福建沿海巨蠣屬(Crassostrea)牡蠣的種類及其分布*
2023-12-06 07:38:18于詩奇韓自強陳燕婷郭團玉闕華勇
海洋與湖沼 2023年6期
于詩奇 韓自強 陳燕婷 郭團玉 闕華勇
福建沿海巨蠣屬()牡蠣的種類及其分布*
于詩奇1韓自強1陳燕婷2郭團玉3闕華勇1①
(1. 集美大學水產學院 農業農村部東海海水健康養殖重點實驗室 福建省水產生物育種與健康養殖工程研究中心 福建廈門 361021; 2. 福建省水產技術推廣總站 福建福州 350002; 3. 廈門海洋職業技術學院海洋生物學院 福建廈門 361100)
福建省位于我國東南沿海, 海岸線曲折, 港灣眾多, 有多條河流入海, 適宜牡蠣棲息繁衍, 是我國牡蠣資源最豐富的地區之一。迄今關于福建沿海牡蠣種類組成及分布情況的研究報道尚少, 鑒于近20余年福建主要海灣經歷了高強度的牡蠣養殖, 有必要了解福建沿海牡蠣自然群體的物種組成和數量占比等自然種質資源狀況。基于現場調查取樣, 結合采用多重種特異性PCR、ITS2雜交種鑒定和CO I測序共同鑒定福建沿海巨蠣屬()牡蠣種類及其分布情況。研究結果表明, 從福建沿海由北至南牡蠣野生種苗海區的19個采樣點共960個樣品中發現3種巨蠣屬牡蠣, 分別是福建牡蠣()、熊本牡蠣()和香港牡蠣(), 并未發現以往報道的近江牡蠣()。其中福建牡蠣(607個)占63.23%、熊本牡蠣(343個)占35.73%、香港牡蠣(10個)占1.04%。除個別采樣點外, 福建牡蠣和熊本牡蠣在各采樣海區均有分布。香港牡蠣僅在泉州灣河口濕地自然保護區(新發現群體)有少量分布。研究結果揭示的福建沿海地區巨蠣屬的物種種類及其分布為后續開展牡蠣遺傳多樣性分析和遺傳分化等種群遺傳研究提供了關鍵基礎資料, 為牡蠣種質資源的保護和利用打下重要基礎。
牡蠣種類; 巨蠣屬; 福建沿海; 多重種特異性PCR; 基于ITS2序列的雜交種鑒定; COI測序
牡蠣屬于軟體動物門(Mollusca)、雙殼綱(Bivalvia)、珍珠貝目(Pterioida)、牡蠣科(Ostreidae), 在我國從北到南沿海均有分布, 其肉味鮮美且具有很高的營養價值。目前在我國得到規模化養殖的牡蠣均來自巨蠣屬(), 包括長牡蠣()、福建牡蠣()、香港牡蠣()和熊本牡蠣() (Peng, 2021)。2022中國漁業統計年鑒中福建省牡蠣養殖為211萬t, 占全國總產量的36.29%, 位居全國第一(農業農村部漁業漁政管理局等, 2022)。
對物種進行準確鑒定是開展生物學研究和種質資源保護的基礎(李琪等, 2021)。由于牡蠣具有廣棲性和表型高可塑性等特點, 僅依靠形態學和解剖學手段無法對其進行有效的鑒定(李孝緒等, 1994; 闕華勇等, 2003)。目前應用于牡蠣物種鑒定的分子手段包括: PCR-RFLP鑒定(Cordes, 2008; Li, 2015)、線粒體CO I基因測序(Wang, 2004; Xia, 2009; Liu, 2011; Liu, 2021)、多重種特異性PCR (multiplex species-specific PCR) (Wang, 2008a)和HRM (Wang, 2014; Xu, 2014)等。其中, 線粒體CO I測序是目前牡蠣鑒定最為準確且運用較為廣泛的方法, 但需要具備測序條件和一定費用; 多重種特異性PCR方法可通過凝膠電泳快速準確地鑒定出巨蠣屬牡蠣的物種, 適用于數量較多的牡蠣樣本的鑒定(Wang, 2008b)。此外, Xu等(2014)研究表明熊本牡蠣(♀)和福建牡蠣(♂)存在單向雜交的可能, 而線粒體CO I基因遵循母系遺傳, 不能鑒定雜交種, 但核基因ITS2不僅能夠用于福建牡蠣和熊本牡蠣雜交種的鑒定, 也可以通過其片段長度多態性對福建牡蠣、熊本牡蠣、香港牡蠣和近江牡蠣()進行物種鑒定(Wang, 2008b; 蘇家齊, 2015; Yan, 2018)。相對而言, 針對大批量牡蠣樣品, 多重種特異性PCR是兼具準確性和經濟性的物種鑒定方法, 而核基因ITS2也是目前鑒定福建牡蠣和熊本牡蠣雜交種最有效的方法。
迄今關于福建沿海牡蠣種類及其分布的研究報道還不多。王海艷(2004)在形態學基礎上采用線粒體16S、CO I和核基因28S序列測序的方法對南方牡蠣物種進行了鑒定和聚類分析, 發現福州、莆田和廈門潮下帶淺水區為福建牡蠣, 廈門礁石上分布為熊本牡蠣。杜玄等(2009)通過多重種特異性PCR對福建沿海11個地點共327個牡蠣樣本鑒定, 發現福建沿海巨蠣屬有4種牡蠣分布, 分別是福建牡蠣、熊本牡蠣、香港牡蠣和近江牡蠣。總體上, 以往有關福建省牡蠣種類和分布的研究存在樣本量少或者采樣點僅涉及福建沿海部分地區(寧德、莆田或廈門) (Wang, 2010, 2013; Li, 2013; Li, 2020), 尚未見到全省沿海大量系統取樣進行牡蠣鑒定的研究報道。
近年來隨著福建沿海牡蠣養殖產業的迅速發展, 牡蠣苗種的異地遷移養殖愈來愈頻繁, 對各海區野生牡蠣資源造成沖擊, 而野生種質資源是保障產業特別是種業發展的重要物質基礎。目前尚未系統深入地開展福建沿海牡蠣種質資源評估、保護和利用, 為此, 有必要厘清福建沿海地區巨蠣屬牡蠣物種組成及其分布規律, 深化對當前巨蠣屬牡蠣種質資源認知。本研究在福建沿海系統采集野生苗種的牡蠣, 采用多重種特異性PCR方法進行物種鑒定, 并利用ITS2序列排查牡蠣雜交種。此外, 鑒于在利用多重種特異性PCR方法鑒定牡蠣發現部分福建牡蠣個體出現額外條帶, 在各采樣點隨機挑選30個牡蠣個體進行線粒體CO I序列分析以進一步確認多重種特異性PCR的物種鑒定結果。本研究為后續開展牡蠣群體遺傳分析提供了關鍵基礎資料, 為牡蠣種質資源的保護和利用打下重要基礎。
1 材料與方法
1.1 樣品的采集與固定
在福建沿海由北至南野生苗種棲息區共設置19個牡蠣采樣點, 采樣位點、采樣日期等詳細信息見圖1和表1。采樣時避開大量人工育苗牡蠣養殖區, 經調查選擇采集本海區自然附苗生長的牡蠣, 采集隨機且不分大小。從每個采樣點樣品中隨機選取50個牡蠣解剖取其閉殼肌, 用95%乙醇固定, 置于–20 °C條件下保存。
1.2 樣品形態特征鑒定
觀察并記錄牡蠣貝殼的各部分特征, 包括殼型、殼硬度、殼厚度、殼表面的顏色、放射肋分布、殼內面顏色、閉殼肌痕顏色等。通過形態特征對牡蠣進行初步鑒定。
1.3 DNA提取與檢測
取10~20 mg牡蠣閉殼肌, 使用北京天根生物技術有限公司的海洋動物DNA提取試劑盒提取基因組DNA, 使用超微量分光光度計進行DNA濃度和純度的檢測。DNA產物在–20 °C保存, 用于后續的PCR的擴增。
1.4 PCR擴增與物種鑒定
1.4.1 多重種特異性PCR擴增 利用Wang等(2008a)基于線粒體CO I基因開發的多重種特異性PCR鑒定方法可以快速鑒定巨蠣屬牡蠣。多重種特異性PCR共7條引物, 其中兩條外引物即CO I基因通用引物LCO1490與HCO2198 (Folmer, 1994), 牡蠣均能擴增出片段在697 bp左右的條帶; 5條內引物分別為巨蠣屬5種牡蠣的特異性引物(Wang, 2008a), 只有在其目標物種中才能擴增出相應的特異性條帶(表2)。使用的PCR反應總體系為25 μL, 兩種外引物分別均為0.5 μL, 5種內引物各為0.4 μL, DNA模板為1.5 μL, 2×Taq Master Mix (Vazyme)為12.5 μL, ddH2O補足至25 μL。循環參數設置: 預變性: 95 °C, 5 min; 30個循環(變性: 95 °C, 30 s, 退火: 48 °C, 1 min, 延伸: 70 °C, 1 min); 終延伸: 72 °C, 10 min。PCR產物經1.5%瓊脂糖凝膠電泳, 使用凝膠成像儀進行觀察PCR產物長度將牡蠣鑒定到種, 拍照并記錄分析。
1.4.2 ITS2擴增與雜交種鑒定 ITS2序列擴增所需要的引物序列為5′-TCTCGCCTGATCTGAGGTCG (5.8S forward)和5′-GCAGGACACATTGAACATCG (18S reverse) (Wang, 2008b)。PCR反應體系總體積為20 μL, 其中模板DNA 1 μL, 2×Taq PCR Master Mix (TIANGEN) 10 μL, 正反引物(F/R)各0.5 μL, ddH2O 8 μL。PCR擴增條件為: 95 °C預變性2 min, 之后進行30個循環(95 °C變性1 min, 51 °C復性1 min, 72 °C延伸1 min), 72 °C延伸10 min。PCR產物經2%瓊脂糖凝膠電泳檢測, 根據ITS2片段長度多態性排查是否存在雜交種。
表1 福建沿海牡蠣樣本采樣信息

Tab.1 Information of oyster sampling along Fujian coast
表2 5種巨蠣屬牡蠣的特異性引物與2種通用外引物序列

Tab.2 Sequences of specific primers and two universal primers of five Crassotrea oysters
1.4.3 CO I序列擴增測序 用CO I通用引物LCO1490和HCO2198進行PCR擴增, PCR反應體系總體積為20 μL, 其中模板DNA 1 μL, 2×Taq PCR Master Mix (TIANGEN) 10 μL, 正反引物(F/R)各0.5 μL, ddH2O為8 μL。反應條件為: 94 °C預變性3 min, 之后進行35個循環(94 °C變性45 s, 48 °C復性45 s, 72 °C延伸45 s), 72 °C延伸10 min。PCR產物經1%的瓊脂糖凝膠電泳檢測后送至上海生工生物工程有限公司進行測序。利用美國國家生物技術信息中心(National Center for Biotechnology In-formation, NCBI)中的BLAST在線分析比對鑒定牡蠣物種。
1.5 福建牡蠣非特異性條帶分析
將多重種特異性PCR鑒定中具有非特異性條福建牡蠣、正常條帶的福建牡蠣(ZJ群體)和熊本牡蠣(ZJ群體的牡蠣取樣于竹江汐露橋附近海區, 為在名稱上清晰區分兩種牡蠣, 故將ZJ群體熊本牡蠣簡稱為XLQ)共30個牡蠣的線粒體CO I基因測序序列, 采用UGENE42.0軟件(Okonechnikov, 2012)中Align with Clustalw進行多序列比對, 對線粒體CO I基因中熊本牡蠣種特異性引物設計區域的堿基序列進行分析, 并再次進行多重種特異性PCR和瓊脂糖凝膠電泳驗證。
2 結果
2.1 形態特征結果
福建沿海巨蠣屬牡蠣有3種, 分別為福建牡蠣、熊本牡蠣和香港牡蠣, 其主要形態學特征: 福建牡蠣殼為長型, 殼薄而脆, 左殼凹陷且附著面小, 右殼具有輕微突起, 殼表面呈黃色并帶有明顯褐色放射肋條紋分布, 殼內面呈白色, 閉殼肌痕褐色或無色; 熊本牡蠣為殼長型, 殼形較福建牡蠣厚且扁, 在右殼表面的邊緣有少量的褐色放射褶, 殼內面呈白色, 閉殼肌痕為褐色; 香港牡蠣殼為卵圓形或長型, 殼重而厚, 左殼凹陷呈杯狀, 右殼突起, 殼表面無明顯放射肋條紋, 右殼邊緣具有褐色薄且脆的鱗片, 殼內面呈瓷白色, 閉殼肌痕呈紫色或褐色(圖2)。
2.2 PCR擴增與物種鑒定結果
在多重種特異性PCR產物瓊脂糖凝膠電泳結果中, PCR產物擴增條帶大小697 bp是共有的外引物條帶, 其中擴增片段長度為222 bp是福建牡蠣特異性條帶; 擴增片段長度為546 bp是熊本牡蠣特異性條帶; 擴增片段大小為387 bp是香港牡蠣特異性條帶。而本研究在牡蠣鑒定過程中大部分(約75%)的福建牡蠣均出現三條帶, 多出一條與熊本牡蠣的特異性條帶(546 bp)大小相同條帶(圖3)。
ITS2序列擴增后經瓊脂糖凝膠電泳顯示, 福建牡蠣、熊本牡蠣、香港牡蠣的特征性片段分別為670、640、620 bp (圖4)。在本研究中960個牡蠣樣品均是一條清晰明亮的條帶, 未發現熊本牡蠣和福建牡蠣的單向雜交種。
牡蠣線粒體CO I基因測序BLAST比對鑒定結果與多重種特異性PCR結果和ITS2物種鑒定是一致的。
2.3 福建沿海巨蠣屬牡蠣種類及分布
對19個采樣點共960個牡蠣樣品的鑒定結果進行統計分析表明, 福建牡蠣(607個)占63.23%和熊本牡蠣(343個)占35.73% (表3), 香港牡蠣(10)占1.04%。其中福建牡蠣除九龍江角美(JM)采樣點外均有分布, 熊本牡蠣除莆田東沃村(DWC)采樣點外均有分布, 香港牡蠣僅在泉州濕地保護區洛陽橋(LYQ)采樣點具有自然分布。福建沿海19個采樣點牡蠣種類及數量統計分析表明, 福鼎市至羅源灣海區福建牡蠣和熊本牡蠣均為優勢種。從福清灣到廈門市10個群體中, 泉州市濕地保護區洛陽江(LYQ)熊本牡蠣為優勢種, 其他9個群體優勢種均為福建牡蠣。漳州市漳浦縣北江村(BJ)和沙西(SX)福建牡蠣為優勢種外, 其他群體優勢種均為熊本牡蠣(圖5)。
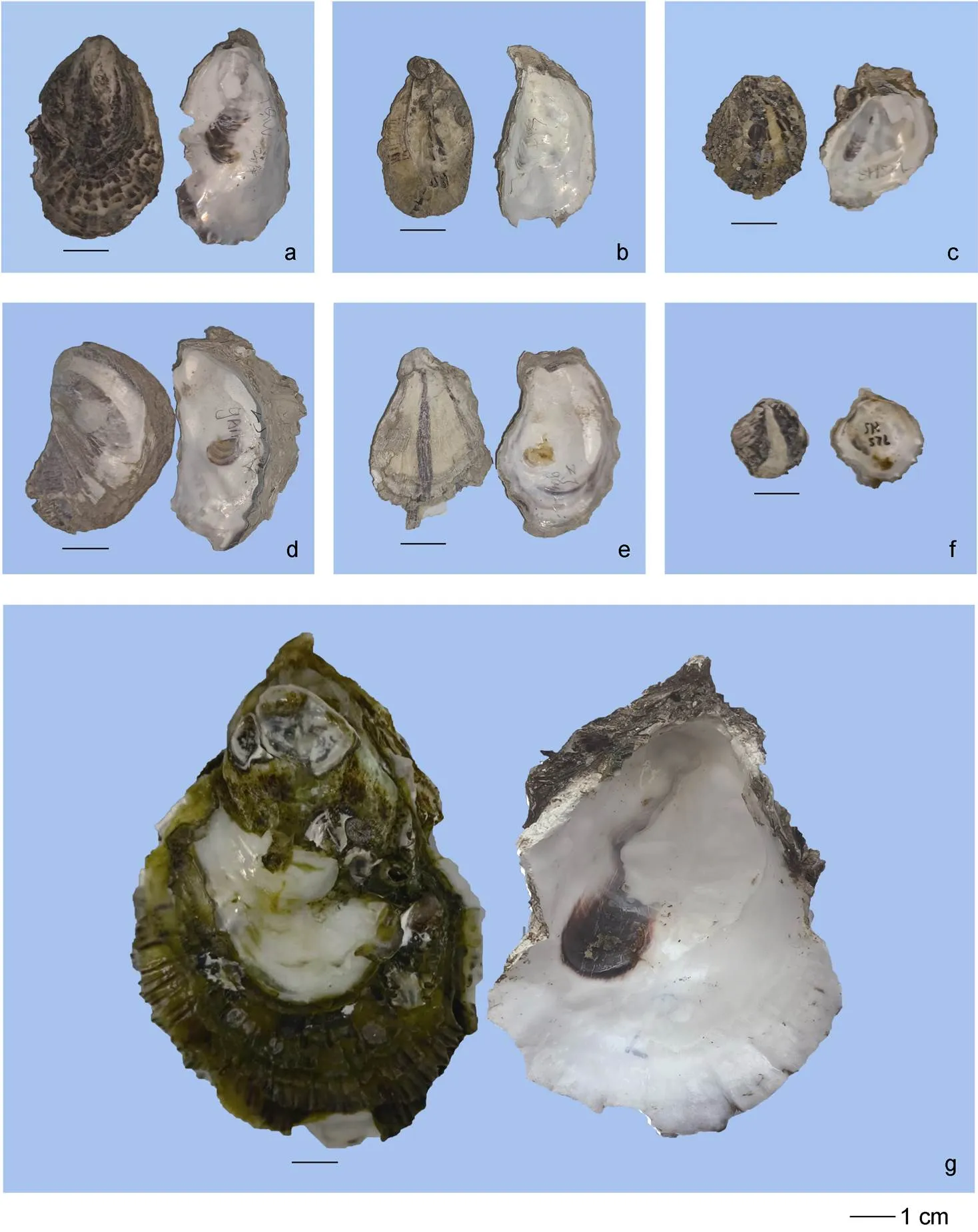
圖2 福建沿海3種巨蠣屬牡蠣形態特征
注: a~c. 福建牡蠣; d~f. 熊本牡蠣; g. 香港牡蠣

圖3 福建沿海巨蠣屬牡蠣多重種特異性PCR產物電泳圖
注: M. 2000 marker; BC. 空白對照; 1, 4~7. 福建牡蠣; 2, 8~11: 香港牡蠣; 3. 熊本牡蠣
2.4 福建牡蠣多重種特異性PCR出現非特異性條帶原因
5種巨蠣屬牡蠣線粒體CO I引物設計序列圖中, 在熊本牡蠣CO I基因特異性引物設計區(圖6a紅色虛線框), 福建牡蠣和熊本牡蠣CO I基因序列僅在第1位(A/G)和第7位堿基(T/C)兩處堿基不同。圖6b是福建寧德竹江群體(ZJ)牡蠣線粒體CO I基因多序列比對圖, 發現福建牡蠣線粒體CO Ⅰ片段在熊本牡蠣引物設計區段第7位堿基處具有T和C兩種堿基(圖6b紅色框)。在圖6b中隨機挑選福建牡蠣個體再次進行多重種特異性PCR擴增驗證, 福建牡蠣第7位堿基為C的牡蠣個體具有三條帶(ZJ18、ZJ19和ZJ31), 而第7位堿基為T的牡蠣個體僅有兩條帶(ZJ0、ZJ15) (圖6c)。

圖4 福建省泉州市LYQ部分樣品ITS2的電泳圖
注: M. 2000bp marker; 1~6. 香港牡蠣; 7~9, 12. 熊本牡蠣; 10~11. 福建牡蠣
表3 福建沿海19個采樣點巨蠣屬牡蠣種類及其數量
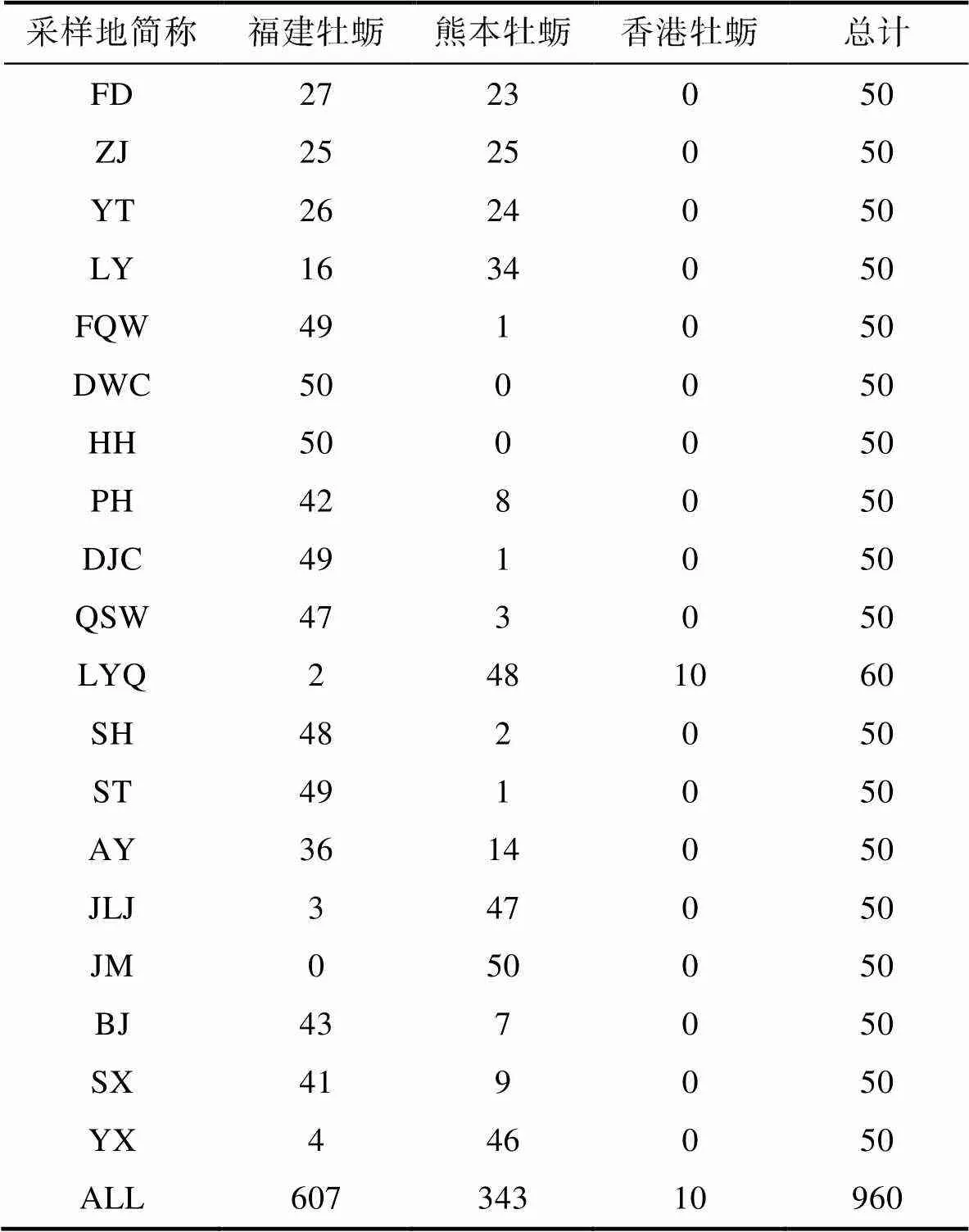
Tab.3 Species identification and the quantity of Crassostrea oysters in 19 sampling sites along Fujian coast

圖5 福建沿海各采樣點物種豐度圖
注: a. 五種巨蠣屬牡蠣線粒體CO I引物設計序列圖(Wang, 2008a); b. 福建沿海寧德竹江(ZJ)群體部分福建牡蠣和熊本牡蠣CO I序列比對圖; c. 福建沿海福建牡蠣寧德竹江群體(ZJ)多重種特異性PCR驗證圖
3 討論
3.1 福建沿海巨蠣屬牡蠣的種類及分布
本研究通過形態學特征、多重種特異性PCR、ITS2雜交種鑒定和CO I測序發現福建沿海巨蠣屬牡蠣有3種, 分別是福建牡蠣、熊本牡蠣和香港牡蠣, 其中福建牡蠣和熊本牡蠣為優勢種, 除個別采樣點外, 2個物種在各采樣海區均有分布。以往研究表明熊本牡蠣常與福建牡蠣在潮間帶呈同域分布且規模有限(Peng, 2021), 僅在江蘇南通市和浙江沿海發現大量分布的熊本牡蠣(Wang, 2013; Liu, 2021)。本研究在福建沿海泉州市濕地保護區洛陽橋海區(LYQ)、漳州市龍海區角美(JM)和漳州市云霄縣漳江長洋村海區(YX)采樣點發現了大規模自然分布的熊本牡蠣群體。此外, 在泉州濕地保護區洛陽橋海區(LYQ)發現了香港牡蠣, 而已有研究報道僅在福建沿海九龍江海區發現過香港牡蠣(Li, 2013, 2020), 本研究卻僅在九龍江海區發現人工養殖香港牡蠣。經我們實地調研發現LYQ的香港牡蠣是以“插石養殖”的方式, 苗種來自本海區的牡蠣群體, 排除人為“異地苗種”養殖因素, 因此泉州洛陽江海區(LYQ)的香港牡蠣很可能是本地種。
本次研究并未在福建沿海發現自然分布的近江牡蠣。杜玄等(2009)發現在漳州市九龍江海區海門島和白礁有近江牡蠣的自然分布, 但本研究在九龍江海區相近的2個采樣點及其他海區采樣點中均未發現近江牡蠣。近江牡蠣曾經在我國有較廣泛的分布(張璽等, 1956; Zhou, 2003), 但近年來近江牡蠣野生資源量急劇減少, 僅在北方有零星發現。據報道在我國南方近江牡蠣與香港牡蠣同域分布, 自然采苗的香港牡蠣苗種往往摻雜較多的近江牡蠣苗種, 而養殖戶排斥這種含高比例的近江牡蠣的香港牡蠣苗種, 造成對近江牡蠣自然苗種的負向選擇(Guo, 1999); 加上過度捕撈、徑流減少、環境污染和棲息地破壞等因素, 在過去的幾十年里, 野生資源急劇下降, 只在北方有少量局部分布(Li, 2020)。為搶救該種牡蠣資源, 在山東濰坊和江蘇省南通市已經建立了2個近江牡蠣種群保護區(Guo, 2009)。我們在福建沿海九龍江海區未發現自然分布的近江牡蠣, 推測一方面可能是由于近年來海區因棲息地的破壞、環境的污染、淡水徑流量減少等因素的影響造成九龍江口海區不適宜近江牡蠣的生存(Li, 2020, 2021; Qin, 2022); 另一方面可能是目前福建沿海近江牡蠣數量較少且呈零星分布, 需要更全面細致的采樣才能發現近江牡蠣。
3.2 影響福建沿海巨蠣屬牡蠣分布的生態與人為因素
本研究發現福建沿海巨牡蠣的分布符合其生態位分布, 其中福建牡蠣位于潮間帶及潮下帶淺水區, 熊本牡蠣位于潮間帶中上層, 香港牡蠣位于潮間帶的下層(Quan, 2012; Xu, 2014; Wang, 2020); 但其分布也同時受河流沖淡水和人為因素的影響(Wang, 2015; Moreira, 2016; Pinsky, 2020)。
內陸河流沖淡水是影響物種分布的因素之一。Dong等(2012)研究表明長江沖淡水在潮間帶生物之間形成隔離屏障, 提出長江沖淡水隔離假說。Ni等(2012)對蛤蜊的歷史冰川作用和長江流出假說的有效性研究表明, 沖淡水對物種的影響取決于生物特征(如鹽度和溫度耐受性)或棲息地特異性。牡蠣生活在環境變化劇烈河口潮間帶區, 而鹽度是直接影響牡蠣生存、生長和生理功能的環境因素之一(Yan, 2012)。福建沿海按平均年流量大小的內陸河流沖淡水主要有3條河流, 分別為閩江(1 707 m3/s)、九龍江(368 m3/s)和晉江(184 m3/s), 而內流沖淡水使其河口鹽度變化劇烈從而影響牡蠣的分布。研究表明在閩江、九龍江和晉江的河口存在較強的表層鹽度鋒, 其河口表層多呈明顯低鹽特性。鹽度鋒的位置受河流徑流量、潮汐和上升流等影響, 閩江和九龍江比晉江對周圍海域鹽度影響范圍廣(王偉強等, 2000; 邳青嶺等, 2008; 陳小紅等, 2009)。據以往研究可知不同牡蠣對鹽度的耐受性范圍不同, 福建牡蠣存活鹽度在10~35, 幼蟲適宜存活和生長鹽度在25~30 (巫旗生等, 2015; Moreira, 2016); 熊本牡蠣存活鹽度在7~35, 幼蟲適宜活和生長鹽度在20以下(Sekino, 2009; 呂曉燕, 2013); 香港牡蠣存活鹽度在5~25, 幼蟲適宜活和生長鹽度在15~30 (Huo, 2014; Xiao, 2018)。由此可以看出, 熊本牡蠣和香港牡蠣對低鹽有較強的適應能力, 適合棲息在與河流徑流相關的鹽度較低的水域(Hedgecock, 1999)。本研究的泉州濕地保護區洛陽橋海區(LYQ)、漳州九龍江口海區(JLJ)、漳州角美海區(JM)和漳州云霄縣漳江口(YX)等3個采樣點熊本牡蠣占比均在92%以上, 反映了這3個海區分別受晉江、九龍江和漳江沖淡水的影響, 其表層水域呈明顯低鹽特性, 有利于耐低鹽的熊本牡蠣棲息繁衍。
人為因素也是影響牡蠣分布的原因之一, 福建沿海牡蠣養殖歷史悠久, 早在宋朝時即在閩東北以“插竹養蠣”, 閩中以投石等方式養殖牡蠣, 因此福建沿海具備牡蠣自然海區采苗場這一特點。福建沿海牡蠣自然海區采苗場主要分布在閩中沿海地區, 主要有福州市福清灣、莆田市興化灣、惠安縣大港灣、石獅市深滬灣和南安縣圍頭灣(林丹等, 2019)。這些自然海區通常在每年立夏前后開始采苗(5~6月), 而福建牡蠣在自然海區繁殖季節為每年的4~9月, 5月上旬到6月下旬最盛(林丹等, 2019); 熊本牡蠣繁殖期在每年7~10月(鄧亞平, 2020)。福建沿海中部主要為福建牡蠣, 究其原因可能是受長期人工自然采苗影響。由于福建沿海中部地區具有自然海區采苗場的特點, 其采苗時間與福建牡蠣繁殖期較為一致, 在長期人工自然選擇情況下, 福建牡蠣在福建中部海區對環境和資源的競爭力比熊本牡蠣更具有優勢, 成為中部地區優勢種。此外, 臨近漳江灣的長洋村海區采樣點(YX)牡蠣優勢種為熊本牡蠣, 位于同一海區臨近臺灣海峽的沙西群體(SX)優勢種卻為福建牡蠣, 一方面可能因為沙西群體位于漳江下游且臨近外海, 其海區鹽度高于漳江上游長洋村采樣點, 適宜福建牡蠣生存; 另一方面可能與沙西當地漁民長期采集自然海區福建牡蠣苗種有關。
3.3 多重種特異性PCR鑒定方法的優化
本次研究發現大部分福建牡蠣多重種特異性PCR分析結果產生三條帶, 與Wang等(2008a)巨蠣屬牡蠣分析鑒定結果不同, 而多出的一條與熊本牡蠣種特異性條帶一致, 這與Liu等(2021)在對浙江沿海牡蠣分類鑒定中發現三條帶的現象一致, 對福建牡蠣鑒定可能會產生干擾。本研究通過福建牡蠣和熊本牡蠣CO I序列比對發現, 福建牡蠣CO I基因序列在熊本牡蠣特異性引物設計區中位于引物3′末端第7位堿基處存在T和C兩種堿基, 雖然Wang等(2008a)在設計熊本牡蠣特異性引物時, 為了提高其引物的特異性在引物3′末端的第2位堿基處加入C替代A的錯配(表2), 但由于該引物3′末端(C/T)和加入錯配(C/A)均是弱的錯配(Newton, 1989; Wangkumhang, 2007), 所以福建牡蠣在熊本牡蠣引物設計序列處3′末端第7位為C時, 在PCR擴增過程中, 由于DNA聚合酶受多種因素影響, 當引物錯配未達一定程度是可以擴增的, 其擴增的效率低于與引物完全互補的序列。這與本研究中福建牡蠣產生弱的非特異性條帶現象一致。因此, 福建牡蠣在熊本牡蠣設計引物處對應引物3′端第7位堿基的不同, 很可能是福建牡蠣額外產生與熊本牡蠣特異性條帶一致的原因。因此, 我們在使用多重種特異性PCR對牡蠣鑒定時, 可以適當調整其引物序列(在3′末端引入強的錯配, 如G/A)提高其引物的特異性或優化PCR反應參數抑制非特異性擴增, 以提高該方法鑒定牡蠣物種的準確性。
4 結論與展望
本研究發現福建沿海地區巨蠣屬牡蠣共3種, 分別是福建牡蠣、熊本牡蠣和香港牡蠣, 其中福建牡蠣與熊本牡蠣為優勢種。本研究在泉州濕地保護區洛陽江(LYQ)發現自然分布的香港牡蠣, 但在九龍江口并未發現自然分布的香港牡蠣。此外, 本研究在福建沿海未發現自然分布的近江牡蠣。牡蠣分布的影響不僅與其生態習性有關, 也受到河流沖淡水和海區人工采苗等因素影響。未來開展福建沿海野生牡蠣分布規律的研究, 牡蠣采樣點的系統設置是至關重要的一點, 必須考慮到潮位等垂直分布因素、沖淡水影響范圍以及海區人工采苗的歷史與現狀等。未來的研究應關注主要經濟物種在福建沿海分布的各野生群體的遺傳多樣性與遺傳分化, 評估高強度人工養殖是否對野生種質資源多樣性造成負面影響, 為開展福建省牡蠣野生種質資源的規劃和保護提供基礎資料。
王偉強, 張遠輝, 黃自強, 2000. 九龍江口-廈門港河口鹽度鋒面的特征[J]. 臺灣海峽, 19(1): 82-88.
王海艷, 2004. 中國近海常見牡蠣分子系統演化和分類的研究[D]. 青島: 中國科學院研究生院(海洋研究所): 94-95.
鄧亞平, 2020. 熊本牡蠣()微衛星標記開發, 野生及養殖群體遺傳多樣性分析[D]. 重慶: 重慶師范大學: 1-2.
呂曉燕, 2013. 熊本牡蠣人工繁育與長牡蠣單體苗種培育技術研究[D]. 青島: 中國海洋大學: 33-34.
農業農村部漁業漁政管理局, 全國水產技術推廣總站, 中國水產學會, 2022. 2022中國漁業統計年鑒[M]. 北京: 中國農業出版社: 27.
蘇家齊, 2015. 葡萄牙牡蠣與熊本牡蠣、僧帽牡蠣的種間雜交研究[D]. 青島: 中國海洋大學: 50-51.
杜玄, 郭希明, 錢魯閩, 2009. 福建沿海巨蠣屬牡蠣的主要種類及其分布[J]. 臺灣海峽, 28(3): 399-404.
巫旗生, 曾志南, 寧岳, 等, 2015. 葡萄牙牡蠣工廠化人工育苗技術[J]. 福建水產, 37(5): 399-405.
李孝緒, 齊鐘彥, 1994. 中國牡蠣的比較解剖學及系統分類和演化的研究[J]. 海洋科學集刊(35): 143-178.
李琪, 冉軻, 孔令鋒, 2021. DNA條形碼技術及其在海洋貝類種質資源保護中的應用[J]. 中國海洋大學學報, 51(5): 18-23.
邳青嶺, 胡建宇, 陳照章, 等, 2008. 2006年8月下旬福建沿海表層溫、鹽度分析[J]. 臺灣海峽, 27(4): 515-520.
張璽, 樓子康, 1956. 中國牡蠣的研究[J]. 動物學報, 5(1): 65-94.
陳小紅, 胡建宇, 邳青嶺, 等, 2009. 廈門—泉州近岸海域海表溫、鹽度密集走航觀測[J]. 地球科學進展, 24(6): 629-635.
林丹, 孫敏秋, 張克烽, 等, 2019. 福建牡蠣產業發展形勢分析[J]. 中國水產(3): 53-57.
闕華勇, 劉曉, 王海艷, 等, 2003. 中國近海牡蠣系統分類研究的現狀和對策[J]. 動物學雜志, 38(4): 110-113.
CORDES J F, XIAO J, REECE K S, 2008. Discrimination of nineoyster species based upon restriction fragment- length polymorphism analysis of nuclear and mitochondrial DNA markers [J]. Journal of Shellfish Research, 27(5): 1155-1161.
DONG Y W, WANG H S, HAN G D,, 2012. The impact of Yangtze River discharge, ocean currents and historical events on the biogeographic pattern ofalong the China coast [J]. PLoS One, 7(4): e36178.
FOLMER O, BLACK M, HOEH W,, 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates [J]. Molecular Marine Biology and Biotechnology, 3(5): 294-299.
GUO X M, 2009. Use and exchange of genetic resources in molluscan aquaculture [J]. Reviews in Aquaculture, 1(3/4): 251-259.
GUO X M, FORD S E, ZHANG F S, 1999. Molluscan aquaculture in China [J]. Journal of Shellfish Research, 18(1): 19-31.
HEDGECOCK D, LI G, BANKS M A,, 1999. Occurrence of the Kumamoto oysterin the Ariake Sea, Japan [J]. Marine Biology, 133(1): 65-68.
HUO Z M, WANG Z P, LIANG J,, 2014. Effects of salinity on embryonic development, survival, and growth of[J]. Journal of Ocean University of China, 13(4): 666-670.
LI A, DAI H, GUO X M,, 2021. Genome of the estuarine oyster provides insights into climate impact and adaptive plasticity [J]. Communications Biology, 4(1): 1287.
LI A, WANG C G, WANG W,, 2020. Molecular and fitness data reveal local adaptation of southern and northern estuarine oysters () [J]. Frontiers in Marine Science, 7: 589099.
LI S, LI Q, YU H,, 2015. Genetic variation and population structure of the Pacific oysterin the northwestern Pacific inferred from mitochondrial COI sequences [J]. Fisheries Science, 81(6): 1071-1082.
LI L, WU X Y, YU Z N, 2013. Genetic diversity and substantial population differentiation inrevealed by mitochondrial DNA [J]. Marine Genomics, 11: 31-37.
LIU J, LI Q, KONG L F,, 2011. Identifying the true oysters (Bivalvia: Ostreidae) with mitochondrial phylogeny and distance‐based DNA barcoding [J]. Molecular Ecology Resources, 11(5): 820-830.
LIU S, XUE Q G, XU H Q,, 2021. Identification of main oyster species and comparison of their genetic diversity in Zhejiang coast, south of Yangtze river estuary [J]. Frontiers in Marine Science, 8: 662515.
MOREIRA A, FIGUEIRA E, SOARES A M VM,, 2016. Salinity influences the biochemical response ofto Arsenic [J]. Environmental Pollution, 214: 756-766.
NEWTON C R, GRAHAM A, HEPTINSTALL L F,, 1989. Analysis of any point mutation in DNA. The amplification refractory mutation system (ARMS) [J]. Nucleic Acids Research, 17(7): 2503-2516.
NI G, LI Q, KONG L F,, 2012. Phylogeography of bivalve: testing the historical glaciations and Changjiang River outflow hypotheses in northwestern Pacific [J]. PLoS One, 7(11): e49487.
OKONECHNIKOV K, GOLOSOVA O, FURSOV M,2012. Unipro UGENE: a unified bioinformatics toolkit [J]. Bioinformatics, 28(8): 1166-1167.
PENG D M, ZHANG S C, ZHANG H Z,, 2021. The oyster fishery in China: Trend, concerns and solutions [J]. Marine Policy, 129: 104524.
PINSKY M L, SELDEN R L, KITCHEL Z J, 2020. Climate-driven shifts in marine species ranges: Scaling from organisms to communities [J]. Annual Review of Marine Science, 12: 153-179.
QIN Y P, LIAO Q L, SHI G P Y,, 2022. Comparison of growth, survival and fertility of the southern and northern populations ofand their hybrids in southern China [J]. Aquaculture, 549: 737-744.
QUAN W M, HUMPHRIES A T, SHEN X Q,, 2012. Oyster and associated benthic macrofaunal development on a created intertidal oyster () reef in the Yangtze River estuary, China [J]. Journal of Shellfish Research, 31(3): 599-610.
SEKINO M, 2009. In search of the Kumamoto oyster(Amemiya, 1928) based on molecular markers: is the natural resource at stake? [J]. Fisheries Science, 75(4): 819-831.
WANG T N, FAN R L, CHENG Q Q,, 2020. Intertidal zonation of the suminoe oysterand the Kumamoto Oysteron the coast of the Northern East China Sea [J]. Journal of Shellfish Research, 39(1): 31-41.
WANG H Y, GUO X M, 2008a. Identification ofand related oysters by multiplex species-specific PCR [J]. Journal of Shellfish Research, 27(3): 481-487.
WANG Y P, GUO X M, 2008b. ITS length polymorphism in oysters and its use in species identification [J]. Journal of Shellfish Research, 27(3): 489-493.
WANG H Y, GUO X M, ZHANG G F,, 2004. Classification of jinjiang oysters(Gould, 1861) from China, based on morphology and phylogenetic analysis [J]. Aquaculture, 242(1/2/3/4): 137-155.
WANG H Y, QIAN L M, LIU X,, 2010. Classification of a common cupped oyster from southern China [J]. Journal of Shellfish Research, 29(4): 857-866.
WANG H Y, QIAN L M, WANG A M,, 2013. Occurrence and distribution of(Amemiya 1928) in China [J]. Journal of Shellfish Research, 32(2): 439-446.
WANG J, TSANG L M, DONG Y W, 2015. Causations of phylogeographic barrier of some rocky shore species along the Chinese coastline [J]. BMC Evolutionary Biology, 15: 114.
WANG J F, XU F, LI L,, 2014. A new identification method for five species of oysters in genusfrom China based on high-resolution melting analysis [J]. Chinese Journal of Oceanology and Limnology, 32(2): 419-425.
WANGKUMHANG P, CHAICHOOMPU K, NGAMPHIW C,, 2007. WASP: a Web-based Allele-Specific PCR assay designing tool for detecting SNPs and mutations [J]. BMC Genomics, 8: 275.
XIA J, YU Z, KONG X J, 2009. Identification of sevenoysters from the South China Sea using PCR-RFLP analysis [J]. Journal of Molluscan Studies, 75(2): 139-146.
XIAO S, WONG N K, LI J,, 2018. Analysis oftranscriptomes reveals divergent adaptive response to hyper-and hypo-salinity in the Hong Kong oyster,[J]. Frontiers in Physiology, 9: 1491.
XU F, LI L, WANG J F,, 2014. Use of high-resolution melting analysis for detecting hybrids between the oystersandreveals bidirectional gametic compatibility [J]. Journal of Molluscan Studies, 80(4): 435-443.
YAN L L, SU J Q, WANG Z P,, 2018. Growth performance and biochemical composition of the oysters,and their hybrids in southern China[J]. Aquaculture Research, 49(2): 1020-1028.
YAN X L, ZHAI W D, HONG H S,, 2012. Distribution, fluxes and decadal changes of nutrients in the Jiulong River Estuary, Southwest Taiwan Strait [J]. Chinese Science Bulletin, 57(18): 2307-2318.
ZHOU M F, ALLEN JR S K, 2003. A review of published work on[J]. Journal of Shellfish Research, 22(1): 1-20.
THE SPECIES AND DISTRIBUTION OFOYSTER IN FUJIAN COAST
YU Shi-Qi1, HAN Zi-Qiang1, CHEN Yan-Ting2, GUO Tuan-Yu3, QUE Hua-Yong1
(1. Fisheries College, Jimei University, Key Laboratory of Mariculture for the East China Sea, Ministry of Agriculture and Rural Affairs, Fujian Engineering Research Center of Aquatic Breeding and Healthy Aquaculture, Xiamen 361021, China; 2. Fujian Fisheries Technology Extension Center, Fuzhou 350002, China; 3. College of Marine Biology, Xiamen Ocean Vocational College, Xiamen 361100, China)
Fujian Province is located on the southeast of China facing the East China Sea. The irregular coastline forms numerous bays with several sea-going rivers, which is suitable for living and reproduction of oysters. It is one of the regions with the most abundant natural oysters in China. At present, there are few reports on the composition and distribution of oyster species along the coast of Fujian Province. Given that the main bays of Fujian Province have experienced high-intensity oyster culture in the past 20 years, it is necessary to understand the natural species diversity along the coast of Fujian Province. Therefore, based on field surveys and sampling, we used multiplex species-specific PCR, identification of ITS2 nuclear gene hybrids, and CO I sequencing to identifyoyster species from the coastal region of Fujian Province and to determine the distribution of specific species. Results show that three species of, namely,, and, were found in a total of 960 samples collected from 19 sites of typical natural oyster seed-production areas along the coast of Fujian Province. Nowas found as previously reported. Among them,(607) accounted for 63.23%,(343) for 35.73%, and(10) for 1.04%.andwere the dominant species and distributed in the majority sampling areas. However,was merely found in the Quanzhou Bay Estuary Wetland Nature Reserve, which was newly discovered natural population. The species and distribution of oysters in the Fujian coastal area revealed in this study provide key basic data for the analysis of genetic diversity and the genetic differentiation of oyster populations, which pave the road for protection and utilization of oyster germplasm resources.
oyster species;; Fujian coast; multiplex species-specific PCR; hybrid identification based on ITS2 sequence; CO I sequence analysis
* 國家自然科學基金項目, 32172979號; 福建省自然科學基金項目, 2021J05159號。于詩奇, 碩士研究生, E-mail: 942571813@qq.com
闕華勇, 教授, 博士生導師, E-mail: hque@jmu.edu.cn
2023-04-19,
2023-07-23
Q789; S931; S968.3
10.11693/hyhz20230400088
