溶桿菌中活性天然產物的研究進展
2023-11-23 09:11:12周閃閃黃遠龍黃建忠李善仁
生物技術通報 2023年10期
周閃閃 黃遠龍 黃建忠 李善仁
(福建師范大學生命科學學院 工業微生物發酵技術國家地方聯合工程研究中心,福州 350117)
由于抗生素的濫用或過度使用,導致耐藥病原體不斷蔓延,嚴重威脅患者的健康與生命,因此尋找新型天然產物藥物迫在眉睫。近年來,隨著高通量基因測序技術的進步,越來越多的微生物基因組被測序,發現除了傳統藥物產生菌放線菌和真菌外,許多以前被忽視的微生物中也含有大量次級代謝產物生物合成基因簇,具有合成新型天然產物的潛力[1-3]。因此,在后抗生素時代,這些研究較少或尚未開發的微生物正成為新型藥物發現的重要來源。
溶桿菌是一類具有滑行運動的革蘭氏陰性細菌,對植物病原真菌、細菌、卵菌和線蟲等具有較強的拮抗作用,作為一種新型的生物防治資源而被廣泛研究[4]。溶桿菌以溶解其他微生物細胞而得名,除了能產生多種胞外水解酶外(如幾丁質酶、β-1,3?葡聚糖酶、蛋白酶等),溶桿菌還能合成多種活性次級代謝產物,在農業生產和藥物開發中具有巨大的應用潛力[5]。本文從溶桿菌屬細菌資源的多樣性、溶桿菌中次級代謝產物種類和應用等方面進行總結和展望,以期從自然環境中發現更多的溶桿菌資源,為發現更多具有新穎結構和生物活性的天然產物藥物奠定研究基礎。
1 溶桿菌資源環境分布的多樣性
1978年,加拿大科學家Christensen和Cook在伯杰氏手冊中首次建立了一個新的屬溶桿菌屬(Lysobacter),根據模式菌株特征命名和描述了4個新種即抗生素溶桿菌(L. antibioticus)、變棕溶桿菌(L.brunescens)、產酶溶桿菌(L. enzymogenes)和膠狀溶桿菌(L. gummosus)以及1個亞種即產酶溶桿菌庫克亞種(L. enzymogenes subsp. cookii)。溶桿菌屬被放置在新的目溶桿菌目(Lysobacterales)新的科溶桿菌科(Lysobacteraceae)內[4]。2015年,Naushad等[6]根據已鑒定的保守簽名插入/刪除(conserved signature inserts/deletions, CSIs)基因序列和系統發育分析,修改了溶桿菌目(Lysobacterales)和溶桿菌科(Lysobacteraceae)為黃單胞菌目(Xanthomonadales)和黃單胞菌科(Xanthomonadaceae),兩者是早期的同義詞。近年來溶桿菌新種不斷被發現,截至2023年6月,在原核生物標準命名列表網站(LPSN)中收錄的溶桿菌已達93種(https://www.bacterio.net/genus/lysobacter),顯示出溶桿菌屬細菌具有豐富的多樣性。
溶桿菌屬細菌在陸地土壤,特別在農作物根際土壤中廣泛存在,大部分溶桿菌對植物病原菌具有廣譜的拮抗作用,表明溶桿菌在維持土壤生態平衡方面起重要作用[7]。在淡水、海水和南極洲海岸沉積物中也分離得到新的溶桿菌,表明溶桿菌對極端環境也具有較強的適應能力[8-9]。在動植物體中,如西紅柿莖[10]、海綿樣品[11]、南極洲企鵝羽毛[12]、大熊貓養殖圈舍[13]、太平洋對蝦腸道[14]、蟒蛇呼吸道[15]、人瞼板腺分泌物[16]、東方白鸛糞便[17]、牛糞堆積物[18]等樣品中也發現溶桿菌新種,表明溶桿菌在動植物體中也有廣泛的存在。綜上可知,溶桿菌資源在自然界環境中廣泛分布,不僅在種屬上具有多樣性,對各種環境也有廣泛的適應性。
2 溶桿菌中活性天然產物
溶桿菌產生的次級代謝產物主要有聚酮類、非核糖體肽類、聚酮-非核糖體肽雜合類和其他小分子等化合物,在化學結構和作用機制上獨具特色,在農業和醫藥上顯示出極大的應用價值。
2.1 聚酮類天然產物
溶桿菌中聚酮類天然產物相對較少,從蜥蜴表皮上分離的L. gummosus產生III型聚酮化合物2,4?二乙酰基間苯三酚,具有抗細菌和真菌的活性,可以為蜥蜴皮膚提供保護屏障[19]。通過序列分析,溶桿菌基因組中含有聚酮合酶生物合成基因簇,但發現的聚酮化合物較少,可能與聚酮合酶基因簇在溶桿菌中沉默不表達有關[20]。因此,激活溶桿菌中沉默聚酮合酶基因簇的表達,有助于發現新型聚酮化合物。
2.2 非核糖體肽類天然產物
溶桿菌中次級代謝產物以非核糖體肽類為主,所以溶桿菌又被稱為“肽合成專家”[3]。目前從溶桿菌中發現的非核糖體肽類化合物有7個,包括3個八肽化合物plusbacins、tripropeptins和hypeptin;2個十一肽化合物lysobactin和WBP?29479A1;2個十二肽化合物WAP?8294A和lysocin(圖1)。溶桿菌來源的非核糖體肽類化合物具有結構新和活性強的特點,對臨床耐藥細菌展示出良好的抗菌效果。
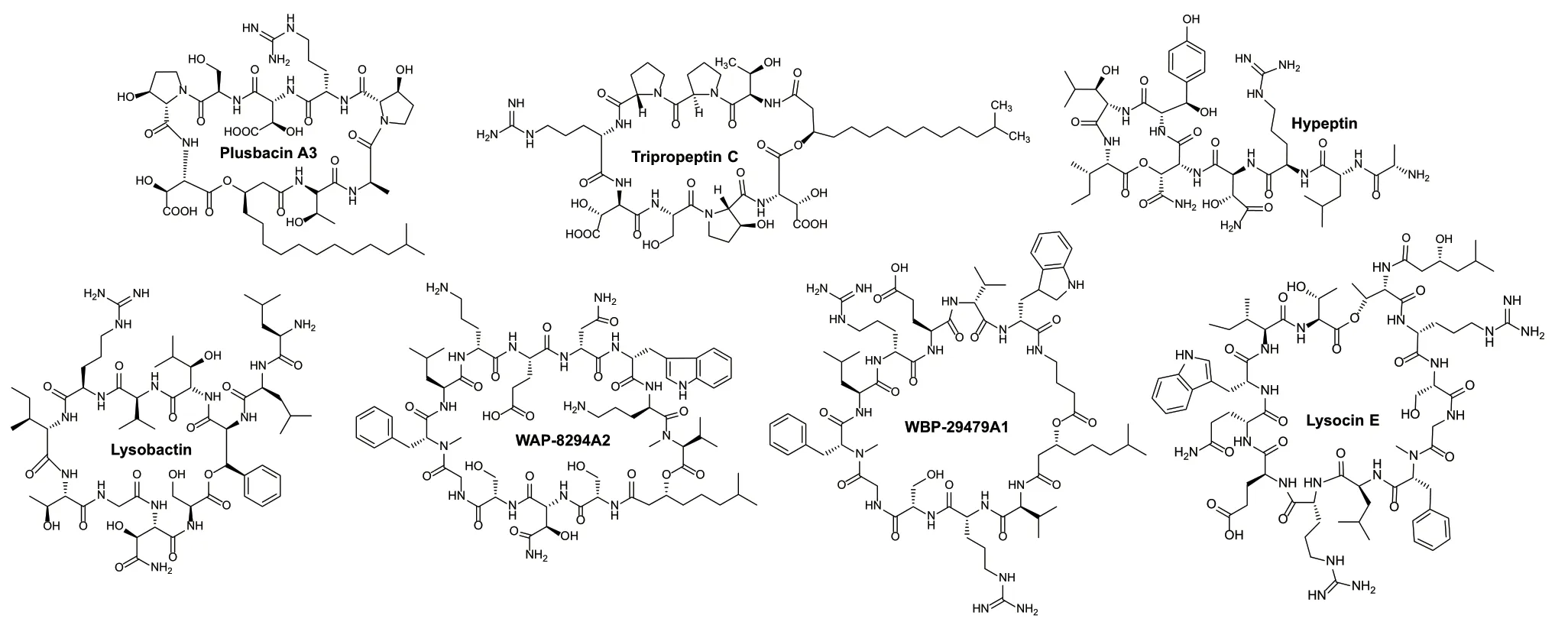
圖1 溶桿菌中非核糖體肽類化學結構Fig. 1 Chemical structures of nonribosomal peptides isolated from Lysobacter spp.
八肽化合物plusbacins是1991年從假單胞菌PB?6250(2016年被重新鑒定為溶桿菌)分離得到8個含有不同側鏈的環脂肽化合物,其中plusbacin A3活性最好,對耐甲氧西林金黃色葡萄球菌(MRSA)和耐萬古霉素腸球菌(VRE)最小抑菌濃度分別為0.78 μg/mL和3.13 μg/mL[21-22]。Tripropeptins是2001年從溶桿菌BMK333?48F3中分離得到6個環脂肽化合物,差別在于脂肪酰基側鏈不同,側鏈越長,抑菌活性越好[23]。Tripropeptins和plusbacins結構相似,僅第2個組成氨基酸不同,其作用機制是通過與焦磷酸十一烯丙基酯形成復合物,阻斷細胞壁生物合成的脂質循環,這不同于已知藥物如萬古霉素和達托霉素的作用機制[24]。Hypeptin是2021年從溶桿菌K5869中分離得到一個環八肽化合物,和其他環肽相比沒有脂肪酰基側鏈,對MRSA最小抑菌濃度為0.25 μg/mL,其抗菌機制是通過與多個十一異戊二烯基磷酸的生物合成中間體形成2∶1復合物,阻止細菌細胞壁的生物合成[25]。
十一肽化合物lysobactin是1988年從溶桿菌ATCC53042分離得到的活性環肽,沒有脂肪酰基側鏈,對MRSA和VRE最小抑菌濃度分別為0.39 μg/mL和0.78 μg/mL,比萬古霉素分別低2倍和50倍,主要抑制細菌肽聚糖合成,lysobactin與肽聚糖合成前體脂質II形成1∶1復合物,這與其他環肽的作用機制也不同[26]。最新研究發現lysobactin對膿腫分枝桿菌(Mycobacterium abscessus)具有良好的抗菌活性,在抗結核病方面具有應用潛力[27]。WBP?29479A1是2019年通過基因組挖掘的方法從抗生素溶桿菌ATCC29479中發現的環脂肽化合物,其結構含有11個氨基酸,其中6個為非蛋白氨基酸,WBP?29479A1對不同MRSA最小抑菌濃度范圍在0.25-2 μg/mL之間,作用機制與WAP?8294A相似[28]。
十二肽化合物WAP?8294A是1997年從溶桿菌WAP?8294 中分離得到的一組環脂肽化合物,其中WAP?8294A2對MRSA最小抑菌濃度為0.78 μg/mL,其通過選擇性作用在細菌細胞質膜中甲萘醌靶點上,干擾細菌呼吸鏈而殺菌。WAP?8294A2已經進入臨床試驗,目前正作為新的化學實體,用于治療皮膚感染、肺炎、細菌性心內膜炎和骨髓炎等[5,29]。Lysocin是2015年從溶桿菌RH2180?5中分離得到的環脂肽化合物,lysocin E對MRSA最小抑菌濃度為4 μg/mL[30]。Lysocin E對結核分枝桿菌也具有很好的體外活性,主要與甲萘醌結合,通過膜破壞和呼吸抑制等協同作用,可以在較短的時間內治愈耐藥性和潛伏期的結核病[31]。由于lysocin、WAP?8294A和WBP?29479A1在結構上具有相似的氨基酸組成,都是甲萘醌結合抗生素,毒副作用低,是具有臨床開發前景的非核糖體抗菌肽。
2.3 聚酮-非核糖體肽雜合天然產物
從溶桿菌中發現的聚酮-非核糖體肽雜合化合物主要有熱穩定抗真菌因子(heat stable antifungal factor, HSAF)、cephabacin和lysohexaenetides等3種(圖2)。HSAF最初是從產酶溶桿菌C3中分離得到一類十七元環內酰胺化合物,其生物合成由一個模塊的I型PKS重復使用合成2條獨立的聚酮鏈,在NRPS催化下與1分子鳥氨酸縮合形成含有特特拉姆酸單元的碳骨架,在4個氧化酶(OX1-OX4)的催化下合成三元環化合物,是一種獨特的PKS?NRPS生物合成途徑[32]。HSAF對植物病原真菌和卵菌具有廣譜抗菌活性,其作用機制是通過干擾真菌鞘脂的合成而抑制絲狀真菌的生長[33];在白色念珠菌中通過誘導產生活性氧而引起細胞凋亡[34]。HSAF生物合成基因簇中含有多個氧化酶基因,除了HSAF外,還產生一系列具有不同環系的衍生物,如lysobacterene、lysobacteramide、alteramide等化合物,也具有抗真菌活性和細胞毒效應[5]。HASF及其衍生物因具有新穎的結構、獨特的作用機制、對環境友好等特點,在農作物病害生物防治中已有廣泛應用。
Cephabacin是從溶桿菌YK90中分離得到的頭孢烯β-內酰胺類化合物,其結構由1個頭孢烯環、1個乙酸和4個氨基酸(Arg?Ala?Ala?Ala)組成,通過抑制肽聚糖的合成來破壞細胞壁從而發揮抑菌作用。Cephabacin生物合成基因簇中包含2個NRPS基因,1個NRPS?PKS基因和頭孢烯合成相關的基因,其生物合成途徑已基本闡明[35]。
在鏈霉菌中通過異源表達溶桿菌DSM3655中一個PKS?NRPS基因簇分離得到的線性脂肽化合物lysohexaenetides,其中lysohexaenetide A由一個不飽和聚酮鏈和4個氨基酸(Ser?His?Gly?Val)組成,不飽和聚酮鏈由單模塊的I 型PKS通過6次迭代催化合成,4個氨基酸由NRPS負責加載合成;lysohexaenetide B中僅含有兩個氨基酸殘基(Ser?His),推測可能是lysohexaenetide A的水解產物或者在生物合成中提前釋放的中間產物。在生物活性上,lysohexaenetides沒有明顯的抗菌活性和細胞毒效應,其生物學功能有待于進一步研究[36]。
2.4 其他天然產物
溶桿菌中除了由PKS和NRPS生物合成途徑合成的天然產物外,還發現一些其他類型的活性天然產物(圖3)。從溶桿菌YK422中分離得到抗細菌化合物lactivicin,結構中不含有β-內酰胺環,但卻與β-內酰胺類抗生素作用機制相同[37]。從辣椒溶桿菌AZ78中分離得到二酮哌嗪類化合物,具有抗細菌活性[38]。從抗生素溶桿菌OH13中分離得到吩嗪類化合物mycin,具有廣譜的抗細菌和抗真菌活性[39]。從抗生素溶桿菌OH13中還分離得到抗真菌化合物對氨基苯甲酸(pABA),在病原真菌的生物防治中具有較大的應用前景[40]。在產酶溶桿菌OH11中通過強啟動子置換激活沉默的NRPS基因簇表達,得到pyrrolopyrazines化合物[41]。在產酶溶桿菌OH11中通過基因組挖掘發現溶桿菌中第一個鐵載體化合物lysochelin,在鐵平衡和細胞信號方面發揮重要功能[42]。分析溶桿菌基因組序列發現,溶桿菌中還有如細菌素、二肽和核糖體肽類等次級代謝產物的生物合成基因簇,但目前尚未見有相關化合物的報道,表明溶桿菌中還有其他類型的天然產物尚待發現。
3 溶桿菌及其天然產物的應用
現代農業生產中越來越重視生物防治,利用有益微生物及其代謝產物來防治農作物病害,與化學防治相比,其最大的優點是對人畜安全和對環境友好。常見的生防細菌如芽孢桿菌和假單胞菌,已廣泛應用于農業病蟲害控制[43]。溶桿菌是新型的生防細菌,與黏細菌相似,通過群體捕食的方式對其他細菌和真菌進行捕食以獲取營養[44]。溶桿菌生防機理主要有定殖作用、抗生作用、產生生物表面活性物質和誘導寄主抗病性等[45-47]。研究發現,在產酶溶桿菌OH11中,利用非接觸的“長武器”(如擴散型抗真菌物質HSAF或水解酶)和通過接觸的“短武器”(如四型和六型分泌系統)協同工作來拮抗病原菌,表明溶桿菌具有多層次的生防機制[48-50]。
3.1 抗病原真菌
溶桿菌屬細菌是一種有效的抗植物病原真菌生物防治劑,例如產酶溶桿菌C3[51]、產酶溶桿菌3.1T8[52]、產酶溶桿菌OH11[53]、辣椒溶桿菌AZ78[54]、抗生素溶桿菌OH13[40]、抗生素溶桿菌HS124[55]等對各種病原真菌具有廣譜的拮抗作用。產酶溶桿菌C3對病原真菌如小麥根腐病菌(Bipolaris sorokiniana)、大豆灰斑病(Cercospora sojina)、大豆斑枯病(Septoria glycines)、北美大豆猝死綜合癥病菌(Fusarium virguliforme)、禾谷鐮孢菌(F.graminearum)、水稻紋枯病菌(Rhizoctonia solani)、油菜菌核病菌(Sclerotinia sclerotiorum)、芝麻莖點枯病菌(Macrophomina phaseolina)和卵菌如蘆薈根腐病菌(Pythium ultimum)、大豆根腐病菌(Phytophthora sojae)等均有防治作用,主要通過溶桿菌分泌的水解酶和次級代謝產物HSAF等發揮抑菌作用[56-57]。產酶溶桿菌OH11對卵菌如大豆根腐病菌(P.sojae)、辣椒疫霉菌(P. capsici)、馬鈴薯晚疫病菌(P.infestans)等也具有很好的拮抗活性,通過抑制病原菌菌絲生長、消化子囊、抑制子囊發芽和激發植物免疫反應來保護植物免受病原體侵染[47]。溶桿菌作為生防制劑使用,不僅達到綠色防治病蟲害的作用,還能減少化學農藥的使用,使農業生產可持續發展。
產酶溶桿菌中次級代謝產物HSAF和alteramide B等衍生物,除了能抑制絲狀真菌生長外,對白色念珠菌也有很好的拮抗活性,主要通過誘導產生活性氧引起細胞凋亡[34,58]。由于HSAF及其衍生物具有獨特結構和作用機制,不同于已知的抗真菌藥物,對植物和哺乳動物安全,因此在開發新型抗真菌藥物方面具有應用潛力。
3.2 抗病原細菌
病原細菌是引起植物病害的重要病原生物,是農作物中較難防治的一類病害。產酶溶桿菌C3對植物病原細菌如番茄潰瘍病菌(Clavibacter michiganensis)具有抑菌活性[56]。抗生素溶桿菌13?6對水稻條斑病菌(X. oryzae pv. oryzicola)、青枯病菌(Ralstonia solanacearum)、胡蘿卜軟腐果膠桿菌(Pectobacterium carotovorum)、煙草野火病菌(Pseudomonas syringae pv. tabaci)等具有抑菌活性[59]。抗生素溶桿菌OH13產生的對氨基苯甲酸對大豆斑疹病菌(X. axonopodis pv. glycines)具有抑菌活性,主要通過改變外膜的完整性來抑制病原菌生長[60]。
從溶桿菌中發現的非核糖體肽類化合物如plusbacins、tripropeptins、hypeptin、lysobactin、WBP?29479A1、WAP?8294A和lysocin等對臨床耐藥細菌MRSA具有良好的抗菌活性,使溶桿菌的應用范圍從生物防治延伸到生物醫藥領域[3,5]。溶桿菌產生的WAP?8294A2(lotilibcin)在MRSA感染的小鼠體內比萬古霉素的殺菌效果高14倍,目前正作為新的化學實體用于臨床治療[5]。除此之外,溶桿菌中lysobactin、lysocin E等對結核分枝桿菌也具有顯著的抗菌活性,可以開發抗結核先導化合物[27,31]。
3.3 抗線蟲
植物線蟲是導致植物病害的四大病原之一,危害幾乎所有的糧食和經濟作物。產酶溶桿菌C3對秀麗隱桿線蟲(Caenorhabditis elegans)、甜菜胞囊線蟲(Heterodera schachtii)、爪哇根結線蟲(Meloidogyne javanica)、穿刺根腐線蟲(Pratylenchus penetrans)和草莓芽葉線蟲(Aphelenchoides fragariae)等線蟲卵的孵化具有抑制作用,可能是因為溶桿菌分泌的幾丁質酶水解卵殼中幾丁質層,破壞線蟲卵的完整性[61]。抗生素溶桿菌HS124可以產生胞外水解酶和4?羥基苯乙酸,對南方根結線蟲(M. incognita)卵的孵化具有抑制作用,對 J2期線蟲具有致死作用,在田間實驗中有較好的防治效果,可進一步開發根結線蟲的生物防治劑[62]。辣椒溶桿菌YS1215對南方根結線蟲具有生物防治作用,通過柱層析法從其發酵液中鑒定出活性物質為乳酸,可以作為化學殺蟲劑的替代品來防治根結線蟲[63]。產酶溶桿菌B25對番茄植物南方根結線蟲和爪哇根結線蟲(M.javanica),顯示出50%-95%的抑制活性和化學防治劑作用相當,是開發可持續殺線蟲產品的良好候選者[64]。
4 總結與展望
溶桿菌在陸生、水生和動植物體環境中廣泛分布,表明溶桿菌具有強大環境適應能力。溶桿菌作為其他微生物的“捕食者”,在土壤微生物生態系統平衡中發揮重要作用。相對于研究較多的芽孢桿菌和假單胞菌等生防細菌,溶桿菌是一類尚未完全開發的生防細菌。雖然溶桿菌屬新種不斷被發現,但自然界中還存在豐富的溶桿菌資源尚待開發和利用。
溶桿菌中活性天然產物種類主要有聚酮類、非核糖體肽類、聚酮-非核糖體肽雜合類以及其他活性小分子化合物,具有化學結構多樣、生物活性顯著和作用機制新穎等特點,在藥物開發方面具有巨大的潛力。抗真菌代謝產物以HSAF及其衍生物為主,具有活性強、抗菌譜廣、穩定性好等優點,在農作物真菌病害防治上具有較好的應用效果,是化學農藥的良好替代品;抗細菌代謝產物以非核糖體肽類為主,在氨基酸組成上具有豐富的多樣性,不僅對植物病原細菌有拮抗作用,對臨床耐藥細菌MRSA和結核分枝桿菌等均具有顯著的抗菌活性,因此在生防制劑和抗耐藥細菌藥物等方面有良好的開發前景。
雖然溶桿菌及其天然產物研究取得了一定成果,但同時也存在一些問題:一是溶桿菌中發現的化合物數量還不夠多,大量新化合物尚待發掘。后續研究一方面可以擴大溶桿菌資源篩選范圍,分離更多溶桿菌,通過構建溶桿菌資源庫,系統篩選和分離溶桿菌中的活性天然產物;另一方面可以利用基因組挖掘技術從現有的溶桿菌資源中挖掘天然產物的合成潛力,例如激活溶桿菌中沉默基因簇表達發現新的天然產物;二是溶桿菌中活性化合物產量低,限制了其大量生產和應用。后續研究中一方面可以通過對野生型溶桿菌進行遺傳改造或發酵工藝改進,提高活性化合物的產量;另一方面可以通過基因簇異源表達,在遺傳背景清楚的底盤宿主中優化表達,提高目標化合物產量,深入研究其生物合成、作用機制以及進一步運用合成生物學改造代謝途徑,實現工業化生產。
綜上所述,溶桿菌資源在自然界廣泛分布,具有豐富的多樣性,在維持土壤生態平衡中發揮重要作用。溶桿菌是新型生防細菌,也是新型抗生素發現的重要來源,在農業和醫藥上具有巨大的應用潛力。隨著人們對溶桿菌資源及其天然產物研究的不斷深入,相信會有更多、活性更強的天然產物被發現,為生物防治和生物醫藥的發展提供豐富的先導化合物。
