羅非魚無乳鏈球菌gap 基因的原核表達及免疫原性研究
2023-11-06 08:08:56孫彤彤黃遠城李恭賀鄭喜邦吳文德
現代畜牧獸醫 2023年10期
關鍵詞:血清
楊 洋,農 源,陳 林,孫彤彤,黃遠城,李恭賀,2,3,4,鄭喜邦,2,3,4,吳文德,2,3,4*
( 1. 廣西大學動物科學技術學院,廣西 南寧 530004 ; 2. 廣西壯族自治區獸用生物制品工程研究中心,廣西 南寧 530004 ; 3. 廣西畜禽繁育與疾病防控重點實驗室,廣西 南寧 530004 ; 4. 廣西高校動物疫病預防與控制重點實驗室,廣西 南寧 530004 )
羅非魚(Oreochromis niloticus)具有耐受性強、飼料轉化率高等優點,具有極高的市場需求[1]。無乳鏈球菌可引起羅非魚大量死亡,造成巨大的經濟損失[2-3]。因此,有必要制定適用于羅非魚養殖的預防或疾病治療措施。目前控制羅非魚鏈球菌病的主要方法是使用抗生素,但易產生耐藥菌株及藥物殘留,不符合綠色防控的原則[4-7]。免疫接種從提高魚體免疫力出發,因其高效特異、環境友好被認為是水產病害防控最具有潛力的手段之一。細菌表面蛋白完全暴露于宿主免疫系統,并能夠誘導高水平的免疫反應。gap基因的產物三磷酸甘油醛脫氫酶(GAPDH)廣泛存在于生物界,通常位于細菌細胞表面,可在感染過程中與宿主基質蛋白結合。有報道在感染小鼠的無乳鏈球菌中檢出GAPDH,并認為其是一種毒力相關的免疫調節蛋白[8]。GAPDH已被確定為許多病原體的候選疫苗,有研究發現[9],母體免疫rGAPDH 可保護新生小鼠免于無乳鏈球菌感染引起的死亡,表明無乳鏈球菌GAPDH 可作為一種合適的靶抗原制備抗無乳鏈球菌感染的有效疫苗。也有研究表明,來自格氏乳球菌的GAPDH 蛋白對羅非魚乳球菌病具有有效的保護作用[10]。本研究對羅非魚無乳鏈球菌gap基因進行擴增,通過原核表達系統表達并純化重組蛋白,動物試驗分析其免疫原性和保護效果,為進一步利用該蛋白防控羅非魚無乳鏈球菌病奠定基礎。
1 材料與方法
1.1 試驗材料
1.1.1 試驗動物
試驗羅非魚200 尾購自廣西某水產養殖有限公司,體重為(50±2) g,體長(13.0±0.5) cm。試驗前暫養10 d,水溫(30.0±0.5) ℃,氣泵24 h 供氧,飼料每日投喂體質量的2%。本試驗于廣西大學水產基地進行。
1.1.2 試劑與儀器
Premix Taq?、pMD?18-T 載體克隆試劑盒、限制性內切酶BamH Ⅰ/Sal Ⅰ、T4 DNA Ligase 購自Takara 公司;DH5α和BL21感受態細胞、抗His標簽鼠單克隆抗體、HRP標記羊抗鼠IgG(H+L)抗體購自北京全式金生物技術有限公司;IPTG 粉末、透析袋、弗氏完全佐劑和不完全佐劑購自Sigma 公司;pET-28a-SUMO 表達載體、HRP 標記的抗羅非魚IgM單克隆抗體、無乳鏈球菌HN016株由廣西大學動物科學技術學院獸醫臨床實驗室保存。
PCR 儀、水平電泳儀(賽默飛公司);細胞超聲波破碎儀(南京先歐儀器制造有限公司);垂直電泳儀(北京六一生物科技有限公司);凝膠分析系統(上海天能生命科學有限公司);低溫高速離心機(Sigma 公司);化學發光成像儀(Cytiva公司);酶標儀(Bio-Rad公司)。
1.2 試驗方法
1.2.1 重組載體pET-28a-gap的構建
無乳鏈球菌提取全基因組DNA,根據GenBank登錄號WP_006738473.1 設計引物:gap_F:5'-GGATCCatggtagttaaagttggt-3',gap_R:5'-cGTCGACttattttgcaatttttgc-3'。
PCR擴增反應體系:上下游引物各10 nmol/L、DNA模板50 ng、Premix Taq 25 μL,加ddH2O 至50 μL。反應程序:95 ℃變性30 s,48 ℃退火50 s,72 ℃延伸2 min,共30 個循環。對PCR 產物進行電泳并回收。gap 片段與表達載體pET-28a-Sumo 進行BamH Ⅰ、Sal Ⅰ分別雙酶切并用T4 連接酶連接。連接產物轉化E.coliDH5α 克隆菌株,提取質粒,進行雙酶切和測序鑒定,測序正確的質粒命名pET-28a-gap。
1.2.2 重組載體誘導表達條件的優化
構建成功的質粒pET-28a-gap 轉化E.coliBL21(DE3)表達菌株,挑取單克隆到含Kan 抗性的TB 培養基中,培養至OD600nm值為0.6~0.8時,加入終濃度分別為0.1、0.2、0.4、0.8、1.2、2.0、3.0 mmol/L 的IPTG,在16、23、30、37 ℃的不同溫度下,220 r/min誘導4、8、12、16 h后,同時設立轉化pET-28a-SUMO空質粒的BL21(DE3)菌株和未加誘導劑的pET-28a-gap 重組質粒的BL21(DE3)菌株分別作為空白和陰性對照。離心收集菌體并通過聚丙烯酰胺凝膠電泳(SDS-PAGE)檢測目的蛋白的表達情況,確立最優表達條件。取適量最優表達條件下的菌體,超聲破碎后分離上清與沉淀,SDS-PAGE分析重組蛋白表達形式。
1.2.3 重組蛋白的Western blot鑒定
設立空質粒pET-28a-SUMO 作為空白對照,與誘導表達的菌液變性并經SDS-PAGE電泳后,通過濕轉法轉至PVDF膜后用5%脫脂奶粉封閉,以抗His標簽鼠單克隆抗體為一抗,HRP標記山羊抗小鼠IgG(H+L)抗體為二抗,按照抗體說明書稀釋后進行孵育,孵育結束后清洗PVDF膜,采用ECL化學發光法顯影并采集圖像。
1.2.4 重組蛋白的純化
按照1.2.2構建的條件大量培養重組載體轉化菌,收集菌體沉淀并清洗后,冰上超聲波破碎菌液,低溫離心收集上清,0.45 μm濾器過濾,Ni瓊脂糖凝膠柱進行親和層析純化。對收集的洗脫液進行SDS-PAGE分析,純化后的蛋白用7000D透析袋除去雜鹽和濃縮,并檢測其蛋白濃度。
1.2.5 重組蛋白疫苗的制備及免疫原性檢測
健康羅非魚隨機分為攻毒組、免疫組、空白組,每組50 尾。用無菌PBS 對純化的gap 蛋白進行稀釋并與弗氏佐劑進行1∶1 混合乳化,按照2 μg 蛋白/1 g 魚體重的劑量對免疫組進行腹腔注射,攻毒組和空白組注射同體積的PBS 與佐劑混合物。試驗魚共接受3 次免疫,兩次注射間隔8 d,首免使用弗氏完全佐劑,后續兩次加強免使用弗氏不完全佐劑。3 次免疫前和攻毒前后每組隨機抽取10 尾魚尾靜脈采血0.5 mL,血樣4 ℃過夜后分離血清,采用間接ELISA方法測定血清免疫球蛋白M(IgM)抗體水平。
1.2.6 重組蛋白疫苗的免疫保護作用
免疫結束后用無乳鏈球菌對羅非魚進行攻毒試驗。菌體濃度1×108CFU/mL。攻毒組和免疫組每尾魚腹腔注射0.1 mL 無乳鏈球菌,空白組注射同體積的生理鹽水,按照公式計算各組相對免疫保護率(RPS)。
1.3 數據統計與分析
試驗數據采用IBM SPSS Statistics 25.0軟件進行單因素方差分析,GraphPad Prism 9.0軟件制圖。P<0.05表示差異顯著,P>0.05表示差異不顯著。
2 結果與分析
2.1 載體pET-28a-gap的構建及鑒定(見圖1)
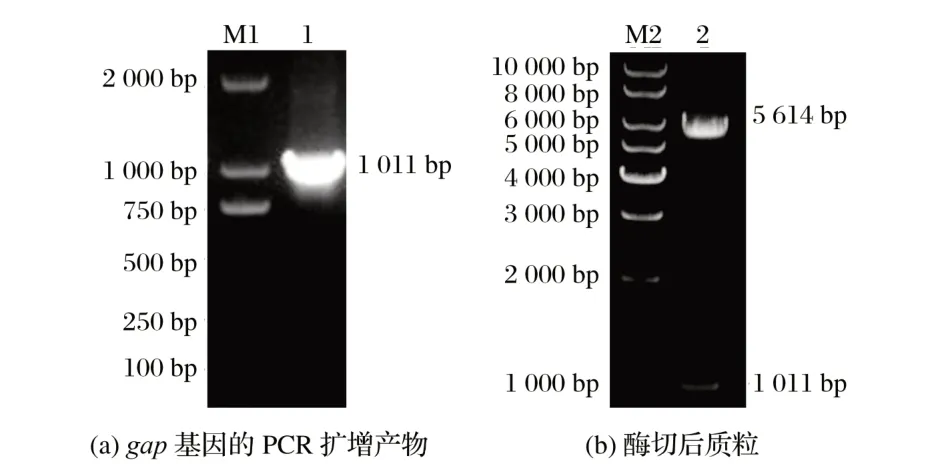
圖1 載體pET-28a-gap的構建及鑒定Fig.1 Construction and identification of carrier pET-28a-gap
由圖1(a)可知,PCR結果為單一且清晰的1 011 bp的片段,與預期結果一致,表明成功擴增gap基因。
由圖1(b)可知,構建的載體經雙酶切鑒定,獲得5 614和1 011 bp 的兩個片段,質粒DNA 測序與預期序列相同,表明重組載體pET-28a-gap成功構建。
2.2 重組蛋白gap高效表達體系的建立(見圖2~圖4)
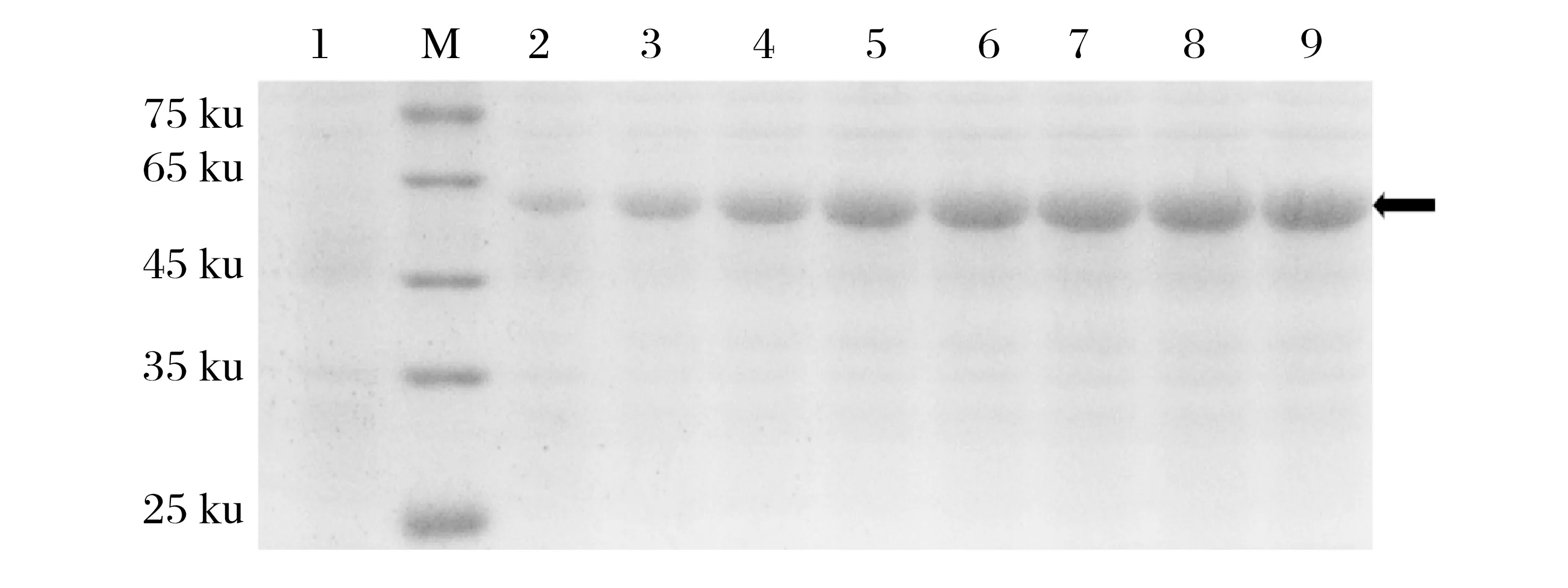
圖2 誘導劑濃度的表達條件優化Fig.2 Optimization of expression conditions of inducer concentration
重組表達載體目的蛋白的分子量理論值為49.8 ku,轉化重組載體pET-28a-gap 的E.coliBL21(DE3)菌株在各條件下均能夠在預期位置成功表達重組蛋白。
由圖2 可知,低濃度下蛋白表達量與誘導劑濃度呈正相關,但0.4 mmol/L以上時濃度蛋白含量不再隨著誘導劑濃度的增大而呈明顯變化,選擇IPTG 0.4 mmol/L為gap蛋白的最優誘導劑濃度。
由圖3 可知,gap 蛋白的表達量隨著誘導溫度增高而增加,但誘導溫度超過23 ℃后表達量不再出現明顯增加,低溫能夠保持蛋白質的活性,選擇23 ℃為gap蛋白的最優誘導溫度。
由圖4 可知,gap 蛋白的表達量隨著誘導時長增加而增加,誘導時間超過12 h后表達量下降,選擇12 h為gap蛋白的最優誘導時長。
2.3 重組gap 蛋白的表達形式分析與Western blot 檢測(見圖5)

圖5 重組gap蛋白的表達形式分析與Western blot檢測Fig.5 Analysis of expression forms and western blot analysis of recombinant protein gap
由圖5(a)可知,在0.4 mmol/L IPTG、23 ℃條件下誘導12 h,gap蛋白主要以可溶性蛋白的形式表達在上清中,沉淀中幾乎未見表達的gap蛋白。由圖5(b)可知,在預期位置49.8 ku處出現清晰的反應條帶,空質粒未出現明顯的免疫印跡,表明重組質粒pET-28a-gap成功表達了gap蛋白。
2.4 重組gap蛋白的SDS-PAGE分析(見圖6)
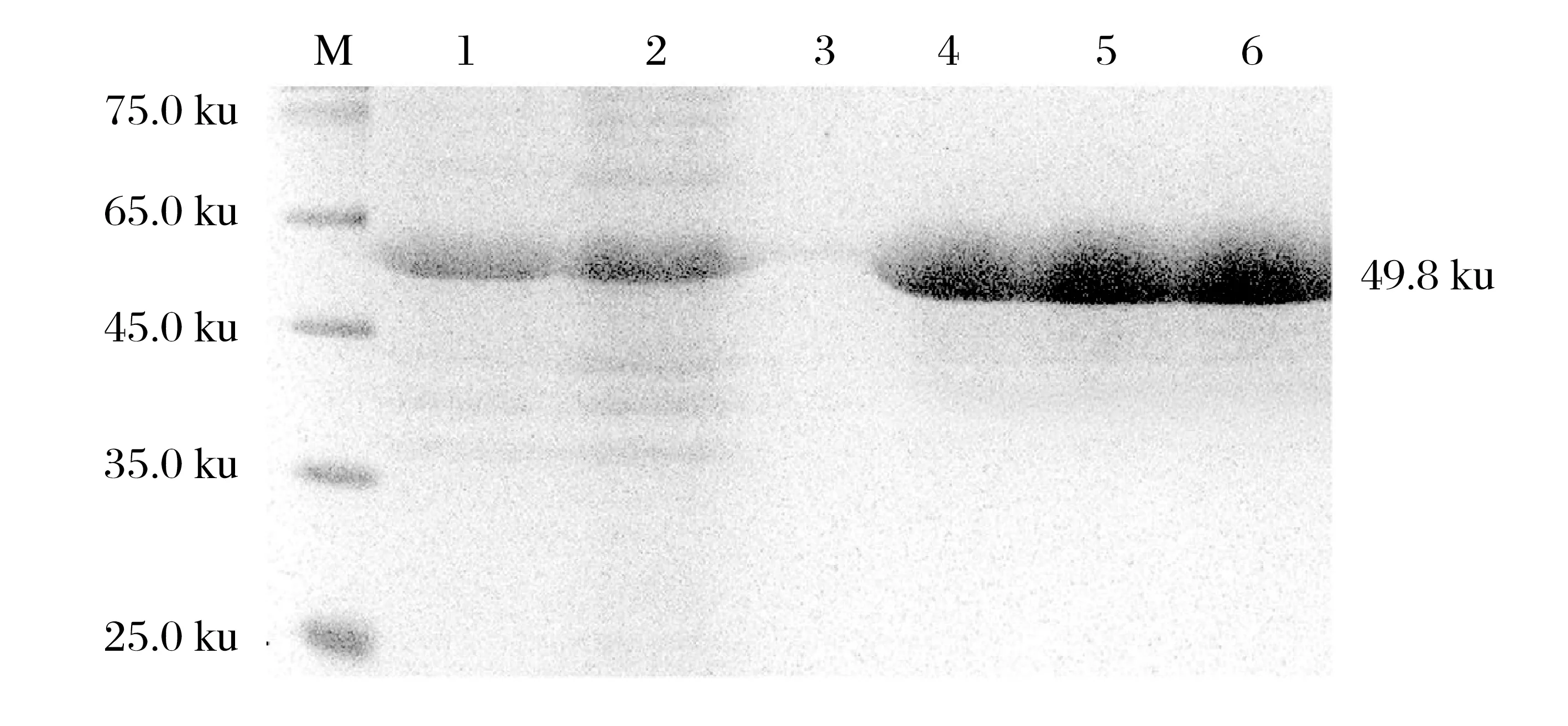
圖6 重組gap蛋白的SDS-PAGE分析Fig.6 SDS-PAGE analysis of purified protein gap
由圖6可知,不同階段的洗脫液通過SDS-PAGE分析進行比較,純化后各管洗脫液均出現大小為49.8 ku的高濃度單一條帶,且與純化前的目的條帶位置相同;流穿液的初段含大量雜蛋白條帶與目的條帶,末段不再含有條帶,表明已洗脫充分。經檢測蛋白濃度約為2.62 g/L。
2.5 重組蛋白疫苗免疫對羅非魚血清抗體效價的影響(見圖7)
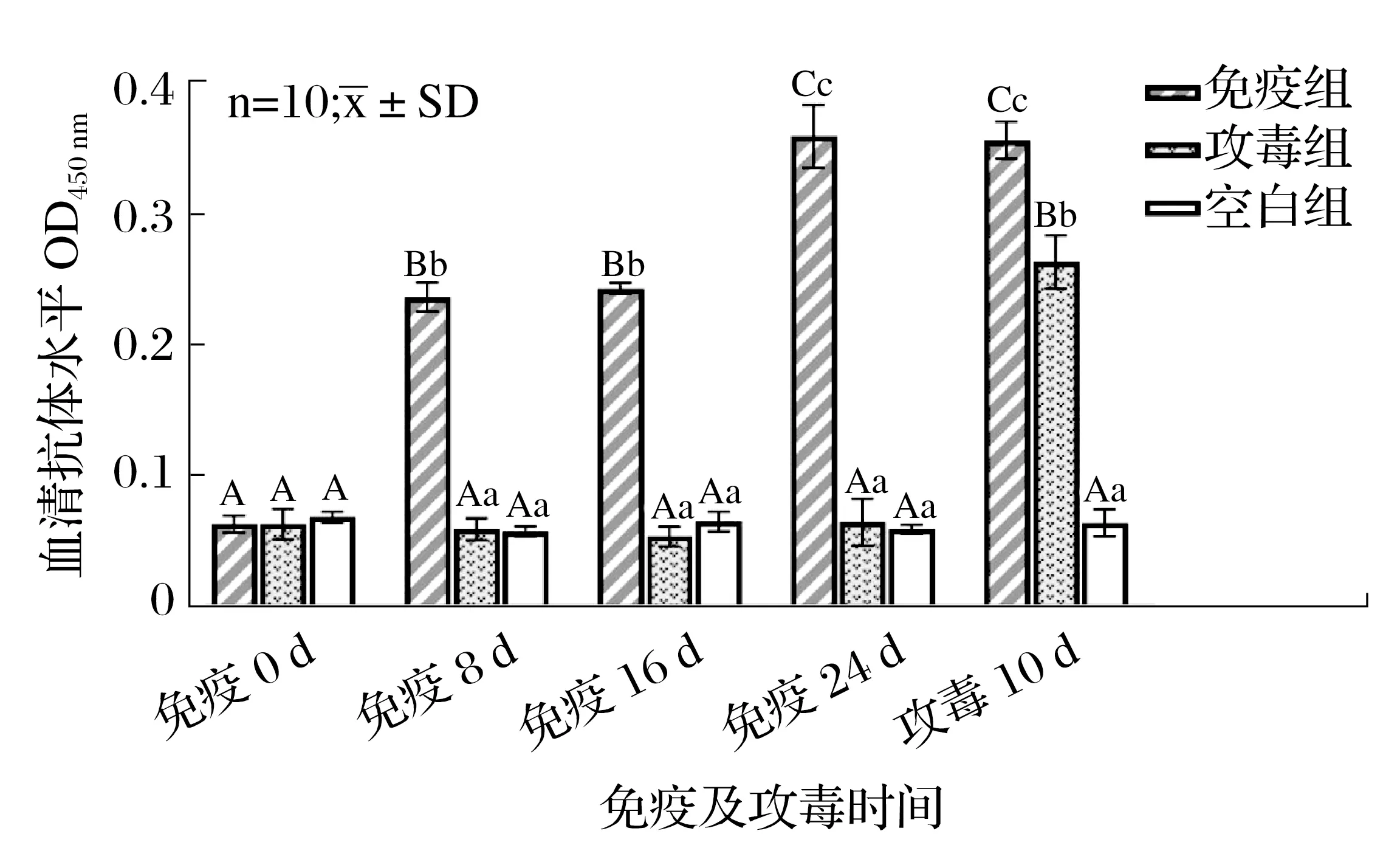
圖7 重組蛋白疫苗免疫對羅非魚血清抗體效價的影響Fig.7 Detection of ELISA antibody level of tilapia at different time periods
由圖7 可知,免疫前各組羅非魚血清抗體效價差異不顯著(P>0.05);免疫8 d后,免疫組羅非魚血清抗體效價顯著高于未免疫組(P<0.05);免疫組羅非魚接受一次免疫注射后血清抗體效價與接受兩次注射差異不顯著(P>0.05),接受3 次免疫注射后的血清抗體效價顯著高于前兩次(P<0.05);攻毒結束后,免疫組羅非魚血清抗體效價顯著高于攻毒組和空白組(P<0.05),免疫組和攻毒組羅非魚血清抗體效價均顯著高于空白組(P<0.05)。
2.6 重組蛋白疫苗免疫對羅非魚的保護效果(見圖8)
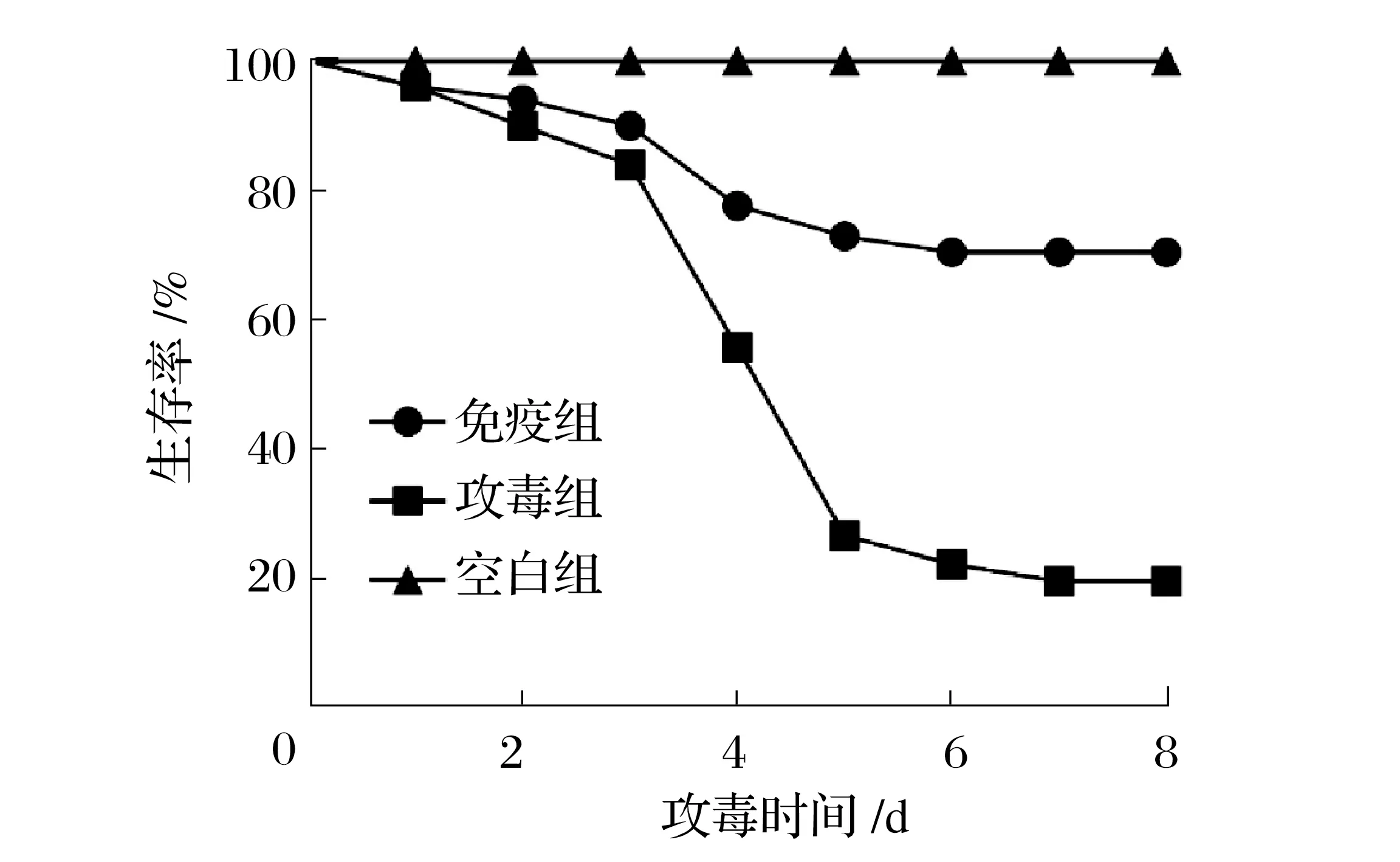
圖8 重組蛋白疫苗免疫對羅非魚的保護效果Fig.8 Protective effect of recombinant protein vaccine on tilapia
羅非魚攻毒后1 d 免疫組和攻毒組均開始出現死亡,攻毒后4 d 達到死亡高峰,7 d 后不再發生死亡。攻毒后,病魚體色發黑,停止進食,魚體失衡,在水面轉圈,最后沉底死亡,為典型無乳鏈球菌感染癥狀;剖檢腹腔有腹水,臟器組織分離并劃線血平板均可見無乳鏈球菌生長。由圖8可知,無乳鏈球菌腹腔注射羅非魚后,免疫組、攻毒組、空白組的死亡率分別為28%、78%、0,相對保護率為64.1%。
3 討論
無乳鏈球菌是引起羅非魚鏈球菌病的主要病原體[11],能否有效控制該病已成為水產養殖研究的重點。為符合食品安全和糧食安全政策,抗生素等逐漸被禁用,因此通過疫苗接種激活魚類的自我免疫力是控制鏈球菌病最實用、最可靠的方法。目前,魚用無乳鏈球菌疫苗趨于多元化發展,包括滅活疫苗、減毒疫苗、DNA疫苗、亞單位疫苗等。然而,滅活疫苗的功效性和減毒活苗的安全性有待考證[12]。近年來,重組蛋白疫苗研究領域受到了越來越多的關注,有以下3方面原因:首先,滅活疫苗等可能出現“毒力返祖”現象,造成抗原損害,重組蛋白疫苗很少出現安全性問題;其次,重組蛋白疫苗選擇高度保守區域進行表達,可用作對抗多種血清型病原體的通用疫苗;最后,該疫苗可以區分動物是否被接種,可作為標記疫苗。膜蛋白最先被宿主的免疫系統識別,激發宿主機體產生相應的免疫機制。近年來,相關免疫蛋白的報道屢見不鮮,大多均可列為亞單位疫苗的候選蛋白[13-14]。gap基因的產物蛋白是糖酵解途徑中的一種關鍵代謝酶,具有多種生物學活性,同樣能夠幫助病原菌對宿主的黏附和侵染[15],其生物學功能在魚類病原體如氣單胞菌、弧菌、愛德華氏菌、海豚鏈球菌以及副乳鏈球菌和停乳鏈球菌中均有報道,但關于對魚源無乳鏈球菌gap蛋白的分析尚未見報道。
本研究擴增出gap基因的序列經過Blast 比對與野毒株重合度100%,高度保守,表明選擇此膜基因作為蛋白疫苗,預期免疫效果相對穩定。包涵體形式表達的蛋白,需要專用溶解液將蛋白先變性再復性[16],過程中存在大量損耗;該蛋白主要以可溶性形式表達,純化簡便,產量高。蛋白足量表達純化后制成重組蛋白疫苗免疫羅非魚,并對其免疫保護作用進行評估。古麗米熱·對山巴依等[17]用馬腺疫鏈球菌GAPDH 蛋白免疫小鼠,結果也在小鼠血清中檢測到較高水平的特異性抗體IgG。本研究數據分析顯示,間接ELISA可檢測到相應的抗體,且隨著免疫接種次數和劑量的增加,血清抗體水平呈現上升趨勢,與曾祖聰等[18]得出的結論一致。對于無乳鏈球菌感染后存活的羅非魚,攻毒組血清中抗體水平顯著升高,免疫組前后無顯著變化,猜測血清抗體水平不能代表羅非魚對無乳鏈球菌抗感染的能力。攻毒羅非魚觀察試驗結果發現,發病初期,免疫組和攻毒組死亡情況相當,4 d后攻毒組持續死亡,免疫組死亡情況趨于平緩,直至不再死亡,表明對于外來致病菌的攻擊需要一定的時間啟動體液免疫[19]。
4 結論
本研究利用PCR 對羅非魚無乳鏈球菌gap基因進行擴增,構建原核表達載體pET-28a-gap,優化表達條件,獲得高純度重組gap 蛋白,通過腹腔注射羅非魚驗證其免疫原性。結果顯示,gap基因表達的重組蛋白能夠顯著誘導羅非魚體內的免疫應答,可作為防控羅非魚無乳鏈球菌感染的候選亞單位疫苗。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
