育肥飼料中添加β-胡蘿卜素對中華絨螯蟹雌體卵巢發育、抗氧化免疫性能和組織色澤的影響*
2023-12-06 07:38:38姜曉東朱凌翔成永旭吳旭干
海洋與湖沼 2023年6期
姜曉東 朱凌翔 成永旭 吳旭干
育肥飼料中添加β-胡蘿卜素對中華絨螯蟹雌體卵巢發育、抗氧化免疫性能和組織色澤的影響*
姜曉東1, 2, 3朱凌翔1, 2, 3成永旭1, 2, 3吳旭干1, 2, 3①
(1. 上海海洋大學 農業農村部淡水種質資源重點實驗室 上海 201306; 2. 上海海洋大學 水產動物遺傳育種上海市協同創新中心 上海 201306; 3. 上海海洋大學 農業農村部魚類營養和環境生態研究中心 上海 201306)
為研究飼料中添加β-胡蘿卜素對中華絨螯蟹成體雌蟹育肥效果的影響, 分別在飼料中添加不同濃度的人工合成β-胡蘿卜素, 配制五組實驗飼料(β-胡蘿卜素測定含量分別為3.3、56.7、108.9、207.9和320.0 mg/kg)投喂生殖蛻殼后雌蟹, 評價了飼料中添加β-胡蘿卜素對增重率(WGR)、性腺指數(GSI)、肝胰腺指數(HSI)、常規生化組成、抗氧化和免疫指標、色澤參數和類胡蘿卜素含量的影響。結果顯示: (1) 飼料中添加人工合成β-胡蘿卜素對雌蟹育肥期間的WGR、GSI和HSI均無顯著影響, 但飼料中添加β-胡蘿卜素顯著增加了中華絨螯蟹肝胰腺中總脂含量; (2) 就抗氧化和免疫指標而言, 肝胰腺和血淋巴中SOD、T-AOC、GSH-Px、AKP和ACP免疫相關酶活均隨著飼料中β-胡蘿卜素添加量的增加呈“先增加后降低”的變化趨勢, 飼料中β-胡蘿卜素含量為100~200 mg/kg時, 組織中抗氧化和免疫相關酶活最高; (3) 卵巢、肝胰腺和蟹殼中蝦青素和β-胡蘿卜素含量均隨著飼料中β-胡蘿卜素添加量的增加而顯著增加, 但飼料中添加β-胡蘿卜素僅顯著增加了卵巢和肝胰腺紅度值(*)。綜上, 飼料中添加人工合成β-胡蘿卜素對中華絨螯蟹雌體育肥期間的增重率和性腺發育無明顯影響, 但添加適量β-胡蘿卜素可以提高中華絨螯蟹抗氧化免疫性能、組織色澤及其類胡蘿卜素含量, 綜合考慮各項指標及飼料成本, 建議中華絨螯蟹雌體育肥飼料中人工合成β-胡蘿卜素的適宜含量為100 mg/kg左右。研究結果不僅有助于了解β-胡蘿卜素對甲殼動物生理功能的影響, 而且有助于中華絨螯蟹功能性飼料的開發。
中華絨螯蟹; 育肥飼料; 類胡蘿卜素; 養殖性能; 色澤參數
中華絨螯蟹(, 以下簡稱河蟹)是我國重要的經濟水產養殖品種, 因富含礦物質元素、脂肪酸、氨基酸、核苷酸和膽固醇等多類營養物質而受到廣大消費者的青睞, 具有較高的經濟價值和食用價值(Shao, 2013)。自20世紀90年代末河蟹土池生態育苗技術得到突破, 河蟹產業規模迅速擴大, 經二十多年發展已成為我國水產養殖行業發展最迅速、最具特色的水產品產業之一。盡管我國河蟹養殖產量巨大, 但池塘養殖河蟹普遍存在煮熟后殼色、卵巢和肝胰腺顏色較淡等問題, 而野生和湖泊增殖河蟹通常殼色和卵巢顏色較紅(Kong, 2012)。色澤是影響消費者對甲殼動物感官評價的重要因素之一, 色澤較淡直接影響到商品河蟹的市場價值(王世會等, 2021)。研究表明甲殼動物的體色與其體內的類胡蘿卜素(尤其是蝦青素)組成和含量密切相關, 因此在飼料中添加類胡蘿卜素是改善池塘養殖河蟹色澤的有效方法(Jiang, 2019)。此外, 河蟹在室外池塘養殖過程中易受到氨氮、亞硝酸鹽、pH值和缺氧等環境脅迫因子的影響, 并且夏季水溫較高時危害尤為嚴重(Qiu, 2011)。這些不利的環境脅迫因子可能會對河蟹的免疫系統、生理過程、存活及生長產生負面影響(張世勇等, 2013)。研究表明, 在飼料中添加類胡蘿卜素來增強河蟹抗氧化性能及抗逆性能, 是減輕河蟹受環境脅迫影響的重要手段(Wei, 2016)。
β-胡蘿卜素是具有重要生理功能的天然色素, 同時也是合成維生素A的前體, 在食品和飼料添加劑等方面得到廣泛應用(Gul, 2015)。除了典型的著色作用外, 研究表明β-胡蘿卜素對甲殼類動物的抗氧化性能、免疫性能、抗逆性能及抗病力均具有一定的提升效果, 突出了β-胡蘿卜素對于甲殼動物健康養殖的重要性(Madhumathi, 2011; Medina Félix, 2017)。眾所周知, 甲殼動物無法從頭合成β-胡蘿卜素等類胡蘿卜素, 僅可以通過儲存或轉化餌料中外源類胡蘿卜素后將其沉積于體內, 因此甲殼動物體內類胡蘿卜素沉積很大程度上依賴于飼料中補充的類胡蘿卜素(Boonyaratpalin, 2001)。此外, 據報道甲殼動物可以將攝入的β-胡蘿卜素轉化為抗氧化活性更強且價值更高的蝦青素(Martín, 2008)。這便增加了一種可能性, 即如果β-胡蘿卜素在甲殼動物體內轉化為蝦青素的效率足夠高, 那在飼料中添加β-胡蘿卜素將可以提供一種可替代且成本更低的方法來實現甲殼動物的所需特性。
目前, 類胡蘿卜素在河蟹飼料中的研究進展主要集中于飼料中添加蝦青素對河蟹色澤、抗氧化性能及抗逆性能影響的研究, 但針對飼料中添加β-胡蘿卜素對河蟹影響的研究報道很少(Long, 2017; Jiang, 2020)。先前在哺乳動物上的研究表明, 當飼料中β-胡蘿卜素含量達到特定物種生長和生理狀態所需的最佳水平后, 過量添加β-胡蘿卜素將對動物產生促氧化作用(Siems, 2005)。本實驗主要探究飼料中添加不同水平β-胡蘿卜素對河蟹性腺發育、抗氧化性能、非特異性免疫和色澤的影響, 并根據這些結果探索河蟹飼料中β-胡蘿卜素的最佳添加量。該研究結果不僅有助于了解β-胡蘿卜素對甲殼類生理功能的影響, 而且有助于河蟹功能性飼料的開發。
1 材料與方法
1.1 飼料配方及制備
通過在飼料原料中添加不同水平人工合成β-胡蘿卜素(11.75%, 廣州立達爾生物科技股份有限公司)配制5組實驗飼料, 使得飼料中β-胡蘿卜素含量分別為50、100、200和300 mg/kg (記為Diet 2, Diet 3, Diet 4和Diet 5), 并以不添加β-胡蘿卜素的飼料組作為對照組(記為Diet 1)。所有飼料原料粉碎之后過80目篩, 按照飼料配方充分混勻, 加入25%蒸餾水混合后, 用制粒機制備粒徑3.0 mm、長度10 mm左右的飼料, 然后在室溫下風干。為了防止飼料中β-胡蘿卜素氧化, 所有風干后的實驗飼料均用黑色塑料袋于–20 °C冰箱中保存備用。表1為實驗飼料配方、飼料常規生化組成及主要類胡蘿卜素的實際含量。
表1 實驗飼料配方、生化組成及類胡蘿卜素含量

Tab.1 The formulation, proximate composition, and actual carotenoid concentration of experimental diets

續表
1.2 實驗設計與養殖管理
2019年8月下旬在上海海洋大學崇明基地的15個PE桶(直徑×高度= 108 cm×120 cm)中進行成蟹養殖實驗。從該基地的土池中隨機選擇300只成熟河蟹雌體(體重70~90 g)并暫養至PE桶中。室內暫養3~5 d后, 進一步選擇四肢健全和活力較好的個體用于接下來的養殖實驗。每組飼料組設定三個重復圓桶, 每個圓桶放16只成熟個體。為避免河蟹相互打斗影響成活率, 圓桶底部放置塑料假草。實驗用水為消毒沉淀后的河道水, 養殖實驗過程中圓桶水位保持在50 cm左右, 圓桶中水體的循環速度為3 L/min。所有圓桶均每天24 h連續曝氣, 以保持養殖水體溶解氧(DO)大于5 mg/mL。實驗期間光照周期為12 h︰12 h。河蟹養殖期間按體重的3%投喂實驗飼料, 每天虹吸清除糞便和殘餌。實驗過程中每天測定水質, 并將水質維持在最佳范圍內(pH 7~8.5, 氨氮<0.4 mg/L, 亞硝酸鹽<0.05 mg/L)。
1.3 生長數據收集及樣本采集
分別于養殖實驗35 d及70 d后從每個圓桶中隨機選擇3只河蟹并對其稱重。從每只河蟹第三條附肢的根部抽取2 mL左右血淋巴放至2 mL離心管中, 隨后–80 °C冷凍保存。解剖河蟹獲得卵巢、肝胰腺和肌肉并稱重, 隨后將每只河蟹頭胸甲、卵巢、肝胰臟和肌肉分別于–40 °C保存, 用于后續生化分析。使用以下公式計算河蟹增重率(WGR)、性腺指數(GSI)和肝胰腺指數(HSI):
WGR(%)=(最終體重–初始體重)/初始體重×100, (1)
GSI(%)=性腺重/體重×100, (2)
HSI(%)=肝胰腺重/體重×100. (3)
1.4 抗氧化及非特異性相關酶活測定
使用IKA微型勻漿器(德國IKA公司)將解凍后的血淋巴樣品勻漿, 隨后將勻漿液在10 000 r/min和4 °C條件下離心20 min, 收集離心后上清液并保存于–40 °C冰箱以備后續使用。稱重來自每只河蟹的約0.1 g肝胰腺樣品, 并按1︰5 (/)的比例添加到預冷的生理鹽水中, 然后在2 mL離心管中使用IKA勻漿器進行勻漿。然后將勻漿液在4 °C和10 000 r/min條件下離心10 min, 取上清液于–40 °C保存供以后分析。使用分光光度計和相應的實驗試劑盒(南京建成生物工程研究所)測定河蟹肝胰臟和血淋巴中的超氧化物歧化酶(SOD)、總抗氧化劑能力(T-AOC)、谷胱甘肽過氧化物酶(GSH-Px)、丙二醛(MDA)、堿性磷酸酶(AKP)、酸性磷酸酶(ACP)、谷氨酸草酰乙酸轉氨酶(GOT)和谷氨酸丙酮酸轉氨酶(GPT)水平。
1.5 常規生化、色澤及類胡蘿卜素組成測定
根據AOAC的方法對實驗飼料的水分、粗蛋白和灰分含量進行測定(AOAC, 2000)。根據Folch等(1957)的方法用氯仿-甲醇(2︰1,/)提取總脂。在提取組織中總類胡蘿卜素之前, 將每只蟹的頭胸甲、卵巢和肝胰臟分別冷凍干燥并研磨, 隨后將每個平行中的同一組織合并混合, 作為一個重復。用丙酮提取所有樣品中的總類胡蘿卜素, 并使用紫外可見分光光度計(型號: T6新世紀, 北京普析通用儀器有限責任公司)在470 nm波長下測定其含量(Johnston, 2000)。首先通過酶水解將酯化的類胡蘿卜素水解(Wade, 2005), 然后使用Agilent 1260高效液相色譜(HPLC)系統(Agilent Technologies Inc., 美國加利福尼亞州), 按照Peng等(2008) 的方法分析組織中蝦青素和β-胡蘿卜素含量。Agilent 1260 HPLC系統配備了YMC?類胡蘿卜素C30色譜柱(4.6×150 mm, 填料直徑=3 μm, YMC Co., Ltd., 日本京都), 流動相則由A和B組成, 其中流動相A由甲醇︰甲基叔丁基醚︰甲酸(3︰2︰0.01,//)組成, 流動相B為醇︰三乙胺(100︰0.04,/)的混合物。根據蝦青素和β-胡蘿卜素市售標準品鑒定并定量這兩種類胡蘿卜素。蝦青素(編號A9335)購自Sigma-Aldrich (美國), β-胡蘿卜素(編號CA10947000)標準品購自Dr. Ehrenstorfer GmbH (德國奧格斯堡)。
使用色差儀(CR400, 日本柯尼卡美能達公司)測定冷凍干燥后頭胸甲、卵巢和肝胰腺的色度值, 色澤參數包括*、*和*值, 分別代表亮度、紅色和黃度(León, 2006)。每個實驗組測定9只成蟹(3只成蟹/重復)顏色參數, 每個頭胸甲、卵巢和肝胰腺樣品選擇三個相對光滑的點進行*、*和*值測定, 結果以每組樣品平均值表示。
1.6 RNA提取和qRT-PCR
參照試劑盒操作步驟, 使用RNAiso Plus (TaKaRa)從每個肝胰腺中提取總RNA。分別使用瓊脂糖凝膠電泳和紫外分光光度計(Q5000, Quawell Technology Inc, 美國)對總RNA進行定量測定。僅使用高質量的總RNA樣品(OD260/280范圍為1.8~2.2, RIN≥8.0)進行cDNA反轉錄。根據試劑盒操作說明, 使用PrimeScriptTMRT-PCR試劑盒(TaKaRa)將總RNA (2 μg)逆轉錄為總體積40 μL。使用SYBR RT-PCR試劑盒(RR420A, TaKaRa)在ABI實時PCR系統(ABI-7500; ThermoFisher, 新加坡)上進行所有實時反應。總反應體積為10 μL, 其中包含5 μL 2x SYBR Master Mix、1 μL稀釋的cDNA、0.2 μL正向和反向引物和ROX Reference DyeII, 以及3.4 μL Rnase-free-ddH2O。擴增條件為: 95 °C持續30 s、95 °C進行5 s的40個循環, 60 °C進行34 s, 95 °C進行15 s, 60 °C進行60 s, 95 °C進行15 s。針對β-肌動蛋白內部參考基因的表達進行標準化, 并根據2–ΔΔCt方法分析數據。表2中描述了CAT (過氧化氫酶)、Trx (硫氧還蛋白)、Prx6 (過氧還蛋白)、GST (谷胱甘肽轉移酶)和β-Actin的引物序列。
表2 熒光定量PCR分析的引物及相關信息

Tab.2 Primers and relative information used for quantitative real-time PCR analysis
1.7 統計分析
使用SPSS 19.0軟件進行統計分析。數據表示為平均值±標準誤(SE), 用 Levene 法進行方差齊性檢驗, 當數據不滿足齊性方差時對百分比數據進行反正弦或者平方根處理, 采用單因子ANOVA對實驗結果進行方差分析, 采用Tukeys-b(K)法進行多重比較; 當數據轉換后仍不滿足齊性方差時, 采用Games- Howell非參數檢驗進行多重比較。取<0.05為差異顯著, 在GraphPad Prism 9軟件上繪制相關圖表。
2 結果
2.1 性腺發育及生化組成
飼料中添加β-胡蘿卜素對成體雌蟹育肥期間增重率(WGR)、性腺指數(GSI)、肝胰腺指數(HSI)和最終成活率的影響如表3所示。經35 d育肥之后, 各組雌蟹WGR、GSI和HSI差異較小(>0.05), 分別為5.5%、7%和8%左右。經70 d育肥之后, 各組雌蟹WGR和GSI在育肥35 d的基礎之上分別提高0.5%和2%左右, 而HSI則降低2.5%左右。就成活率而言, 各組成活率均在75%~85%范圍內, 其中Diet 1組成活率最低, 為77.08%, Diet 4組成活率最高, 為83.33%。
表3 育肥飼料中添加β-胡蘿卜素對成體雌蟹性腺發育的影響
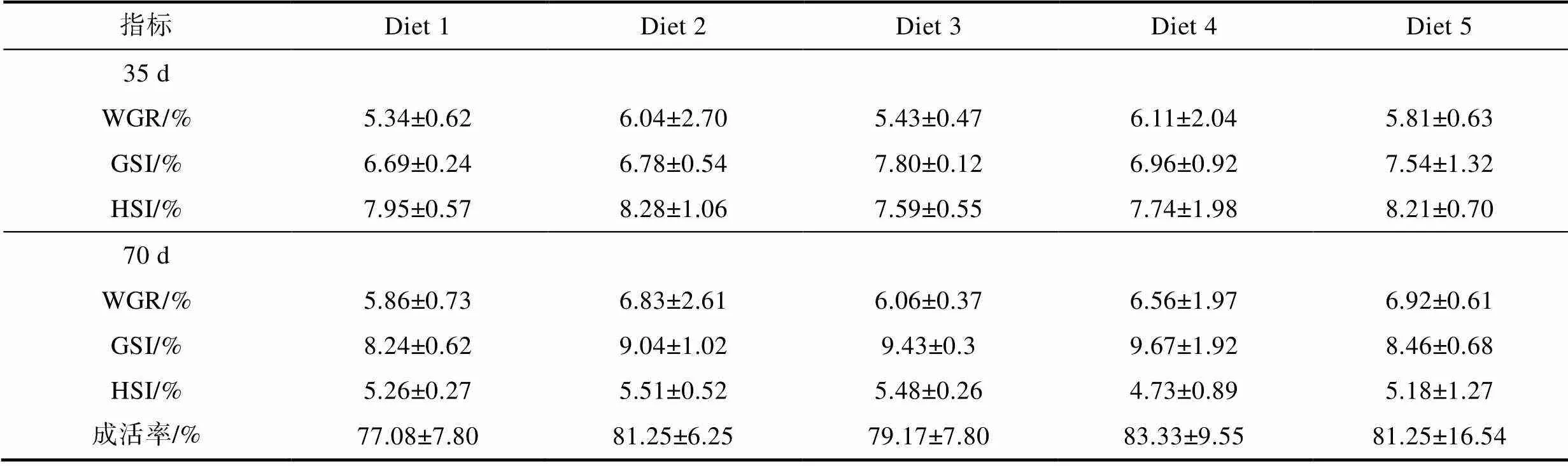
Tab.3 Effects of dietary β-carotene on the gonad development of adult female E. sinensis
育肥飼料中添加β-胡蘿卜素對成體雌蟹卵巢、肝胰腺及肌肉生化組成的影響如表4所示。結果表明, 飼料中添加不同水平β-胡蘿卜素對成體雌蟹卵巢和肌肉中的常規生化影響較小。肝胰腺中對照組水分含量顯著高于β-胡蘿卜素添加組, 而粗脂肪含量顯著低于各β-胡蘿卜素添加組(<0.05), 說明飼料中添加β-胡蘿卜素可以顯著增加成體雌蟹肝胰腺中粗脂肪含量。
表4 育肥飼料中添加β-胡蘿卜素對雌蟹卵巢、肝胰腺及肌肉生化組成的影響(%濕重)

Tab.4 Effects of dietary β-carotene on the proximate composition in ovary, hepatopancreas, and muscle of adult female E. sinensis (% wet weight)
注: 上標不同小寫字母表示差異顯著(<0.05)。下同
2.2 免疫及抗氧化性能
育肥飼料中添加β-胡蘿卜素對成體雌蟹抗氧化相關酶活的影響如表5所示。肝胰腺和血淋巴中SOD、T-AOC和GSH-Px活性均隨著飼料中β-胡蘿卜素添加量的增加呈“先增加后降低”的變化趨勢, 且β-胡蘿卜素添加量為100~200 mg/kg時這些抗氧化相關酶活的數值顯著較高。與之相反, 肝胰腺和血淋巴中MDA含量均隨著飼料中β-胡蘿卜素添加量的增加而呈“先降低后增加”的變化趨勢, 且當β-胡蘿卜素添加量為100 mg/kg時含量最低。
飼料中添加β-胡蘿卜素對成蟹雌體免疫相關酶活的影響如表6所示。肝胰腺和血淋巴中AKP及ACP活性均隨著飼料中β-胡蘿卜素添加量的增加而呈“先增加后降低”的變化趨勢, 且當β-胡蘿卜素添加量為100~200 mg/kg時這些免疫相關酶活最高。與之相反, 肝胰腺和血淋巴中GPT及GOT活性均隨著飼料中β-胡蘿卜素添加量的增加而呈“先降低后增加”的變化趨勢, 且當β-胡蘿卜素添加量為100 mg/kg時酶活最低。
表5 育肥飼料中添加β-胡蘿卜素對抗氧化相關酶活的影響

Tab.5 Effects of dietary β-carotene meal on the antioxidant indices of adult female E. sinensis
表6 育肥飼料中添加β-胡蘿卜素對血淋巴中免疫相關酶活的影響

Tab.6 Effects of dietary β-carotene on the immune indices of adult female E. sinensis
飼料中添加β-胡蘿卜素對成體雌蟹肝胰腺中抗氧化相關基因表達量的影響如圖1所示。CAT、Trx、Prx6及GST等基因的表達量均隨著飼料中β-胡蘿卜素添加量的增加而呈“先增加后降低”的變化趨勢。其中, 當飼料中β-胡蘿卜素添加量為100 mg/kg時,CAT、Trx、Prx6和GST表達量均為最高。基于雙折線回歸分析的結果表明, 當飼料中β-胡蘿卜素添加量為113.64、114.20、98.17及98.97 mg/kg時, 肝胰腺中CAT、Trx、Prx6及GST等基因的表達量分別最高。
2.3 色澤及類胡蘿卜素組成
飼料中添加β-胡蘿卜素對各組織色澤的影響如圖2所示。蟹殼色度值中*、*和*均隨著飼料中β-胡蘿卜素添加量的增加而呈升高趨勢, 但不同β-胡蘿卜素添加水平之間無顯著差異; 肝胰腺色度值中*值隨著飼料中β-胡蘿卜素添加量的增加而顯著升高, 肝胰腺*值在前四個飼料組中顯著升高, 但在第五組中趨于穩定(<0.05); 卵巢*和*值均隨著飼料中β-胡蘿卜素添加量的增加而顯著升高, 但飼料中不同水平β-胡蘿卜素添加量對卵巢*值影響較小。
飼料中添加β-胡蘿卜素對各組織中類胡蘿卜素組成的影響如圖3所示。隨著飼料中β-胡蘿卜素添加量從0 mg/kg增加至100 mg/kg, 蟹殼中蝦青素及β-胡蘿卜素含量顯著增加(<0.05), 但隨著飼料中β-胡蘿卜素添加量從100 mg/kg進一步增加至300 mg/kg, 蟹殼中蝦青素、β-胡蘿卜素及總類胡蘿卜素含量均趨于穩定, 分別穩定于16.7、1.7及29 mg/kg左右。肝胰腺中蝦青素、β-胡蘿卜素及總類胡蘿卜素含量隨著飼料中β-胡蘿卜素添加量從0 mg/kg增加至200 mg/kg而顯著增加(<0.05), 但隨著飼料中β-胡蘿卜素添加量從200 mg/kg進一步增加至300 mg/kg而趨于穩定, 分別穩定于2、116及135 mg/kg左右。卵巢中蝦青素、β-胡蘿卜素及總類胡蘿卜素含量一直隨著飼料中β-胡蘿卜素添加量的增加而顯著增加(<0.05)。
注: a.CAT; b.Trx; c.Prx6; d.GST

圖2 育肥飼料中添加β-胡蘿卜素對雌蟹蟹殼、肝胰腺和卵巢色澤的影響
注: a. 頭胸甲; b. 肝胰腺; c. 卵巢。柱上不同小寫字母表示差異顯著(<0.05)。下同
3 討論
3.1 性腺發育及常規生化
飼料添加類胡蘿卜素對甲殼動物生長性能的影響取決于不同養殖條件、生長階段、特定物種對胡蘿卜素的需求以及不同類胡蘿卜素來源(Wade, 2017; Angell, 2018)。本實驗中雖然β-胡蘿卜素添加組河蟹的最終體重和WGR比對照組高15%左右, 但各飼料組之間生長性能并無顯著差異, 表明飼料中添加β-胡蘿卜素對河蟹生長性能影響較小, 該結果與之前在斑節對蝦(, Boonyaratpalin, 2001)和紅螯螯蝦(, Harpaz, 1998)上的研究結果類似。然而另外一些研究表明, 飼料中添加β-胡蘿卜素可以顯著提高甲殼動物生長性能, 比如Supamattaya等(2005)和Niu等(2014)研究表明飼料中添加β-胡蘿卜素顯著增加了斑節對蝦的平均體質量, Yamada等(1990)和Chien等(1992)研究也表明飼料中添加β-胡蘿卜素可以有效提高日本對蝦生長性能。這些β-胡蘿卜素對生長性能促進效果上的差異可能與不同的養殖條件、研究對象的生長階段、研究對象對β-胡蘿卜素的需求量以及β-胡蘿卜素的來源有關(張飲江等, 2001; Wade, 2017)。性腺和肝胰腺是成熟雌蟹重要的可食部分, 而其性腺指數(GSI)是河蟹卵巢發育的重要指標(Shao, 2014)。關于飼料中添加β-胡蘿卜素對甲殼動物性腺發育的影響, 之前的研究表明飼料中添加β-胡蘿卜素對河蟹卵巢發育的促進作用非常有限(袁春營等, 2007), 這與本研究的結果相一致。

圖3 育肥飼料中添加β-胡蘿卜素對雌蟹蟹殼、肝胰腺和卵巢類胡蘿卜素含量的影響
注: a. 頭胸甲; b. 肝胰腺; c. 卵巢
常規成分是評價河蟹可食組織營養價值的重要指標(Shao, 2013)。在本研究中, 對照組雌蟹肝胰腺中總脂含量顯著低于各β-胡蘿卜素添加組, 說明飼料中添加β-胡蘿卜素有利于河蟹雌體肝胰腺中脂肪的沉積, 該結果與Niu等(2014)的研究一致, 即飼料中添加β-胡蘿卜素顯著增加了斑節對蝦總脂含量。雖然飼料中添加β-胡蘿卜素對甲殼動物常規生化影響的相關研究較少, 但部分學者在飼料中添加蝦青素對甲殼動物營養組成影響方面開展了有益探索, 比如吳仁福等(2018)研究表明飼料中添加蝦青素顯著增加了三疣梭子蟹雌體卵巢和肝胰腺中總脂含量, 此外, Jiang等(2022)研究表明飼料中添加人工合成蝦青素顯著增加了河蟹雌體肝胰腺中的總脂含量。飼料中添加類胡蘿卜素可以增加甲殼動物組織中總脂含量, 可能與類胡蘿卜素具有較少機體氧化應激及能效消耗的特性有關。
3.2 免疫及抗氧化性能
維持生物體復雜的免疫系統需要多種類型的抗氧化酶, 它們構成了針對自由基的酶防御機制的第一道防線。總抗氧化能力(T-AOC)是衡量機體抗氧化系統功能狀況的綜合指標(崔惟東等, 2009), MDA是酶促和非酶促脂質過氧化反應最常見的終產物之一, MDA含量可以作為衡量機體脂質過氧化程度的重要指標(Devasena, 2001)。隨著飼料中β-胡蘿卜素添加量從0 mg/kg增加至300 mg/kg, 河蟹肝胰腺和血淋巴中T-AOC含量呈“低-高-低”的變化趨勢, 并且當飼料中β-胡蘿卜素添加量為100 mg/kg時T-AOC水平最高。與之相反, 隨著飼料中β-胡蘿卜素添加量從0 mg/kg增加至100 mg/kg, 肝胰腺和血淋巴中MDA含量逐漸降低, 而飼料中高β-胡蘿卜素添加量從100 mg/kg進一步增加至300 mg/kg, 肝胰腺和血淋巴中MDA水平異常升高。這些結果表明, 飼料中添加適量β-胡蘿卜素(100 mg/kg左右)可以維持機體活性氧(ROS)產生和抗氧化活性之間的平衡, 但飼料中添加過量β-胡蘿卜素對投喂對象的抗氧化性能產生負面影響, 并存在一定的毒性。Stahl等(2003)研究結果表明, 類胡蘿卜素的抗氧化作用是基于它們可以猝滅單線態氧的特性, 類胡蘿卜素對脂質過氧化的預防作用主要得益于該特性, 此外, 類胡蘿卜素還擁有捕獲過氧自由基的能力, 是過氧自由基的清除劑。類胡蘿卜素與過氧自由基的相互作用可能通過不穩定的β-胡蘿卜素自由基加合物進行, Diet 5中河蟹抗氧化性能的降低可以假設是在抗氧化作用過程中氧化攻擊期間產生大量類胡蘿卜素分解產物, 并且會通過損害線粒體功能來增加氧化應激(Siems, 2005), 但該假設有待進一步驗證。
由于缺乏適應性免疫系統, 甲殼類動物依靠其體液免疫及細胞先天免疫反應來對抗入侵的微生物(Chen, 2019)。AKP和ACP是兩種重要的磷酸酶, 它們在體液免疫中擁有多種功能, 其活性通常可以反映甲殼動物的免疫狀態(Xue, 2000)。飼料中添加100~200 mg/kg β-胡蘿卜素顯著增加了河蟹肝胰腺和血淋巴中AKP及ACP活性, 說明飼料中添加適量β-胡蘿卜素有效提高河蟹免疫性能。β-胡蘿卜素對投喂對象免疫力的提升效果與之前在其他甲殼類動物上的報道類似, 比如, 在飼料中添加0.5%~2%富含β-胡蘿卜素的杜氏鹽藻可以顯著提高斑節對蝦()免疫性能(Madhumathi, 2011), 此外, 在飼料中添加1.5%~3%富含β-胡蘿卜素的杜氏鹽藻也顯著增加了南美白對蝦()免疫性能(Medina Félix, 2017)。
關于飼料中添加β-胡蘿卜素對水生動物抗氧化相關基因表達量影響的研究很少, 本研究結果擬為β-胡蘿卜素如何調節河蟹的抗氧化性能提供一個初步的認識。核因子紅細胞系2相關因子2 (Nrf2)是一種轉錄因子, 通過同時控制內源性抗氧化酶系統中的關鍵成分來調節細胞氧化還原狀態(Hayes, 2014)。先前的研究表明, Nrf2主要通過增加煙酰胺腺嘌呤二核苷酸磷酸(NADPH)的水平來激活依賴于Trx和GSH的抗氧化系統來維持生物體的氧化還原穩態(Bellezza, 2018)。在依賴于Trx的抗氧化系統中, Prx和Trx是關鍵的抗氧化蛋白, 兩者都具有氧化還原酶活性(Mu, 2009a, 2009b)。在依賴于GSH的抗氧化系統中, GST催化GSH與ROS損傷細胞結合, 從而促進其解毒(Mohankumar, 2006)。肝胰腺Prx、Trx和GST的表達水平均隨著β-胡蘿卜素添加量的增加呈現“低-高-低”的變化趨勢, 且在Diet 3組中表達水平最高, 與肝胰腺中抗氧化相關酶活的變化趨勢相一致, 這說明飼料中添加100 mg/kg β-胡蘿卜素有利于維持河蟹的ROS穩態及抗氧化性能的提升。
3.3 色澤及類胡蘿卜素組成
甲殼動物對類胡蘿卜素具有一定的需求量, 它們可以儲存大量類胡蘿卜素并將這些類胡蘿卜素轉化為蝦青素后沉積于肝胰腺、甲殼和卵巢等組織中(Babin, 2019)。成體雌蟹攝食含100~300 mg/kg β-胡蘿卜素的飼料后, 實驗蟹蟹殼、肝胰腺及卵巢中的蝦青素均得到顯著提升, 這驗證了河蟹組織具有將β-胡蘿卜素轉化為蝦青素的能力。隨著飼料中β-胡蘿卜素添加量從0 mg/kg增加至100 mg/kg左右, 蟹殼和肝胰腺中的蝦青素和β-胡蘿卜素含量均顯著增加, 但隨著飼料中β-胡蘿卜素添加量進一步由100 mg/kg增加至200 mg/kg, 蟹殼和肝胰腺中的蝦青素和β-胡蘿卜素含量的變化趨于平緩, 甚至保持不變。該結果表明, 甲殼動物組織中類胡蘿卜素沉積量一旦達到飽和, 飼料中類胡蘿卜素添加量進一步增加將無法有效增加投喂對象組織中類胡蘿卜素含量(Chien, 1992)。與實驗蟹蟹殼和肝胰腺中類胡蘿卜素含量“先增后緩”的變化趨勢不同, 實驗蟹卵巢中蝦青素和β-胡蘿卜素的含量一直隨著飼料中β-胡蘿卜素添加量的增加而顯著增加。這種組織間的差異表明, 同一物種在相同條件下對飼料中β-胡蘿卜素的沉積能力存在一定的組織特異性。
甲殼動物的色澤與其類胡蘿卜素組成密切相關, 飼料中添加β-胡蘿卜素往往可以有效改善甲殼類動物色澤, 進而提高其質量和價格。Boonyaratpalin等(2001)研究表明, 相較于添加50 mg/kg蝦青素的實驗飼料相比, 飼料中添加175 mg/kg β-胡蘿卜素可以更加有效地改善斑節對蝦的色澤。此外, 另一個研究同樣表明在飼料中添加200~300 mg/kg β-胡蘿卜素可以顯著改善斑節對蝦的甲殼色澤(Supamattaya, 2005)。前期的這些研究結果與本實驗結果類似, 即飼料中添加200~300 mg/kg人工合成β-胡蘿卜素顯著增加了成體雌蟹肝胰腺和卵巢紅度值。此外, 各組織紅度值和蝦青素含量均隨著飼料中β-胡蘿卜素添量的增加而同步增加, 這表明實驗蟹各組織所表現出的紅度值增加一方面是由于由β-胡蘿卜素轉化而來的蝦青素含量的增加, 另一方面是β-胡蘿卜素直接誘導的著色作用。
4 結論
綜上所述, 飼料中添加50~300 mg/kg人工合成β-胡蘿卜素對成體雌蟹的生長性能和性腺發育均無顯著影響, 但飼料中添加β-胡蘿卜素可以顯著提高河蟹肝胰腺中總脂含量。就免疫及抗氧化相關指標而言, 飼料中添加適量β-胡蘿卜素(50~100 mg/kg)可以顯著提高成體雌蟹的免疫及抗氧化性能, 但添加過量β-胡蘿卜素(200~300 mg/kg)將影響河蟹免疫及抗氧化性能。此外, 飼料中添加適量β-胡蘿卜素顯著提高了河蟹蟹殼、肝胰臟和卵巢中蝦青素和β-胡蘿卜素含量, 但飼料中添加β-胡蘿卜素僅顯著增加了河蟹肝胰腺和卵巢紅度值。綜合考慮成本、著色效果、抗氧化性能和免疫力等因素, 建議成體雌蟹育肥飼料中人工合成β-胡蘿卜素的添加量為100 mg/kg左右。
王世會, 成永旭, 賈智英, 等, 2021. 兩種肝胰腺色澤中華絨螯蟹營養品質比較[J]. 水產學雜志, 34(4): 27-34.
吳仁福, 龍曉文, 侯文杰, 等, 2018. 飼料中添加雨生紅球藻粉對三疣梭子蟹雌體卵巢發育、色澤、抗氧化能力和生化組成的影響[J]. 水生生物學報, 42(4): 698-708.
張世勇, 傅洪拓, 喬慧, 等, 2013. 中華絨螯蟹遺傳育種研究進展[J]. 中國農學通報, 29(20): 39-45.
張飲江, 何培民, 何文輝, 2001. 螺旋藻對中華絨螯蟹生長和體色的影響[J]. 中國水產科學, 8(2): 59-62.
袁春營, 崔青曼, 2007. β-胡蘿卜素對中華絨螯蟹卵巢發育及免疫學指標的影響[J]. 海洋科學, 31(6): 25-28.
崔惟東, 冷向軍, 李小勤, 等, 2009. 蝦青素和角黃素對虹鱒肌肉著色和肝臟總抗氧化能力的影響[J]. 水產學報, 33(6): 987-995.
ANGELL A, DE NYS R, MANGOTT A,, 2018. The effects of concentration and supplementation time of natural and synthetic sources of astaxanthin on the colouration of the prawn[J]. Algal Research, 35: 577-585.
AOAC, 2000. Official Methods of Analysis [M]. Gaithersburg: Association of Official Analytical Chemists.
BABIN A, MOREAU J, MORET Y, 2019. Storage of carotenoids in crustaceans as an adaptation to modulate immunopathology and optimize immunological and life‐history strategies [J]. BioEssays, 41(11): 1800254.
BELLEZZA I, GIAMBANCO I, MINELLI A,, 2018. Nrf2-Keap1 signaling in oxidative and reductive stress [J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 1865(5): 721-733.
BOONYARATPALIN M, THONGROD S, SUPAMATTAYA K,, 2001. Effects of β-carotene source,, and astaxanthin on pigmentation, growth, survival and health of[J]. Aquaculture Research, 32(S1): 182-190.
CHEN F Y, WANG K J, 2019. Characterization of the innate immunity in the mud crab[J]. Fish & Shellfish Immunology, 93: 436-448.
CHIEN Y H, JENG S C, 1992. Pigmentation of kuruma prawn,Bate, by various pigment sources and levels and feeding regimes [J]. Aquaculture, 102(4): 333-346.
DEVASENA T, LALITHA S, PADMA K, 2001. Lipid peroxidation, osmotic fragility and antioxidant status in children with acute post-streptococcal glomerulonephritis [J]. Clinica Chimica Acta, 308(1/2): 155-161.
FOLCH J, LEES M, SLOANE STANLEY G H, 1957. A simple method for the isolation and purification of total lipides from animal tissues [J]. Journal of Biological Chemistry, 226(1): 497-509.
GUL K, TAK A, SINGH AK,, 2015. Chemistry, encapsulation, and health benefits of β-carotene-A review [J]. Cogent Food & Agriculture, 1(1): 1018696.
HARPAZ S, RISE M, ARAD S M,, 1998. The effect of three carotenoid sources on growth and pigmentation of juvenile freshwater crayfish[J]. Aquaculture Nutrition, 4(3): 201-208.
HAYES J D, DINKOVA-KOSTOVA A T, 2014. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism [J]. Trends in Biochemical Sciences, 39(4): 199-218.
JIANG M, ZHAO H H, ZAI S W,, 2019. A defatted microalgae meal () as a partial protein source to replace fishmeal for feeding juvenile yellow perch[J]. Journal of Applied Phycology, 31(2): 1197-1205.
JIANG X D, PAN K W, YANG Y H,, 2022. Dietary DHA oil supplementation promotes ovarian development and astaxanthin deposition during the ovarian maturation of Chinese mitten crab[J]. Aquaculture Nutrition, 2022: 9997317.
JIANG X D, ZU L, WANG Z Y,, 2020. Micro-algal astaxanthin could improve the antioxidant capability, immunity and ammonia resistance of juvenile Chinese mitten crab,[J]. Fish & Shellfish Immunology, 102: 499-510.
JOHNSTON I A, ALDERSON R, SANDHAM C,, 2000. Muscle fibre density in relation to the colour and texture of smoked Atlantic salmon (L.) [J]. Aquaculture, 189(3/4): 335-349.
KONG L, CAI C F, YE Y T,, 2012. Comparison of non-volatile compounds and sensory characteristics of Chinese mitten crabs () reared in lakes and ponds: Potential environmental factors [J]. Aquaculture, 364-365: 96-102.
LEóN K, MERY D, PEDRESCHI F,, 2006. Color measurement inunits from RGB digital images [J]. Food Research International, 39(10): 1084-1091.
LIU L, LONG X W, DENG D,, 2018. Molecular characterization and tissue distribution of carnitine palmitoyltransferases in Chinese mitten craband the effect of dietary fish oil replacement on their expression in the hepatopancreas [J]. PLoS One, 13(8): e0201324.
LONG X W, WU X G, ZHAO L,, 2017. Effects of dietary supplementation withcell powder on coloration, ovarian development and antioxidation capacity of adult female Chinese mitten crab,[J]. Aquaculture, 473: 545-553.
MADHUMATHI M, RENGASAMY R, 2011. Antioxidant status offed withsupplemented diet and resistance against WSSV [J]. International Journal of Engineering, Science and Technology, 3(10): 7249-7259.
MARTíN J F, GUDI?A E, BARREDO J L, 2008. Conversion of β-carotene into astaxanthin: Two separate enzymes or a bifunctional hydroxylase-ketolase protein? [J]. Microbial Cell Factories, 7(3): 3.
MEDINA FéLIX D, ELíAS J A L, CóRDOVA á I C,, 2017. Survival ofshrimp fed on diets supplemented withsp. is improved after challenges by[J]. Journal of Invertebrate Pathology, 148: 118-123.
MOHANKUMAR K, RAMASAMY P, 2006. White spot syndrome virus infection decreases the activity of antioxidant enzymes in[J]. Virus Research, 115(1): 69-75.
MU C K, ZHAO J M, WANG L L,, 2009a. A thioredoxin with antioxidant activity identified from[J]. Fish & Shellfish Immunology, 26(5): 716-723.
MU C K, ZHAO J M, WANG L L,, 2009b. Molecular cloning and characterization of peroxiredoxin 6 from Chinese mitten crab[J]. Fish & Shellfish Immunology, 26(6): 821-827.
NIU J, WEN H, LI C H,, 2014. Comparison effect of dietary astaxanthin and β-carotene in the presence and absence of cholesterol supplementation on growth performance, antioxidant capacity and gene expression ofunder normoxia and hypoxia condition [J]. Aquaculture, 422-423: 8-17.
PENG J, XIANG W Z, TANG Q M,, 2008. Comparative analysis of astaxanthin and its esters in the mutant E1 ofand other green algae by HPLC with a C30 column [J]. Science in China Series C: Life Sciences, 51(12): 1108-1115.
QIU R J, CHENG Y X, HUANG X X,, 2011. Effect of hypoxia on immunological, physiological response, and hepatopancreatic metabolism of juvenile Chinese mitten crab[J]. Aquaculture International, 19(2): 283-299.
SHAO L C, WANG C, HE J,, 2013. Hepatopancreas and gonad quality of Chinese mitten crabs fattened with natural and formulated diets [J]. Journal of Food Quality, 36(3): 217-227.
SHAO L C, WANG C, HE J,, 2014. Meat quality of Chinese mitten crabs fattened with natural and formulated diets [J]. Journal of Aquatic Food Product Technology, 23(1): 59-72.
SIEMS W, WISWEDEL I, SALERNO C,, 2005. β-Carotene breakdown products may impair mitochondrial functions— potential side effects of high-dose β-carotene supplementation [J]. The Journal of Nutritional Biochemistry, 16(7): 385-397.
STAHL W, SIES H, 2003. Antioxidant activity of carotenoids [J]. Molecular Aspects of Medicine, 24(6): 345-351.
SUPAMATTAYA K, KIRIRATNIKOM S, BOONYARATPALIN M,, 2005. Effect of aextract on growth performance, health condition, immune response and disease resistance in black tiger shrimp () [J]. Aquaculture, 248(1/2/3/4): 207-216.
WADE N M, GABAUDAN J, GLENCROSS B D, 2017. A review of carotenoid utilisation and function in crustacean aquaculture [J]. Reviews in Aquaculture, 9(2): 141-156.
WADE N, GOULTER K C, WILSON K J,, 2005. Esterified astaxanthin levels in lobster epithelia correlate with shell colour intensity: Potential role in crustacean shell colour formation [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 141(3): 307-313.
WANG M, WANG L, ZHOU Z,, 2013. The molecular characterization of a catalase from Chinese mitten crab[J]. International Journal of Immunogenetics, 40(3): 230-240.
WEI J J, ZHANG F, TIAN W J,, 2016. Effects of dietary folic acid on growth, antioxidant capacity, non-specific immune response and disease resistance of juvenile Chinese mitten crab(Milne‐Edwards, 1853) [J]. Aquaculture Nutrition, 22(3): 567-574.
XUE Q G, RENAULT T, 2000. Enzymatic activities in European flat oyster,, and Pacific oyster,, hemolymph [J]. Journal of Invertebrate Pathology, 76(3): 155-163.
YAMADA S, TANAKA Y, SAMESHIMA M,, 1990. Pigmentation of prawn () with carotenoids: I. Effect of dietary astaxanthin,-carotene and canthaxanthin on pigmentation [J]. Aquaculture, 87(3/4): 323-330.
ZHAO D X, CHEN L Q, QIN C J,, 2010. A delta-class glutathione transferase from the Chinese mitten crab: cDNA cloning, characterization and mRNA expression [J]. Fish & Shellfish Immunology, 29(4): 698-703.
EFFECT OF DIETARY β-CAROTENE ON OVARY DEVELOPMENT, ANTIOXIDANT CAPABILITY, IMMUNITY, AND TISSUE COLOR OF ADULT FEMALE CHINESE MITTEN CRAB,
JIANG Xiao-Dong1, 2, 3, ZHU Ling-Xiang1, 2, 3, CHENG Yong-Xu1, 2, 3, WU Xu-Gan1, 2, 3
(1. Key Laboratory of Freshwater Aquatic Genetic Resources, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China; 2. Shanghai Collaborative Innovation for Aquatic Animal Genetics and Breeding, Shanghai Ocean University, Shanghai 201306, China; 3. Fish Nutrition and Environmental Ecology Research Center, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China)
To investigate the effects of dietary supplementation of synthetic β-carotene on weight gain rate (WGR), gonadosomatic index (GSI), hepatosomatic index (HSI), proximate composition, antioxidant status, color parameters, and carotenoid content of adult female Chinese mitten crab, five diets containing 3.3, 56.7, 108.9, 207.9, and 320.0 mg/kg β-carotene (denoted as Diets 1~5, respectively) were formulated to feed.that have completed puberty molting.Results show that: (1) Although the WGR, specific growth rate (SGR) and HSI of adult female.was not affected by dietary β-carotene supplementation, significant promotion of lipid content was observed in the hepatopancreas of β-carotene supplemented treatments. (2) Indicators of antioxidant capacity (T-AOC, MDA, GSH-Px) and immunity (AKP, ACP) in the hepatopancreas and hemolymph showed an apparent trend of “low-high-low” with the increase of dietary β-carotene supplementation, and the highest levels were observed in the 100 to 300 mg/kg treatments. (3) Dietary β-carotene significantly improved the redness (*) of carapace and hepatopancreas, which agreed with the significantly increased astaxanthin and total carotenoid concentrations in these tissues from different treatments. Therefore, the supplement ofβ-carotene in diet had no effect on WGR and gonad development of adult female., but enhance the antioxidative capability, non-specific immunity, color and luster, and tissue carotenoid contents. Based on these results, the optimal β-carotene content in the diets was around 100 mg/kg for adult female.. The research findings not only contribute to understanding the impact of β-carotene on the physiological functions of crustaceans but also facilitate the development of functional feeds for..
; fattening feed; carotenoid; culture performance; color parameters
* 國家自然科學基金面上項目, 31873041號; 科技部藍色糧倉項目, 2018YFD0900103號;上海市河蟹產業體系專項, 滬農科產字(2022)第4號。姜曉東, 博士后, E-mail: 310410555@qq.com
吳旭干, 博士生導師, 教授, E-mail: xgwu@shou.edu.cn
2023-04-06,
2023-07-02
S963.73
10.11693/hyhz20230400079
