我國玉米南方銹病的抗病育種進展和防治策略
2023-11-18 06:59:32任雅琨朱勝男計懷春王啟磊
中國種業 2023年11期
任雅琨 韓 托 朱勝男 計懷春 王啟磊
(1 農業農村部玉米小麥等作物優質抗逆精準育種重點實驗室,鄭州 450001;2 河南金苑種業股份有限公司,鄭州 450001)
玉米南方銹病是由多堆柄銹菌引起的一種氣流性傳播病害,主要危害玉米葉片、葉鞘。最初廣泛發生在非洲、東南亞、美洲南部等熱帶和亞熱帶地區,隨著全球熱帶氣旋的活動,該病害在全球范圍內不斷擴大,已在我國22 個省份發生,對黃淮海夏玉米區和南方玉米區的危害尤為嚴重,嚴重時可導致減產50%以上。2015 年南方銹病在全國致病區域達到一個高峰,發病面積達523.9 萬hm2,實際損失約75.6 萬t[1]。2021 年玉米南方銹病在黃淮海區域再次大爆發,山東省發生面積達歷史之最,發病面積占山東省玉米總種植面積的43.2%,平均病株率46%[2]。近年來受西太平洋熱帶氣旋頻繁發生的影響,南方銹病在我國的發病面積和嚴重程度不斷擴大,對我國玉米主產區的安全生產構成嚴重威脅。抗病育種是防治南方銹病最經濟有效的方法。了解我國玉米品種的抗性水平、鑒定抗病種質、挖掘和利用抗病基因仍是抗病育種的重要工作,并在此基礎上探討了南方銹病的防治策略。
1 我國玉米品種對南方銹病的抗性評價
了解我國當前玉米品種對南方銹病的抗性水平,是品種改良和抗病育種的基礎。近年來多位學者對我國參試、已審和主推玉米品種進行南方銹病抗病性鑒定。劉駿等[3]從黃淮海區域47 個主推品種中鑒定出2 份高抗南方銹病品種,14 份抗性品種,先玉335、鄭單958、浚單20 均為高感品種。王群[4]在2017-2018 年從山東省400 份玉米品種中鑒定出63 份南方銹病抗性品種,其中高抗和抗性品種分別有3 份和24 份。劉炎琨[5]在2019-2020 年對358份河南省引種和區域試驗品種進行抗性鑒定,高抗、抗和中抗南方銹病的品種分別有66 份、80 份和111 份。黃莉群等[6]在2020 年對國內85 個主栽玉米品種進行田間抗性評價,高抗品種僅有9 份,占比10.59%,中抗品種19 份,占比22.35%。郭寧等[7]在2018-2020 年黃淮海夏玉米區國審的437 個品種中未鑒定出高抗品種,抗和中抗品種占比4.45%和4.92%。在47 個主栽品種中,抗性品種和中抗品種占比12.22%和27.66%。總體而言,我國玉米品種對玉米南方銹病的抗性較差,高抗品種少,可能是由于我國玉米育種使用的抗性種質資源狹窄,過度使用78599 系列,造成本土種質和品種趨同化嚴重且抗性較差[8]。然而,近年來抗病品種比例呈逐年上升趨勢,說明科研院所和育種單位對抗銹病品種的選育工作的重視程度增加。因此,拓寬種質資源、創新育種模式以及選用不同的抗病種質仍是未來抗病育種的重要方向。
2 我國玉米抗南方銹病種質的鑒定和利用
2.1我國南方銹病抗性種質的鑒定培育抗病品種的關鍵是選育抗病自交系,鑒定和利用多種抗病種質是選育抗病自交系的基礎。較多學者不斷地搜集和鑒定南方銹病抗性種質。黃飛燕[9]對1136 份玉米種質資源進行南方銹病抗性鑒定,高抗、抗、中抗種質所占比例分別為2.46%、7.13%和29.93%,國外地方品種的抗性比例高于國內地方品種的抗性比例。江凱等[10]從1589 份玉米種質資源中鑒定出137 份抗南方銹病材料;高抗材料僅26 份,其中16份來自于國外材料(美國材料選系5 份、CIMMYT材料7 份、非洲津巴布韋材料4 份),國內材料只有10 份。姚國旗等[8]鑒定了34 份熱帶亞熱帶自交系,發現僅自交系CML144、CML247、CML451 和CML470 高抗南方銹病。陳文娟等[11]在廣西和北京對903 份種質資源做了南方銹病抗性鑒定,高抗材料占比0.9%,抗性材料占比2.3%,中抗材料占比11.1%,高抗和抗性材料主要源自內蒙古和山西。此外作者用56 份抗性材料進行了聚類分析,可將抗性材料分為兩大類群,第一大群又被劃分成5 個亞群,包含PB 亞群、唐四平頭亞群、旅大紅骨亞群、BSSS亞群和Lan 亞群;第二大群為PA 群,與其他亞群材料的遺傳相似度較低,表明抗病材料有較高的遺傳多樣性。以上研究結果顯示我國國內高抗南方銹病的種質資源較少,且多數高抗南方銹病的資源來自國外熱帶種質資源。
2.2南方銹病抗性種質資源的利用熱帶種質資源在我國玉米育種中多數無法直接利用,需要與本土優異種質多次回交,將熱帶抗病種質中的抗銹病基因導入到本土優異材料中,才能實現本土自交系南方銹病抗性基因的多元化。任轉灘[12]將熱帶種質CML311 與掖478 雜交,經多代混粉后自交選擇,選出了既具有掖478 的優異性狀又兼備南方銹病抗性的新材料;用同樣的方法將CML246 與CA971雜交后再與Mo17 雜交,經多代自交選擇,最終篩選出了高抗南方銹病的適應性種質;除此之外,B73、K12、黃早四、黃金96B 等常用自交系也通過此方法提高了抗銹病能力。張志方等[13]以高抗南方銹病的M119 為基礎材料,經多代自交和姊妹交,經河南、貴州、海南三生態區的混合抗性選擇和配合力測定,選出了塘四平頭類型的抗病優異自交系浚M9;之后用浚M9 對本地種質進行改良,形成高抗莖腐病和南方銹病的優異種質浚96、浚M97、浚M98。同時用浚M9 與國外優異種質和國內骨干自交系進行選系,構建了1 個廣適多抗父本群。新的抗病自交系的育成也證明了利用熱帶抗病資源培育本土化抗病自交系的可行性。進一步利用CIMMYT、Suwan 和非洲等熱帶地區的抗南方銹病種質資源,仍是抗病自交系選育的關鍵環節。這些資源能夠為我國抗病育種提供非常優秀的抗性基礎材料。
3 玉米南方銹病抗病基因的挖掘及應用
3.1玉米南方銹病抗病基因的定位和克隆基于南方銹病的發病范圍不斷擴大,對玉米產量造成的損失程度不斷增加,國內外科研人員對玉米南方銹病基因的挖掘開展了較多研究。Storey 等[14-15]從抗南方銹病材料AFRO.29 和AFRO.24 中分別發現了完全顯性抗病基因Rpp1和不完全顯性抗病基因Rpp2,Rpp1專抗小種EA1,Rpp2對小種EA1和EA2 具有部分抗性;Rpp1與Rpp2連鎖,遺傳距 離 約 有12.23cM。Ullstrup[16]從PI 186208 中 定位到一個專抗小種PP.9 的單顯性抗病基因Rpp9;Storey 等[17]又發現了對EA1 和EA3 有抗性的顯性基因Rpp10和對EA1 和EA3 有不完全抗性的抗病 基 因Rpp11,且Rpp1、Rpp10和Rpp11均 不 連鎖。我國研究顯示多數南方銹病抗病位點定位在10 號染色體短臂上,且為單顯性主效基因。不同抗性材料中定位的抗性基因及連鎖的分子標記見表1[18-28]。Zhou 等[18]將齊319 的抗病基因RppQ定位于分子標記MA7、M-CCG/EAGA157 之間。Liu等[19]將抗性基因RppP25定位在10 號染色體上,與phi059 的遺傳距離為5.8cM;Zhao 等[20]進一步將RppP25定位在分子標記P091 和M271 之間,GRMZM2G060884可能為候選基因。Zhang 等[21]從W2D 和W222 構建的分離群體中定位主效單顯性抗病基因RppD,與分子標記umc1291 和CAPS858的遺傳距離分別為2.9cM 和0.8cM。RppD與RppQ和RppP25不等位。Wu 等[22]用熱帶材料SCML205與京24 構建F2分離群體,將抗病基因RppS定位在10 號染色體短臂遠端,與IDP4283 和G047 的遺傳距離為8.4cM。RppS與RppQ、RppD和RppP25均不等位。姚國旗等[23]發現CML470 的抗性主要由顯性單基因RppC控制,該基因定位在分子標記umc1380 與umc1291 之間,與2 個分子標記的遺傳距離分別為3.5cM 和8.8cM。王兵偉等[24]用S313×PHW52 的F2分離群體將抗病基因RppS313定位在SNP 分子標記A005915 與 A009920 之間,區間內有3 個候選基因。艾堂順等[25]從P178 和G41 構建的BC2F5群體中發現主效抗南方銹病位點qSCR10.01,可解釋45.31%的表型變異,該QTL 被定位在分子標記umc1380 和C(10)3595071 之間,物理距離1.34Mb。王文潔等[29]將高抗南方銹病材料K381 的抗病位點定位在分子標記umc2018 與phi059 之間,與phi059 緊密連鎖。
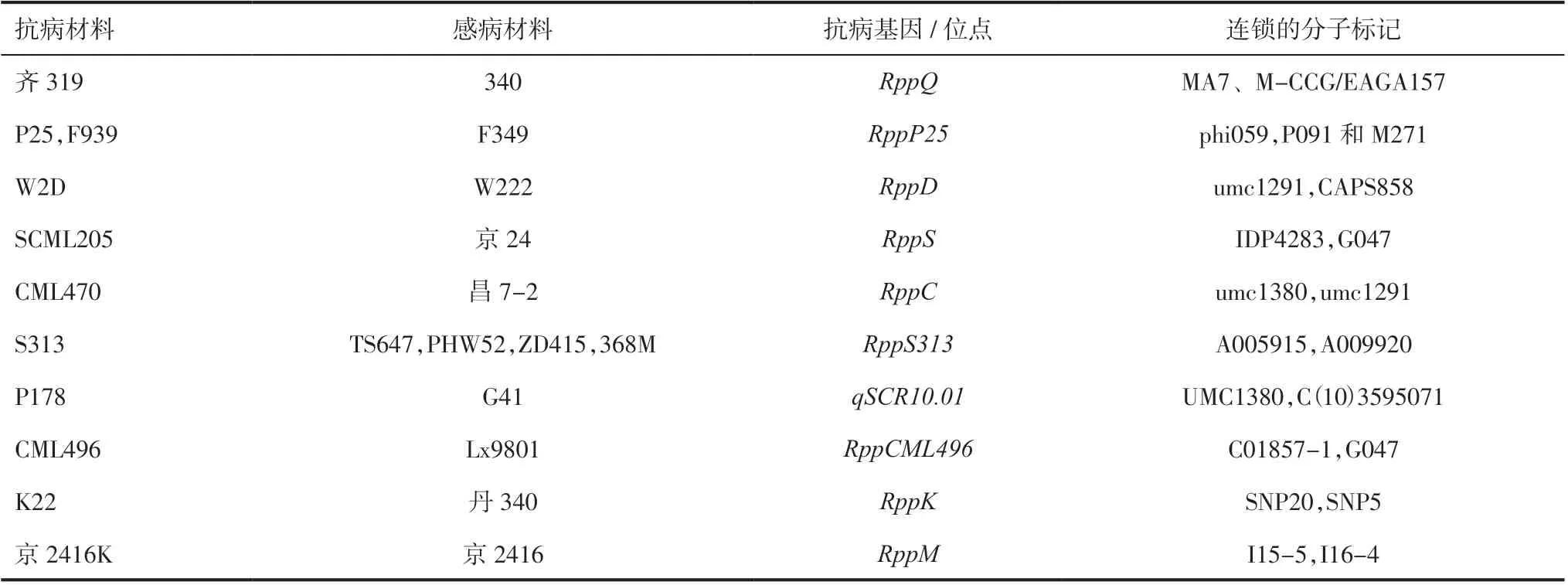
表1 不同抗性材料中抗性基因及連鎖的分子標記
除主效基因的定位,科研人員在玉米的1、2、3、4、5、6、8、9 號染色體上也發現了南方銹病的抗病位點[25,30-32]。其中Lu 等[30]用齊319 和掖478 構建重組自交系,在第6 號染色體上發現的qSCR6.01位點抗病效應最大,可解釋表型總變異的24.15%。
南方銹病抗性位點報道較多,但目前只有3個 抗 病 基 因 被 克 隆。Lv 等[26]和Deng 等[27]以CML496 和Lx9801 構建的重組自交系為材料,采用圖位克隆和轉基因驗證的方法,從CML496 的10 號染色體上克隆到一個NLR 類的抗玉米南方銹病的新基因RppC。利用轉錄組分析和玉米原生質體超敏反應篩選體系等多種研究方法,在南方銹病病原菌中鑒定到被RppC識別的效應蛋白AvrRppC,并發現新的病原菌小種通過AvrRppC氨基酸突變的方式來逃避RppC的識別。Chen 等[28]用K22 和丹340 構建的F6:7分離群體和重組自交系,在10 號染色體上定位到一個主效的單顯性基因RppK,轉基因驗證確定K22 基因組中的R3為抗病基因,編碼典型的NLR 蛋白,能夠與AvrRppK互作激活植物體內的ETI 反應,從而提高植物抗病性,在抗病育種上應用潛力巨大。Wang 等[33-34]通過對京2416K 和京2416 構建的F2群體進行定位、測序分析和轉基因驗證,在10 號染色體短臂上定位并克隆到南方銹病抗性基因RppM,并且基于RppM兩個保守位點(AA576、AA817)開發了可以鑒定RppM的特異性KASP 分子標記KM23 和KM19。
3.2南方銹病抗病功能基因在玉米品種改良上的應用目前已克隆抗南方銹病基因RppC、RppK為基因與基因互作,進而觸發植物體的ETI 和PTI防御反應,提高植物的抗病性,在育種中有較高的應用價值[27-28]。RppC在商業品種中應用較廣,裕豐303、中科玉505、農大108、漢單777、登海3622、蠡玉16、蠡玉88、農大108、豫單9953 等多個主栽抗銹病品種均含有RppC基因,具有較好玉米南方銹病抗性[33];12 個浚單玉米自交系含有RppC抗病基因[35];在審定品種中遼單707、康農玉598、康農2 號、高玉14022 對玉米南方銹病有較好抗性,經檢測均含有抗病基因RppK[28],含有RppM的京2416K自交系已成為我國高抗玉米南方銹病的核心種質,為我國玉米南方銹病品種的改良提供了基礎材料,先后組配出京農科767、京科682、現代965 等玉米品種,對南方銹病具有較好抗性。
4 玉米南方銹病的防治策略
4.1挖掘抗病基因進行種質創新和抗病育種利用抗病基因進行抗病種質資源創新和品種選育是控制玉米南方銹病最經濟有效的方法。RppQ、RppP25、RppD、RppC、RppK和RppM等優良抗病基因的定位和克隆以及與抗病基因連鎖的分子標記的開發,促進了傳統育種與現代育種技術相結合,為抗病種質資源創新和品種選育、改良提供了更精準有效的方法。還應加大對含有未知抗病基因的高抗南方銹病的基因挖掘工作,加強高抗南方銹病的熱帶材料在本土優異自交系的改良和品種選育方面的應用。另外分子標記輔助育種、單倍體育種、全基因組選擇和基因編輯等現代育種技術為南方銹病抗病育種提供了一種更精確快速的途徑。常規育種和現代分子技術相結合,可加快抗病育種進程。
4.2加強抗病種質的利用南方銹病的高抗種質在國內較少,已鑒定的抗病種質多為熱帶或亞熱帶材料。對已鑒定的國內高抗南方銹病的種質可以直接利用或者進行骨干材料的改良。CIMMTY、Suwan、非洲津巴布韋等高抗南方銹病的材料不能直接利用,需多代輪回選擇改良自交系。在改良過程中,可使用分子標記輔助選擇進行背景篩選,利用與南方銹病抗病基因連鎖的分子標記進行前景篩選,結合單倍體等現代育種技術可以加快品種改良進程,但材料改良還要以接種抗病鑒定結果和配合力測定為主;改良的育種材料要在黃淮海、海南、廣西等多個生態區進行田間抗性鑒定,更有利于選出廣譜抗性的優質自交系。
4.3加強病源菌的監測和預警我國南方銹病的發生和熱帶氣旋活動路徑有關,在銹病發生前期結合氣象變化,監測病源菌的動態變化,為病情發生做出預警。使用南方銹病特異性分子標記和快速檢測體系監測南方銹病在田間的侵染動態[36];發病嚴重區域和經常發病區域可建立玉米南方銹病的病害預測模型,為病情預測提供指導[30]。加快開發和利用對南方銹病檢測準確度高的田間無損化監測工具[37],建立玉米不同生態區的南方銹病病害預測模型,在病害發生前做出預警并及時采取干預措施,可有效抑制病源菌的侵染和擴散,防止病害的大規模爆發。
4.4適期配合使用化學防控監測玉米南方銹病的侵染動態,在適宜的時期及時施用化學藥劑能顯著抑制南方銹病的發生和蔓延[38-40]。在南方銹病初發期,用250g/L 的吡唑醚菌酯乳油、125g/L 的氟環唑懸浮液、5%己唑醇、25%丙環唑和43%戊唑醇處理,可顯著抑制病害的蔓延,降低病害對植株和產量的危害程度[38,40]。大喇叭口期至抽雄期是南方銹病激發期;在大喇叭口期噴施戊唑醇懸浮劑430g/L、氟硅唑乳油400g/L、吡唑醚菌酯乳油250g/L、30%肟菌·戊唑醇懸浮劑、吡唑醚菌酯乳油250g/L 與戊唑醇懸浮劑混劑430g/L,可對南方銹病有較好的防治效果[39]。由于大喇叭口期玉米植株高大、田間郁閉,玉米病蟲害防治比較困難,建議有條件的可用植保無人機進行飛防;進行人工防治時,可以用高壓噴槍進行粗防或進行精細噴霧防治,控制病情,抑制病源菌的繁殖、擴散和傳播。
4.5加強玉米主產區致病小種的鑒定工作前人研究初步確定了我國多堆柄銹菌的主要來源,遼寧、黃淮海區域和浙江、福建的南方銹病初始菌源主要來自我國臺灣地區;廣東、廣西和海南的初始菌源主要來自菲律賓;云南、貴州等西南地區初始菌源主要來自泰國、老撾、緬甸等國家,并在境內形成病害的周年循環[41]。我國不同玉米生態區的氣候差異較大,不同區域的南方銹菌的病原小種復雜。海南菌群年度間遺傳差異較大,來源較多且不穩定;山東多堆柄銹菌遺傳多樣性豐富,說明病原小種較多;廣西、廣東、浙江、福建、海南的初始菌源相近,但廣西河池的菌株致病性最強。致病小種的更迭或病菌變異會導致推廣的抗病品種喪失部分抗性,因此加強各區域的致病小種的鑒定有助于開展針對性的抗病育種。
隨著全球氣候變暖,暴雨、臺風等災害性氣候頻發,南方銹病在我國玉米主產區的發病面積不斷擴大,后期爆發速度快,尤其對黃淮海區域和南方沿海地區的玉米產量危害嚴重。抗病育種是病害防治最綠色、經濟、有效的措施。但我國高抗南方銹病種質資源較少,且抗病種質存在一定的地域限制,因此加快抗病種質的搜集,對已鑒定出的高抗種質加以利用,建立起科研院所和育種企業自己的核心抗病自交系尤為重要。深度挖掘南方銹病抗病基因,利用已開發的分子標記和前沿分子育種技術培育抗南方銹病品種。推廣部門重點推廣抗性品種,同時在抗病品種不多的情況下,動態監測病害的變化,及時有效地采取化學防控,多措并舉,以保證玉米主產區的安全生產。
