基于CRISPR-Cas9技術構建TET2基因敲除的雞胚成纖維細胞系
2023-11-22 11:07:28王強州潘詩雨方夢雅李微王佳興劉茵茵陳世豪
中國農業科技導報 2023年11期
王強州, 潘詩雨, 方夢雅, 李微, 王佳興, 劉茵茵, 陳世豪*
(1.揚州大學表觀遺傳學與表觀基因組學研究所,江蘇 揚州 225009; 2.揚州大學獸醫學院,江蘇 揚州 225009; 3.江蘇省家禽科學研究所,江蘇 揚州 225125)
甲基胞嘧啶雙加氧酶2 (tet methylcytosine dioxygenase 2,TET2)是雙加氧酶家族的成員之一,作為細胞中重要的DNA 去甲基化酶,催化5-甲基胞嘧啶(5mC)轉化為5-羥甲基胞嘧啶(5hmC),在細胞基因組表觀遺傳學中起著重要調控作用[1-2]。近年來,TET2在免疫應答和炎癥調控中的作用逐漸被認識,成為免疫學研究熱點之一。TET2 對于T 細胞和B 細胞免疫穩態的維持以及相關免疫因子的激活具有重要的調控作用[3-5]。TET2作為啟動天然免疫應答的關鍵分子,在漿細胞樣樹突狀細胞中被CXXC5 招募至TLR7/9 啟動子并促進轉錄,啟動一系列免疫級聯反應,激活天然免疫應答[6]。小鼠骨髓來源樹突狀細胞中敲除TET2 蛋白,其Toll-like 受體信號傳遞受阻[7],表明TET2與Toll-like受體信號通路密切相關。本團隊前期研究證實,DNA 甲基化參與調控雞的抗病毒天然免疫反應[8],而雞TET2 作為去甲基化過程的關鍵蛋白,對天然免疫的調節作用很大程度上未知。雞胚成纖維細胞系DF-1 是自發永生化形成的細胞系[9],是研究雞天然免疫調控機制的理想細胞系。因此,構建穩定的TET2敲除的雞胚成纖維細胞系,可為進一步研究雞TET2在天然免疫反應中的調控作用提供可靠的體外細胞模型。
CRISPR-Cas9 是新一代基因編輯技術,通過人工設計的向導sgRNA,引導Cas9 核酸酶在基因組特定位點進行靶向編輯,包括基因敲除(knockout)、敲入(knockin)[10-11]。該技術已廣泛應用于細胞敲除、基因篩選、基因治療、分子育種等領域,尤其在構建敲除細胞或敲除動物模型方面具有不可替代的作用,也為動植物的分子育種提供了新的技術與思路[12-13]。本研究利用CRISPRCas9 技術構建敲除TET2基因的雞胚成纖維細胞系,通過靶向干預TET2基因的表達,為進一步探索雞TET2在天然免疫激活和消退中的功能及分子機制奠定基礎。
1 材料與方法
1.1 試驗材料
1.1.1細胞和質粒 雞胚成纖維細胞系DF-1 源于美國ATCC,由本課題組傳代保存。非病毒載體Cas9-T2A-GFP 由本課題組通過MLM3613 Cas9質粒(Addgene,#42251)改造而來,攜帶綠色熒光蛋白(green fluorescent protein,GFP) 標簽[11]。pEX-A-U6-sgRNA購自Addgene(#65626)。
1.1.2試劑 DMEM 高糖培養基、胎牛血清購自美國Gibco 公司;DL2000 DNA Marker、2×Phanta Max Master Mix (Dye Plus)、FastPure Gel DNA Extraction Mini Kit、FastPure Cell/Tissue DNA Isolation Mini Kit 購自諾唯贊生物科技股份有限公司(南京);DH5α 感受態細胞,ExpressCast PAGE 彩色凝膠快速試劑盒和化學發光試劑盒均購自新賽美生物科技有限公司;Triton X-100 和Proteinase K(20 mg·mL-1)購自碧云天生物技術有限公司;亞甲基藍染色液(0.2%)購自北京索萊寶科技有限公司,用ddH2O 稀釋至0.02%;無內毒素質粒提取試劑盒Endo Free Plasmid Midi Kit 購自OMEGA 公司;BbsI限制性內切酶、T7核酸內切酶I (T7E I)購自NEB 公司;T4 連接酶購自Thermo Scientific 公司;HighGene 轉染試劑購自ABclonal公司;尼龍膜(正電荷)購自Whatman 生物公司;兔源TET2 多克隆抗體購自CST 公司;兔源Actin多克隆抗體購自ABclonal公司;兔源5hmC多克隆抗體購自Active Motif 生物公司;HRP 標記親和純化羊抗兔IgG購自Abcam公司。
1.2 試驗方法
1.2.1靶向雞TET2 基因的sgRNA 設計 通過NCBI 在線數據庫檢索雞TET2基因組序列,NCBI登錄號為NM_001277794.3,其另一個剪切變體登錄號為XM_015276223.3。在2 個mRNA 的共有序列設計sgRNA,即把NM_001277794.3序列的第2 個外顯子,XM_015276223.3 序列的第3 個外顯子作為靶標設計區域。利用sgRNA 在線設計工具(https://chopchop.cbu.uib.no/),按照CRISPRCas9靶點設計原則,設計4對針對TET2CDS 序列的sgRNA,且預測其在基因組內無其他靶點。在靶序列正義鏈的5’端前部添加caccg,同時合成sgRNA互補的寡核苷酸,并在5’端添加aaac,3’端添加c,合成的正反向oligo DNA序列見表1。

表1 sgRNA寡核苷酸序列Table 1 Sequences of sgRNA oligonucleotide
同時依據目的基因的序列,利用NCBI引物設計工具設計CRISPR-Cas9 敲除鑒定引物,并由蘇州金唯智生物科技公司合成。鑒定引物序列如下:TET2-ck1-F:CAATCTGCTATCCTGAGTGA和TET2-ck1-R:ATTGGAGATGCTTGGTGTT,用于檢測sgRNA-1 和sgRNA-2 位點;TET2-ck2-F:GCAGCACCCAGAAGAAAT和TET2-ck2-R:AGCAGGGAATCCATCTTTG 用于檢測sgRNA-3和sgRNA-4位點。
1.2.2靶向雞TET2 的sgRNA 載體的構建 將合成的4 對反向互補的sgRNA oligo DNA 單鏈退火,形成帶有粘性末端的短雙鏈DNA。退火步驟如下,將正反向sgRNA 序列以100 mmol·L-1溶于ddH2O,配制以下體系:1 μL Forward oligo,1 μL Reverse oligo,2 μL 10× NEB buffer 2,16 μL ddH2O,置于100 ℃水,自然降溫至25 ℃左右即可。
將短雙鏈DNA 與經BbsI 酶切后的質粒pEXA-U6-sgRNA 連接,并將連接產物轉化至DH5α 感受態細胞,涂菌板后置于37 ℃過夜培養。挑取單克隆菌落,送蘇州金唯智生物科技公司測序鑒定,顯示4 對sgRNA 序列均正確插入sgRNA 表達質粒,提示載體構建成功,提取質粒備用。
1.2.3sgRNA 切割效率的測定 DF-1 細胞鋪于6 孔板,待生長至80%密度,每個孔分別轉染4 μg的sgRNA 和Cas9-T2A-GFP 2 μg,不轉染的細胞作為對照,轉染6 h 后換成含10% FBS 的DMEM 高糖完全培養基,48 h 后收獲細胞,利用FastPure Cell/Tissue DNA Isolation Mini Kit 提取基因組DNA。
分別以TET2-ck1 和TET2-ck2 上下游引物,PCR擴增sgRNA-1、sgRNA-2和sgRNA-3、sgRNA-4靶位點附近的DNA 片段。PCR 反應體系為50 μL:5× SF Buffer 10 μL,Phanta Super-Fidelity DNA Polymerase 1 μL,dNTP Mix (10 mmol·L-1each) 1 μL,上、下游引物各2 μL,模板DNA 2 μL,無酶水32 μL。PCR 擴增條件:95 ℃預變性30 s;95 ℃變性10 s,58 ℃退火10 s,72 ℃延伸15 s,30 個循環;72 ℃充分延伸5 min 后,進行退火。退火條件:95 ℃變性5 min,以0.1 ℃·s-1的速率降溫至25 ℃。將退火后的PCR 產物進行純化,用T7E1 酶37 ℃酶切2 h,其產物用2 %瓊脂糖凝膠電泳,分析不同位點的sgRNA 引導Cas9 蛋白的切割效率。
1.2.4微量DNA 提取 待96 孔細胞長至80%~90%融合度,用胰酶消化并重懸細胞,一半繼續傳代培養,另一半用于DNA 提取,鑒定4 條sgRNA的切割效率。配制微量DNA 提取液:0.01 mol·L-1Tris pH 8,0.002 mol·L-1EDTA,0.2% Triton X-100,200 μg·mL-1蛋白酶K。
2 500 r·min-1離心收集微量細胞(>1 000個),加入50 μL 微量DNA 提取液,充分渦旋后置于56 ℃孵育3 h。孵育結束后,將樣本煮沸10 min用于滅活蛋白酶K。短暫離心,將樣本收集至管底,凍存至-20 ℃。在50 μL PCR 體系中,取10 μL 作為DNA模板。
1.2.5單克隆敲除細胞株篩選 選用Cas9 蛋白切割活性較高的靶點的sgRNA,用于制備TET2敲除的雞胚成纖維細胞系DF-1。將DF-1 細胞鋪于6 cm 細胞培養皿,培養24 h 至細胞生長至70%融合度,按說明書進行轉染。在滅菌EP 管中加入sgRNA 質粒4 μg 和Cas9-T2A-GFP 質粒2 μg,與轉染試劑混合均勻,室溫孵育10 min。將混合液逐滴加至培養皿中,37 ℃培養6 h。棄去培養基換成含10% FBS 的DMEM 高糖完全培養基,繼續培養48 h。用胰酶消化轉染48 h 后的DF-1 細胞,制備細胞懸液進行流式細胞分選,以獲得高表達GFP 基因的細胞。將獲得的GFP 陽性細胞通過有限稀釋法,在96 孔板中培養單細胞克隆,待亞克隆的細胞密度生長至80%時,將一半細胞用于提取DNA,另一半細胞繼續擴大培養。采用微量DNA 提取法,通過PCR 擴增靶位點序列,送至蘇州金唯智生物有限公司進行DNA 測序鑒定。
1.2.6Western blot 分析蛋白水平 將雞TET2敲除的DF-1細胞系與野生型DF-1細胞系分別鋪至6孔細胞板,待細胞密度長至90%左右時收獲細胞,裂解獲得蛋白,進行Western blot 分析。簡要步驟如下:用預冷PBS洗滌細胞沉淀,加入RIPA裂解液進行細胞裂解,冰上孵育30 min;4 ℃,12 000 r·min-1離心15 min。吸上清,通過BCA法測定蛋白含量。然后加入5×SDS-PAGE蛋白上樣緩沖液,99 ℃金屬浴加熱5 min,充分變性蛋白后,進行SDS-PAGE電泳。230 mA 恒流條件下轉膜2.5 h。將膜用5%脫脂牛奶封閉緩沖液室溫封閉30 min。稀釋TET2(1∶1 000)和ACTIN (1∶2 000) 抗體,4 ℃孵育過夜。第2天,洗膜3 次后,用HRP 標記親和純化羊抗兔IgG(1∶10 000)室溫孵育1 h 后,用TBST 緩沖液洗滌3 次,每次5 min。最后置于蛋白印跡成像系統下觀察條帶。
1.2.7Dot blot分析方法 將雞TET2敲除的DF-1細胞系與野生型DF-1 細胞系分別鋪至6 孔細胞板,待細胞密度長至90 %左右時,收獲細胞,提取基因組DNA,進行Dot blot 分析。基因組DNA 提取方法按照諾維贊FastPure Cell/Tissue DNA Isolation Mini Kit說明書進行。
將DNA 梯度稀釋到200、100、50 μg·μL-1,99 ℃變性5 min后置于冰上迅速冷卻,放置10 min。然后,將不同含量水平的DNA各上樣2 μL到尼龍膜上,室溫干燥15 min后,置于80 ℃烘箱加熱固定1 h。將固定后的尼龍膜用5 %脫脂牛奶封閉緩沖液室溫封閉30 min。稀釋5 hmC(1∶1 000)抗體,置于4 ℃孵育過夜。第2 天,用TBST 緩沖液洗膜3 次后,室溫下用HRP 標記親和純化羊抗兔IgG(1∶10 000)室溫孵育1 h,再用TBST洗滌3次,每次5 min。最后置于蛋白印跡成像系統拍照。拍照完成后,尼龍膜用TBST 緩沖液洗滌2 次后,置于0.02%亞甲基藍染色液染色K,用水漂洗后通過普通相機進行拍照,獲得DNA上樣內參。
2 結果與分析
2.1 靶向雞TET2的sgRNA切割效率的測定
瓊脂糖凝膠電泳觀察目的基因片段被T7E1酶切割情況(圖1),結果可知sgRNA-1、sgRNA-2、sgRNA-3 出現被T7E1 酶切割的小片段,說明轉染這些sgRNA 后細胞基因組DNA 發生了編輯事件。其中,設計的sgRNA-2 引導Cas9 蛋白對靶標序列表現出較高切割活性,經T7E1酶切后的全長擴增片段(520 bp)產生329和191 bp的片段,大小符合預期。由此,后續篩選單克隆細胞選取sgRNA-2進行轉染。
2.2 篩選單克隆敲除細胞株和測序鑒定結果分析
將篩選的單克隆細胞株提取基因組DNA,經PCR 擴增后進行Sanger 測序,測序結果與原始基因組序列進行比對,確定具體突變位置及其突變序列(圖2),發現有1 株細胞(命名為DT-6)發生了堿基的缺失突變。TET2-sgRNA-2 切割位點如圖2 所示,DNA 雙鏈分別發生了2 和28 bp 的缺失,導致雞TET2基因移碼突變,轉錄提前終止,推測該DT-6細胞中TET2蛋白將失活。
2.3 TET2敲除細胞形態學變化分析
由圖3 可知,與正常DF-1 細胞相比,TET2基因敲除的DT-6 細胞在形態上呈現較為明顯的差別,表現為形態多樣,較為飽滿,呈現無規則的菱形或雙錐形,原因可能是TET2敲除導致某些骨架蛋白或黏附蛋白等的表達受到影響。
2.4 敲除細胞TET2 蛋白水平分析
敲除細胞擴大培養后,進行Western blot 驗證。結果如圖4 所示,相對于DF-1 細胞,敲除細胞系DT-6 中TET2 蛋白幾乎不表達,說明敲除TET2 的DF-1 細胞系構建成功。
2.5 敲除細胞羥甲基化水平分析
TET2 維持細胞的羥甲基化(5hmC)水平,可以通過調控基因的羥甲基化水平控制相關基因的表達。本研究利用Dot blot 分析穩定敲除細胞的羥甲基化水平,結果如圖5 所示,相對于正常DF-1 細胞,敲除細胞系DT-6 中5hmC 水平顯著下降。由此可見,TET2 是維持DF-1 細胞全基因組5hmC 水平的關鍵蛋白。
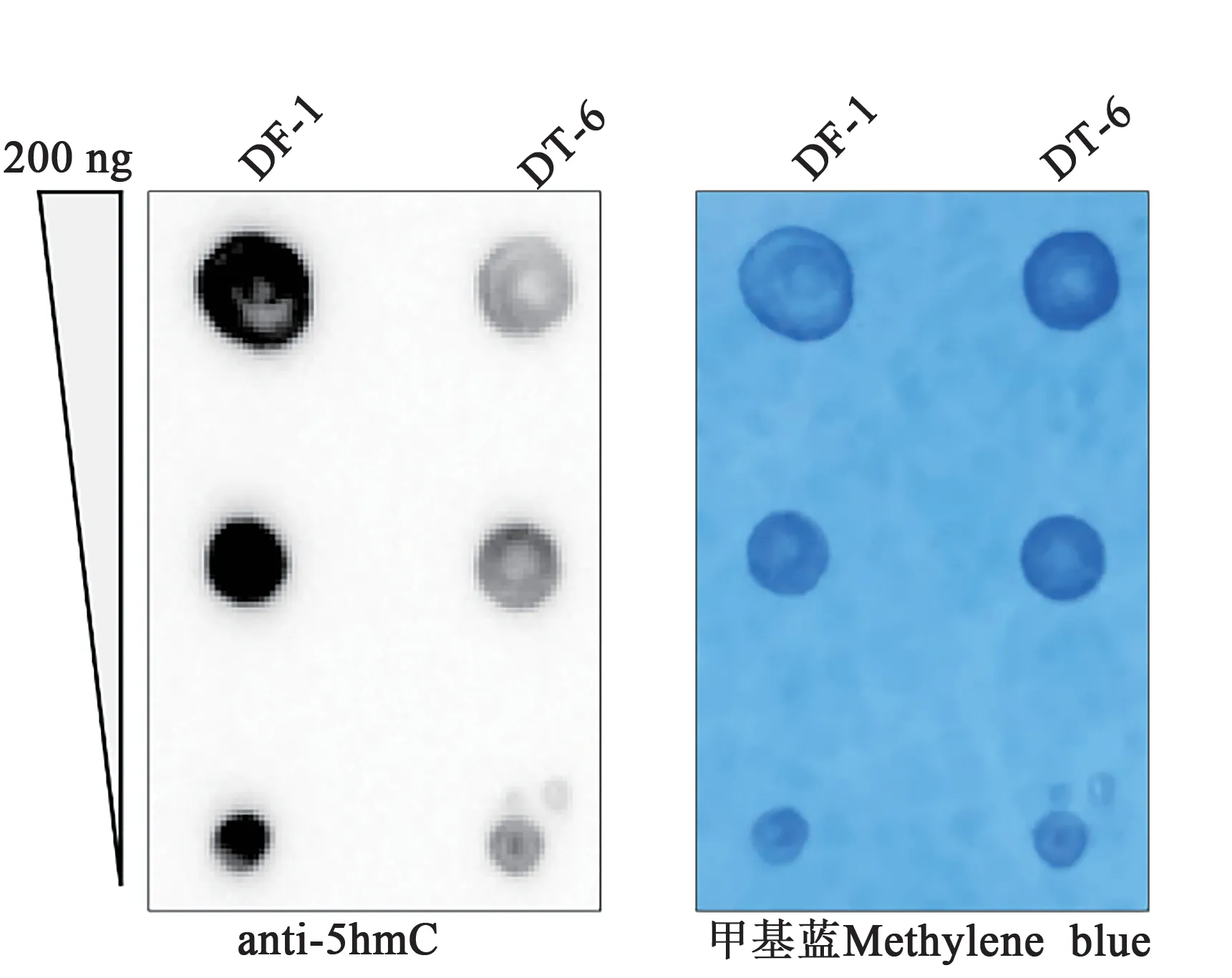
圖5 Dot blot檢測野生型和TET2敲除細胞的羥甲基化水平Fig. 5 Dot blot detected levels of hydroxymethylation(5hmC) in wild type and TET2 knockout cells
3 討論
已有研究表明,DNA甲基化抑制劑顯著增強雞胚成纖維細胞的抗病毒天然免疫反應[9],表明DNA甲基化修飾參與調控雞天然免疫反應的激活。而羥甲基化酶TET2 是一種內源性的DNA 去甲基化酶,對于調控DNA甲基化的平衡具有重要作用[14-15]。但TET2基因及其介導的羥甲基化修飾在雞抗病毒天然免疫反應中的功能及其作用機制尚未明確。
CRISPR-Cas9 是新一代基因編輯技術,通過人工設計的sgRNA 來識別目的基因組序列,并引導Cas9 蛋白酶在原間隔序列臨近基序(protospacer adjacent motif, PAM)區上游的3~8 堿基處切割目的DNA,形成雙鏈斷裂,從而啟動細胞內的DNA 損傷修復機制,造成基因敲除或敲入,最終達到對基因組DNA 進行編輯的目的[16-17]。與上一代基因編輯技術,如鋅指核酸酶(zincfinger nucleases,ZFN)和類轉錄激活因子效應分子核酸酶(transcriptionactivation-like effector nucleases,TALEN)相比,CRISPR-Cas9 基因編輯技術因操作簡單、編輯效率高、試驗周期短等特點被廣泛用于構建各種細胞系和各種用途的動物疾病模型[17-18]。應用CRISPR-Cas9 技術構建基因敲除的細胞也是禽類基因功能研究的重要手段之一。例如通過構建雞TBK1 敲除的DF-1 細胞,分析了雞TBK1在抗病毒信號通路中的功能[19]。
雞TET2基因位于4號染色體,由11個外顯子和10 個內含子組成,該基因也存在另一種剪接變體,由13個外顯子和12個內含子組成。sgRNA的設計普遍共識是選擇靶基因的第1、第2外顯子區域,為保證能夠將雞TET2蛋白徹底敲除失活,本研究選擇在雞TET2 基因的CDS 區的共有序列進行sgRNA 靶點設計,與預期結果一致,該位點sgRNA可以完全敲除TET2基因。本研究還發現,TET2敲除后雞成纖維細胞發生了明顯的形態變化。研究表明,TET2調控諸多發育相關的基因[20-21],雞TET2還通過DNA去甲基化作用方式調節成肌調節因子的表達,促進雞成肌細胞的分化[22]。本研究中,TET2敲除是否影響雞胚成纖維細胞骨架蛋白等基因表達造成細胞形態的變化,還有待進一步驗證。
已有研究指出,TET2 對于T 細胞和B 細胞免疫穩態的維持以及相關免疫因子的激活具有重要的調控作用[3-6],也是樹突狀細胞啟動天然免疫應答的關鍵分子,且與Toll-like 受體信號通路密切相關[7]。上述研究表明,TET2 與免疫細胞功能密切相關,且依賴其DNA 去甲基化的方式來發揮免疫調控作用。本研究利用CRISPR-Cas9技術敲除TET2基因后,DF-1 細胞全基因組5hmC 水平顯著下調,表明雞TET2對于維持細胞基因組羥甲基化水平具有重要作用。鑒于TET2 對免疫反應的激活具有重要的作用,推斷TET2敲除后將顯著影響雞胚成纖維細胞天然免疫反應的激活。
綜上所述,本研究在未引入抗性篩選基因的條件下獲得了TET2基因敲除的單克隆細胞株,不必考慮抗性基因的插入對目標基因功能的影響,為其他類型細胞的篩選提供了適合的方法。本研究應用CRISPR- Cas9 技術成功篩選獲得TET2基因敲除的雞胚成纖維細胞系(DT-6),為研究雞TET2的生物學功能提供可靠的體外模型,同時為在其他雞細胞系TET2的敲除提供了有效靶點。
