0.01)。
2.2 馬尾松人工林徑階分布特征
馬尾松人工林大、中和小徑材株數隨林齡增加均具有顯著性差異(P<0.01)(圖5),其中大徑材株數隨林齡增加呈近似“S”形變化,14 年開始出現大徑材,14~17 年間大徑材株數增加較慢,18~22 年間大徑材株數增加最快,23 年后逐漸平穩;22~24 年間不同造林密度大徑材株數具有顯著差異(P<0.05),均呈現C密度顯著高于其他密度。中徑材株數隨林齡增加呈先增加后減少趨勢,均在17~19 年間出現極大值;10~19 年間馬尾松人工林中徑材株數均在A 密度下最多,表明A 密度適用于中徑材馬尾松速生豐產林培育,且主伐年齡在19 年后適宜。小徑材株數隨林齡增加呈顯著減少趨勢,10~17 年間均呈現D 密度顯著高于他密度,而隨林齡增加不同造林密度馬尾松人工林小徑材株數逐漸減小并趨于一致。徑階分布特征三元相圖表明,馬尾松人工林大徑材株數占比在25%及以上包括A 密度在21~24 年、B 和C 密度均在22~24 年、D 密度在24 年;中徑材株數占比在50%及以上包括A 密度在17~24 年、B 密度在20~24 年、C 密度在24 年、D 密度在23 年;小徑材株數占比在50%以下包括A 密度在17~24 年、B 密度在19~24 年、C 和D密度均在20~24 年。
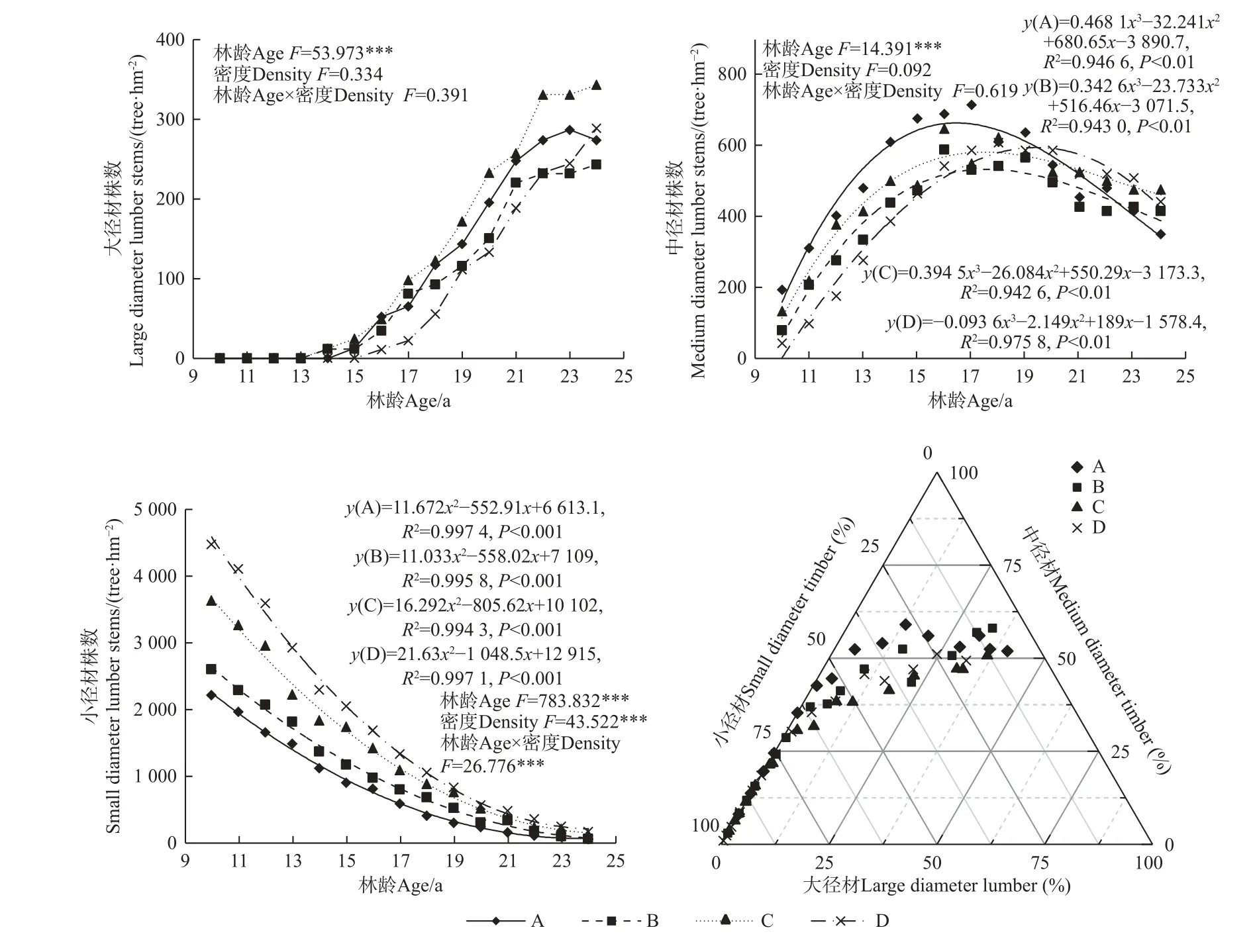
圖5 不同造林密度馬尾松人工林大、中和小徑材株數變化及分布特征Fig. 5 Changes and distribution characteristics of large, medium and small-diameter trees with different densities of P. massoniana plantations
2.3 馬尾松人工林林分生產力
重復測量方差分析(圖6)表明,馬尾松人工林林分蓄積量和林分生產力隨林齡變化均存在極顯著差異(P<0.001),造林密度、林齡與造林密度的交互作用對林分蓄積量和林分生產力無顯著影響。不同造林密度馬尾松人工林林分蓄積量隨林齡增加呈對數函數增加,其中A 密度下林分蓄積量變化幅度最大,從9 年生的183 m3·hm-2至24 年生的420.03 m3·hm-2,15年間增加237.03 m3·hm-2,其次為C 和B 密度,分別增加202.51 和186.76 m3·hm-2,D 密度下林分蓄積量增加幅度最小,為148.95 m3·hm-2。不同造林密度馬尾松人工林林分生產力隨林齡增加整體呈先增加后減小并逐漸趨于穩定,其中A 和B 密度在13 年生時最大,分別為19.31 和18.76 m3·hm-2,C 和D 密度均在11 年生時最大,分別為20.48 和19.67 m3·hm-2。不同造林密度林分生產力的大小順序在不同林齡階段呈現不同變化趨勢,其中10~11 年林分生產力為C > D > A > B,12~15 年林分生產力為C > A > B > D,16~24 年林分生產力為A > C > B > D。
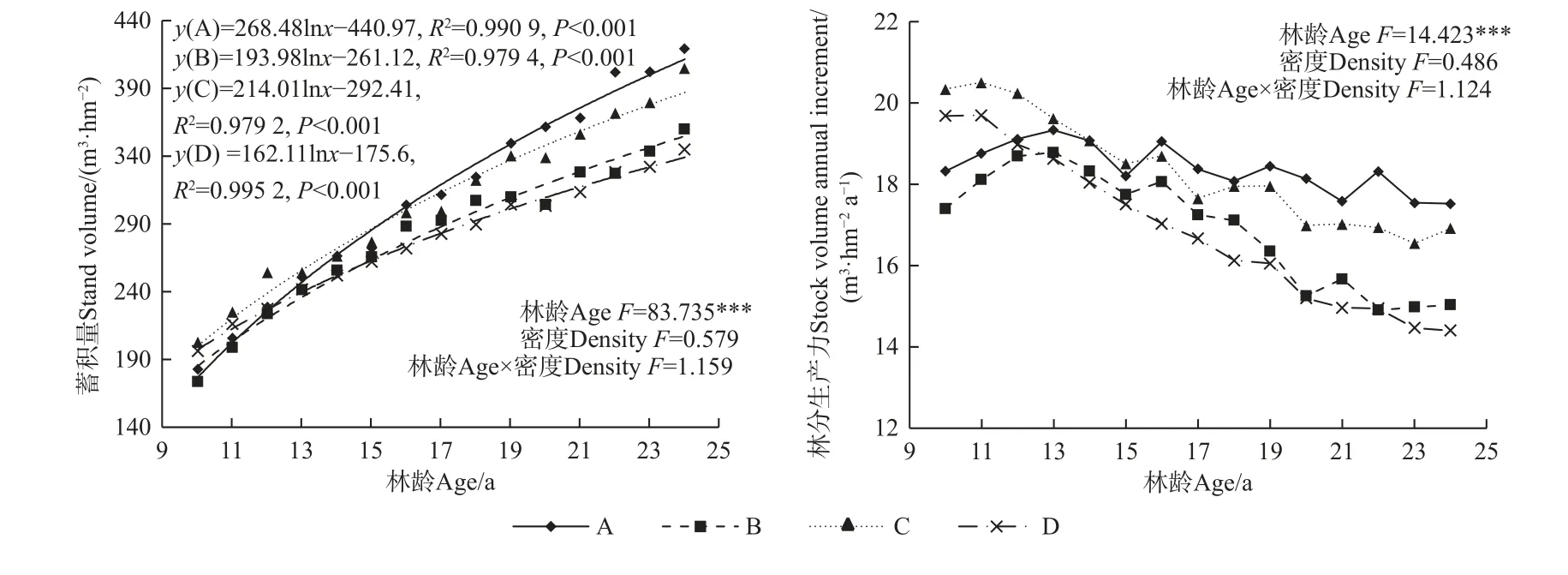
圖6 不同造林密度馬尾松人工林林分生產力變化Fig. 6 Changes in stock volume and stock volume annual increment with different densities of P. massoniana plantations
馬尾松人工林個體分化程度Gini 系數與林分蓄積量呈顯著負相關(P<0.001),與林分生產力呈顯著正相關(P<0.01),且Gini 系數越大林分蓄積量越小,林分生產力越大(圖7)。馬尾松人工林活立木株數與林分蓄積量呈顯著負相關(P<0.001),A 和B 密度林分生產力隨活立木株數增多呈先增加后減小的二次函數關系,均在2 000~2 500 株·hm-2時達到極大值,C 和D密度林分生產力隨活立木株數增多呈顯著正相關(P<0.001)。馬尾松人工林自然稀疏率與林分蓄積量和生產力僅在D 密度下呈顯著正和負相關(P<0.001),而A、B、C 密度無顯著相關。
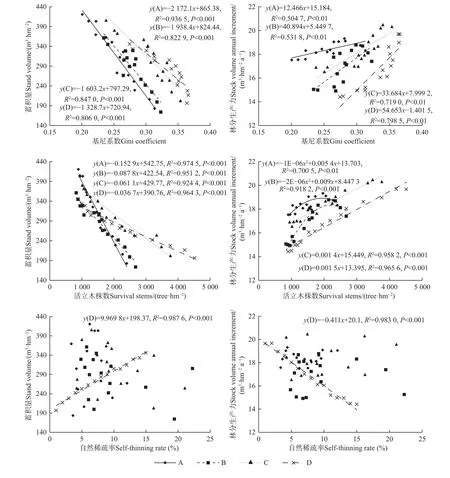
圖7 林分生產力與個體大小分化和自然稀疏效應的關系Fig. 7 The relationship between stock volume annual increment and individual size differentiation and self-thinning effect
2.4 林分生產力和Gini 系數的影響效應分析
隨機森林分析(圖8A)表明,林齡、造林密度和活立木株數對Gini 系數均具有顯著影響(P<0.01),自然稀疏率對Gini 系數無顯著影響,其中造林密度的影響效應最大(38.41%),其次是活立木株數(28.92%)和林齡(28.15%)。林齡、造林密度、活立木株數和Gini 系數對林分生產力均具有顯著影響(P<0.01)(圖8B),自然稀疏率對林分生產力無顯著影響,其中活立木株數的影響效應最大(32.03%),其次是造林密度(26.25%)、Gini 系數(24.64%)和林齡(23.23%)。
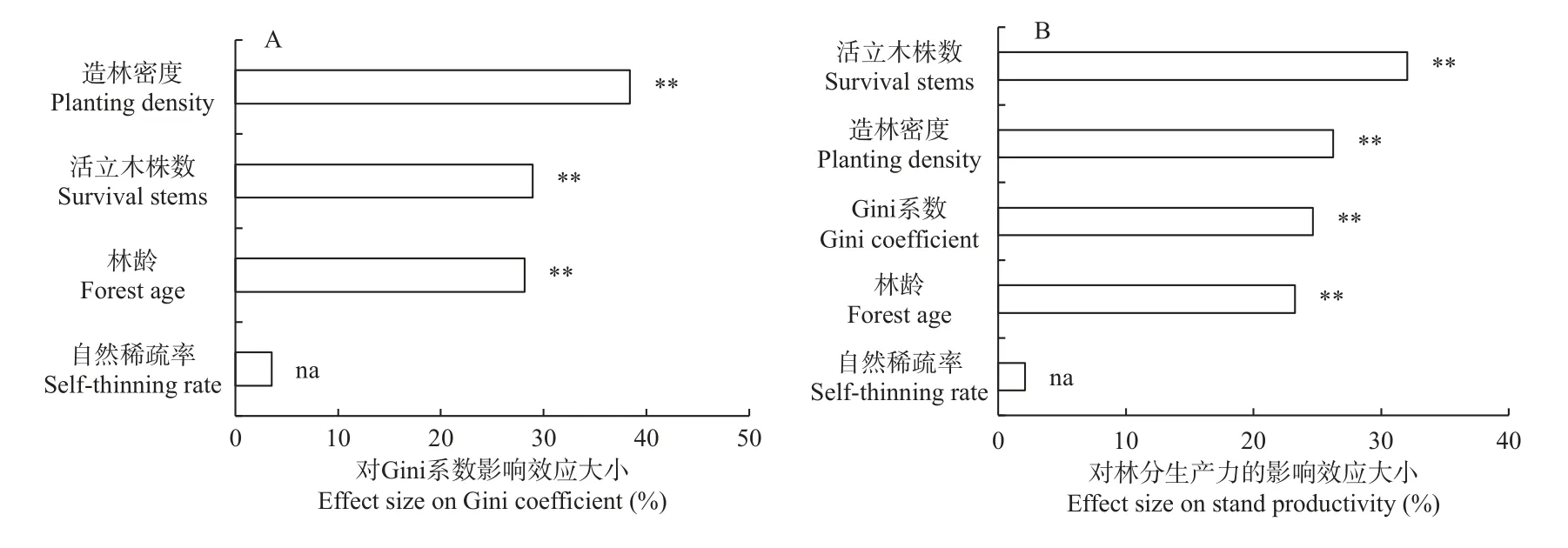
圖8 林齡、造林密度和自然稀疏對Gini 系數和林分生產力的影響效應分析Fig. 8 Effect analysis of forest age, planting density and self-thinning on Gini coefficient and stand productivity
結構方程顯示(圖9),林齡對馬尾松人工林活立木株數和Gini 系數均具有顯著負向影響(路徑系數分別為-0.798 和-0.464),對自然稀疏率和林分生產力具有負向影響(路徑系數分別為-0.163 和-0.072),活立木株數對林分生產力具有顯著正向影響(路徑系數為0.953),表明馬尾松人工林林分生產力隨林齡的變化主要是通過影響活立木株數變化的間接作用,即隨林齡增加經過自然稀疏過程,導致林分活立木株數改變而間接影響林分生產力。造林密度對Gini 系數具有顯著正向影響,對自然稀疏率、活立木株數均具有正向影響(路徑系數分別為0.252 和0.274),對林分生產力具有負向影響(路徑系數為-0.083),Gini 系數對林分生產力具有顯著負向影響(路徑系數為-0.63),表明造林密度對林分生產力產生的影響主要是通過不同造林密度條件對馬尾松人工林林分個體大小分化(Gini 系數)影響,從而間接影響林分生產力。
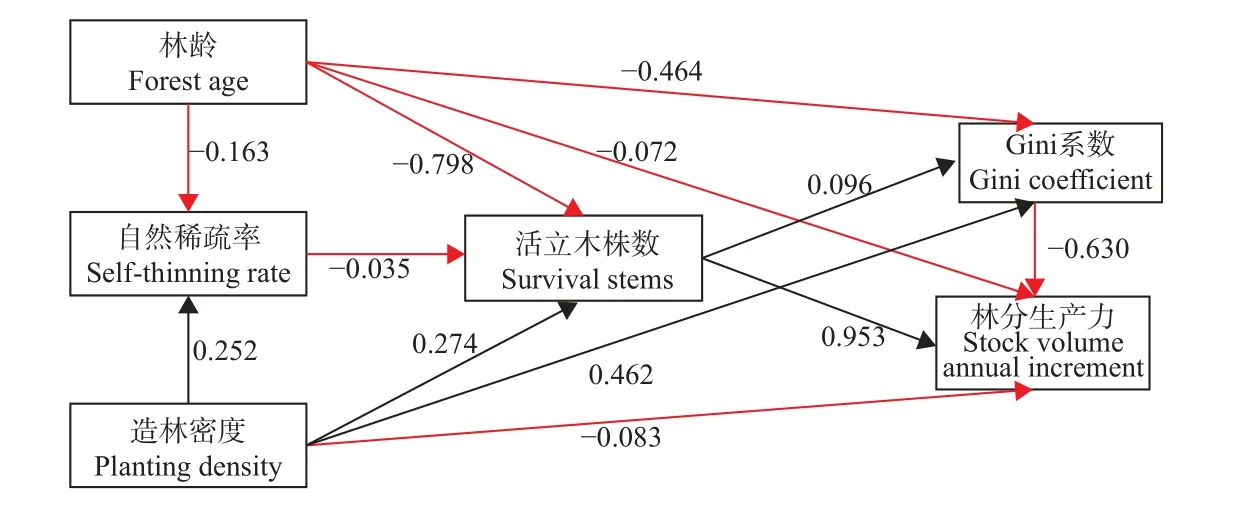
圖9 Gini 系數和林分生產力的影響路徑分析Fig. 9 Analysis of the impact path of Gini coefficient and stock volume annual increment
3 討論
林分內個體大小分化反映其對生長資源的獲取和利用差異,初始造林密度越大,林分進入競爭階段時間越早,進而導致林分內個體間分化程度越大(諶紅輝等,2011),與本研究馬尾松人工林個體間分化程度隨造林密度增加顯著增大的結果一致,均呈現D>C>B>A,表明A 密度條件具有保持較低林分分化水平。楊桂娟等(2019)對杉木(Cunninghamia lanceolata)人工林個體分化大小研究表明,個體間分化程度與林齡以及是否發生自然稀疏無關,與本研究結果相一致。而馬尾松人工林個體大小分化隨林齡增加顯著減小與杉木研究規律(楊桂娟等,2019)并不一致,其原因一方面是本研究采用的材料為同一家系,具有相對分化程度較小等優良特性,而杉木研究為種子園混系,個體差異較大;另一方面可能是因樹種不同,馬尾松具有向陽性、不耐陰,隨著林齡增加,林分內個體間競爭強度增大,林分優勢個體對相鄰劣勢個體產生冠層“欺壓”,根系伸展受限,被壓的個體很快發生自然稀疏而枯死(Sunet al., 2011),加之馬尾松活枝枝下高較高,林木存留可供光合作用針葉數量較少,林分內劣勢被壓木大量死亡(馬尾松自然稀疏率平均達10%),從而導致個體大小分化隨林齡增加而減小,這與本研究林分自然稀疏過程中活立木株數隨林齡增加顯著減小并逐漸趨于一致、個體大小分化隨活立木株數增加而增大的對數函數分布特征結果相一致。此外,本研究還發現林分自然稀疏過程中,林分死亡個體斷面積與林分平均斷面積比值(BAdeath: BAtotal)隨林齡增加顯著增大,且在19 年后均有出現>1 的情況,表明隨林齡增加林分自然稀疏過程中,死亡單株平均大小大于林分整體水平,死亡單株對林分整體影響隨林齡增加而加大。
林木大小徑階分布特征可用于評估林分結構,了解林分不同徑階水平的變化規律,可指導林分中大徑材的培育(Zasada,2013)。本研究表明,林齡對馬尾松人工林大、中和小徑材株數變化均具有顯著影響(P<0.01),徑階分布特征中14 年生馬尾松人工林開始出現大徑材,以A 密度(21 年生和17 年生)出現25%大徑材和50%中徑材最優,加之A 密度下林分蓄積量變化幅度最大,15 年間增加237.03 m3·hm-2,且16年后林分生產力均處于較大水平,綜合林分生長和中、大徑材演變的林齡效應來看,A 密度適用于馬尾松中大徑材人工林培育。此外,本研究發現C 密度是最晚出現小徑材低于50%的林分(20 年生),且15 年前均以C 密度下馬尾松人工林生產力水平最佳,表明C 密度可作為馬尾松短周期工業原料林培育模式,與陳旋等(2020)對馬尾松優良家系人工林生長及直徑分布的影響結果相一致。隨林齡增加林木進行光合作用,吸收利用養分累積生物量,導致林分蓄積量不斷增加(Jagodzińskiet al., 2018),與本研究馬尾松人工林林分蓄積量隨林齡呈對數函數顯著增加的結果相一致。不同林齡馬尾松人工林林分年均蓄積量(林分生產力)隨林齡增加呈現波動性變化,主要是由于林木生長受水、溫、干旱等自然環境長期作用的結果(馮源等,2019)以及林木生長大小年對生物量累積的效應,但整體呈先增加后減小并逐漸趨于穩定,主要是由于馬尾松人工林生長初期屬于快速生長階段,其快速累積生物量導致林分生產力水平快速增加,林木生長后期由于養分供應和種內競爭加劇,導致林分生產力有所下降并逐漸趨于穩定(吳鵬等,2011),而不同造林密度達到生產力峰值的時間并不一致,其中A 和B 密度在13 年生,C 和D 密度在11 年生,這主要是由于不同造林密度林分種內進入競爭的時間大小不一致,產生生態位林分化時間差異引起的(諶紅輝等,2011;丁貴杰,2003),林分密度越大,越早進入競爭階段,生產力達到峰值時間越短,與本研究結果相似。
隨機森林分析表明,林分生產力和Gini 系數均受林齡、造林密度、活立木株數的顯著影響,受自然稀疏率的影響并不顯著,其中Gini 系數受造林密度的影響效應最大,達38.41%,表明造林密度對林木個體大小分化影響效應較大,選擇適宜造林密度減小林木個體分化尤為重要。活立木株數對林分生產力的影響效應最大(32.03%),表明林分生產力主要由在競爭過程中具有優勢個體的生長狀況決定,發生自然稀疏被淘汰的劣勢個體對林分生產力和個體大小分化的影響較小。通過構建結構方程模型進一步明晰林齡、造林密度和自然稀疏過程對林分生產力和Gini 系數的直接或間接影響,可為明確馬尾松人工林受林齡、造林密度和自然稀疏的調控機制提供理論依據(Zhuet al., 2019)。結構方程模型顯示,林齡對馬尾松人工林活立木株數和Gini 系數均具有顯著負向影響,造林密度對Gini 系數具有顯著正向影響,這是因為隨著林齡增加,林分發生自然稀疏效應,活立木株數逐漸減小,優勢木所占有生態位加大,林分個體大小分化減小,而林分密度增大必然會引起林木生態位分化增大,個體大小分化增加。林齡對生產力的影響通過“林齡-活立木株數-林分生產力”的間接作用產生,其原因是隨林齡增加,林分內活立木株數改變,而林分生產力又由林分內優勢個體生長狀況決定(Fleminget al.,2014),從而導致林齡與生產力間的間接作用。造林密度對林分生產力的影響通過“造林密度-Gini 系數-林分生產力”的間接作用產生,表明不同造林密度通過調控林分內個體大小分化方式,進而調控林分生產力水平。因此,在生產實踐過程中,當林分生態位分化達到一定程度,通過撫育間伐方式,伐除被壓木既可以減小個體分化維持林分生產力不下降,又可以作為土壤養分的來源,成為優勢木生長的重要養分來源,維持土壤地力。
4 結論
馬尾松人工林林木個體分化隨林齡增加呈顯著線性負相關,隨造林密度增加顯著增大。林分自然稀疏過程中,活立木株數呈指數函數顯著減小,且造林密度越大林分內存活株數減小越快,減少數量越多;林分自然稀疏率較大的林齡為13~14 年和20 年生,表明在這2 個時間點林分內個體競爭程度較大,競爭劇烈;隨林齡增加死亡單株對林分整體影響逐漸加大,尤其是19 年生死亡單株平均大小大于林分整體水平。徑階分布特征中以A 密度最早出現25%大徑材和50%中徑材(21 年生和17 年生),C 密度最晚出現小徑材低于50%的林分(20 年生),且15 年間林分蓄積量增加順序為A(237.03 m3·hm-2)>C(202.51 m3·hm-2)>B(186.76 m3·hm-2)>D(148.95 m3·hm-2),綜合林齡效應和徑階分布來看A 密度適宜中大徑材馬尾松人工林培育。林齡、造林密度、活立木株數對林分個體分化和生產力均具有顯著影響,其中造林密度對林分個體的影響效應最大(38.41%),活立木株數對林分生產力的影響效應最大(32.03%),林齡和造林密度對林分生產力影響主要通過“林齡-活立木株數-林分生產力、造林密度-Gini 系數-林分生產力”的間接作用。本研究主要明晰林齡、造林密度和自然稀疏過程對馬尾松人工林個體分化特征的影響及其與林分生產力的關系,并通過構建各因素間結構通路為調控個體分化特征、提高林分生產力、確定適宜南亞熱帶地區馬尾松人工林經營密度提供依據。
