多環境因子對微藻生長競爭影響的室內模擬研究綜述
2023-12-13 02:28:48葛裕豪劉雪梅吳燕鋒
環境科技 2023年6期
葛裕豪,劉雪梅,楊 旭,吳燕鋒 張 婧
(1.黑龍江大學 水利電力學院,黑龍江 哈爾濱 150080;2.中國科學院東北地理與農業生態研究所,吉林 長春 130102;3.黑龍江大學 信息管理學院,黑龍江 哈爾濱 150080)
0 引言
微藻是一類包括綠藻、藍藻、硅藻、金藻、裸藻、甲藻等多個門類的單細胞或多細胞光合水生微生物,除藍藻門外均屬于真核生物[1]。微藻作為水生生態系統的初級生產者,為水生食物鏈和食物網提供物質和能量支撐,是水生生態系統的重要組成部分[2]。適量微藻可提高水體中溶氧量和魚類食物,促進水產養殖生物的生長和繁殖;過量微藻則會引發藻華,降低水質、透明度和自凈能力,并通過產生的藻毒素威脅水生生物和人畜健康[1]。此外,由于微藻的生長繁殖受氣候變化和人類活動的雙重影響,多環境因子(物理、化學和生物等)對微藻的生長、競爭機制尚不清晰,故給水體藻華的有效治理帶來了挑戰[3-4]。為了深入了解多環境因子對不同微藻種類生長、競爭的影響機制和規律,通過綜述國內、外近十年關于多環境因子對不同微藻種類生長、競爭影響的室內模擬研究進展,提出未來的研究方向和展望。室內模擬是揭示多環境因子對微藻生長、競爭機制的有效方式之一[5-7],通過模擬不同的生態環境,研究微藻的生理特征和生長響應機制,也可通過模擬微藻的生物量積累和競爭過程,為水華的形成、維持和消退提供理論依據。
1 物理因素對微藻生長競爭的室內模擬試驗
對微藻產生影響的物理因素主要包括光、溫度和擾動等因子,通過作用于藻種光合作用、呼吸速率、硝化速率和反硝化速率等生物地球化學過程來影響藻種生長競爭。物理因子實驗的設置梯度及對象見表1。

表1 物理因子實驗的設置梯度及對象
1.1 光照
在影響微藻生長、競爭的環境因素中,光照可通過作用于微藻的光合作用、色素合成、酶活性等方面來調控微藻的代謝速率、生物量積累和營養吸收等生理過程[8]。如:王菁等[9]對普通小球藻(Chlorella vulgaris)和魚腥藻(A nabaena sp.)的研究發現,魚腥藻和普通小球藻的最大細胞濃度均隨光照強度的增加而升高,但兩者的最適光強不同,且在共同培養時,兩者之間的競爭抑制參數隨著光照強度而變化[10]。除光照強度,其他光照條件(如光照周期、光照頻率、光照波長和光照強度波動等) 也可對微藻生長和競爭產生影響。如:XU Y 等[11]研究發現,鹽生杜氏藻(Dunaliella salina)的生長速率和生物量均隨光照時間的延長而增加。李默楠等[12]研究發現,當光照波長組合分別為450~465 和650~660 nm 時,鈍頂螺旋藻(Spirulina platensis)的產量達到最高,同時紅光對螺旋藻的影響大于藍光。卿人韋等[13]研究發現,在光照周期性波動的條件下,銅綠微囊藻(Microcystis aeruginosa)和斜生柵藻(Scenedesmus obliquus)的生物量、色素含量和酶活性均有所降低。由此可見,光照對不同藻種的生長和競爭均有顯著影響,不同藻種對光照的需求和利用有所差異,光照變化可導致微藻群落結構和功能的變化[14]。因此,了解不同藻種的最適光強和光照對微藻競爭的影響機制,對于水體富營養化治理和水產養殖有重要意義。
1.2 溫度
溫度是影響微藻生長和競爭的重要環境因子之一,通過改變細胞膜流動性、脂質組成和酶活性等方式,可調節微藻的代謝速率、光合效率和營養吸收等生理過程。溫度變化可導致微藻群落結構和功能變化,過高或過低的溫度均可抑制微藻的生長[8,15-16]。王菁等[16]研究發現,普通小球藻單種培養時,該藻的最大細胞濃度隨溫度的增加而增加,35 ℃為最適生長溫度;而魚腥藻在單獨培養時,30~35 ℃為最適生長溫度。李曉莉等[17]研究發現,舟形藻(Navicula pelliculosa)在單種培養中,其最大細胞濃度在25~30 ℃時達到最大值,而達到35 ℃時則停止生長。除了溫度數值變化,其他溫度條件(如溫度波動、溫度梯度、溫度周期等)也可對微藻生長和競爭產生影響。DESCAMPS-JULIEN 等[18]研究發現,小環藻(Cyclotella)和脆桿藻(Fragilaria)在波動溫度下均表現出相互補償動態,導致了它們共存。水溫的增加使微藻更容易獲取和利用光能,增加光抑制發生的光能水平,導致深層水體中光能限制的情況更加嚴重,微藻生長受到光限制的深度變得更淺[19]。因此,研究溫度對微藻生長和競爭的影響,需考慮多種溫度因素和多種微藻物種及它們之間的相互作用。
1.3 水力條件
水力條件是影響藻華爆發的重要因素之一。湖泊作為主要的水華發生水體,其流速通常較低,較少超過0.2 m/s[20],受外界影響通常可導致其水力條件發生變化。相關研究表明,水流對單種培養和共同培養的微藻生長和競爭均有顯著影響,不同藻種對水流的需求和利用均有差異,水流改變可導致優勢藻種的變化[21-23]。銅綠微囊藻生長的最適流速為0.15 m/s,相對有利流速為低流速(0.05~0.12 m/s),相對不利流速為高流速(0.20~0.30 m/s)[21]。野外觀測表明,水動力降低容易發生藻華,如:王紅萍等[24]研究發現,當漢江流速低于0.225 m/s 時,硅藻水華容易發生。趙孟緒等[25]認為,湯溪水庫汛期時水體反常的穩定性是導致水華提前暴發的關鍵因子。國外學者也嘗試采用改變水動力的方法來控制微藻水華,如:HEO W M 等[26]研究發現,揚水曝氣裝置可抑制Dalbang湖藍藻生長,由于湖水深度較淺并不會對浮游植物的總生物量產生影響。TSUKADA H 等[27]在Yogo 湖采用揚水曝氣裝置發現,改變水動力可控制有害藍藻,使原本藍藻優勢轉變為硅藻優勢。一些學者通過室內模擬探究水動力對微藻生長的作用機理[28],湖泊水動力的改變可引起一系列復雜變化(如:①顆粒物沉降、水體透明度變化;②水體光合作用強度變化;③微藻的自然沉降速率、絮凝等變化)。王華等[29]研究表明,小擾動有利于微藻生長的原因:一方面是因為小擾動可提高微藻營養物質的利用率。且可降低藻細胞生長到一定階段產生次級代謝產物的濃度,減少對其生長的抑制作用;另一方面擾動可防止綠藻自然的沉降。而靜止狀態與高強度擾動對微藻生長均有明顯的抑制作用。王藝兵等[30]認為,擾動本身對微藻生長影響不大,在擾動速度為0.1 m/s 時有微弱的促進作用,在擾動速度為0.2~0.5 m/s 時有較大的促進作用;因此,水體擾動速率的降低并不是藍藻水華快速生長的直接原因,其它因素(如懸浮物濃度降低)引起光合作用增強,補充碳源,水動力降低有利于藍藻發揮浮力機制而形成優勢藻種等間接原因是藍藻水華暴發的主要誘因[30]。
2 化學因素對微藻生長競爭的室內模擬試驗
2.1 營養鹽
微藻作為河流水質狀態與生態健康的重要指示生物,其種類、數量和群落結構均主要受營養鹽水平影響。其中,N,P 等營養元素是浮游微藻生長的必需營養物質,也是水體富營養化的主導因素。當水體中TN 質量濃度大于0.2 mg/L,TP 質量濃度大于0.02 mg/L 時,水體為富營養化狀態[31]。除了N,P 的絕對濃度外,氮磷比的變化也會明顯改變水體中營養鹽的限制狀態,對微藻生長及群落結構演替具有重要影響。在一定Fe2+濃度條件下,微藻生長速率v1>v2>v3>v4(v1,v2,v3和v4時的ρ(N)∶ρ(P)分別為40∶1,80∶1,20∶1 和5∶1),最佳ρ(N)∶ρ(P)為40∶1[32]。①當ρ(N)∶ρ(P)小于7~10 時,普遍認為水體中微藻生長處于氮限制狀態;②當ρ(N)∶ρ(P)大于22.6~30時,則認為其處于磷限制狀態;③當ρ(N)∶ρ(P)介于以上兩者之間時,為微藻的最適生長區間[33]。水體中無機磷濃度超過微藻實際需求量時,可造成微藻的物質成分(如葉綠素a 含量)發生變化,進而限制其生長[34]。磷脅迫對有毒微藻產毒代謝的影響尚不明確,有毒微藻的產毒基因在不同磷源條件下表達有差異[35]。ZHOU Y P 等[36]認為,氮與產毒有關,磷可能只是協同作用。李璇等[37]認為,有毒微藻在磷缺乏環境中啟動各種磷利用機制,通過調節細胞內儲存或利用外源有機磷(如提高磷的轉運速率或增加有機磷的水解等)。因此,探究不同磷源對有毒微藻生長和產毒代謝的影響及其分子機理,對于理解和控制水華暴發具有重要意義。
2.2 pH 值
除以上影響因素外,藻類生長還與pH 值、Fe2+和CO2濃度等因素有關。pH 值是影響藻類生長和競爭的重要環境因子,其通過影響藻類的碳酸氫鹽平衡、光合作用、色素合成和酶活性等方式調節藻類的生理過程(如生長速率、生物量積累和營養吸收等)。許海等[38]研究發現,3 種淡水藍藻(銅綠微囊藻、魚腥藻和斜生柵藻)適宜的pH 值分別為9.0,8.0 和10.0,而3 種淡水綠藻(浮游顫藻、綠球藻和雷氏衣藻)適宜的pH 值分別為7.0,8.0 和7.0。其他pH 值條件(如pH 值波動、pH 值梯度和pH 值周期等) 可對藻類生長和競爭產生影響。不同pH 值波動也可對魚腥藻和普通小球藻的生長和競爭產生影響,王菁[10]研究發現,pH 值在周期性波動下,魚腥藻和普通小球藻的生物量、色素含量和酶活性均有所下降。王藝兵等[30]研究發現,四尾柵藻對pH 值適宜范圍較廣,具有較強的耐酸耐堿性;惠氏微囊藻適宜在偏堿性的水體中生長;菱形藻更適宜在偏中性的水體生長。在碳源充足(pH 值≤8.3)情況下,對硅藻生長有微弱的促進作用;在碳源比較缺乏(pH 值>8.3)情況下,3 種藻在高擾動下的生長速率均較高[30]。然而,試驗中對培養基的pH 值調節較為困難,pH 值調節需使用適當的酸堿物質(如稀鹽酸、稀氫氧化鈉、碳酸氫鈉等),酸堿物質可能對藻類本身或其他水質參數(如滲透壓、電導率、溶解氧等)產生影響;需考慮水體中其他物質(如CO2,HCO3-,HPO42-等)對pH 值的影響,以上物質可能與酸堿物質發生反應或緩沖作用,影響pH 值變化[39]。
2.3 水中Fe2+和CO2 濃度
鐵元素作為藻類生長的必需元素,在適宜的氮磷比條件下,Fe2+質量濃度為0 時,藻類基本不生長;Fe2+質量濃度為1.2 mg/L 時,藻類的葉綠素含量和增長速率均最大;Fe2+質量濃度達到4.8 mg/L 時,藻類生長則受到抑制[32]。因此,除控制氮磷比外,控制水體中鐵元素含量也可控制或減弱水華發生強度。藻類光合作用的高效性依賴于羧化酶RuBisCO 催化位點的CO2聚集機制(CCM),這種機制有助于CO2的固定[40]。CO2濃度變化可改變水體中不同形式碳源的比例,從而影響不同藻種對碳源的利用效率和競爭力。當CO2濃度增加時,具有CCM 的藻種相對于CCM 的藻種會處于劣勢,原因是因為CCM 需要消耗能量,而高濃度下CO2則能量供應不足;反之,當CO2濃度降低時,具有CCM 的藻種相對于CCM的藻種會處于優勢。此外,不同類型的CCM 也會影響不同藻種之間的競爭力。在高pH 值下,具有類囊體CCM 可利用水體中較豐富的碳酸根作為碳源,而細胞質CCM 只能利用水體中較稀缺的游離CO2作為碳源,因此,具有類囊體CCM 的綠藻相對于具有細胞質CCM 的硅藻會處于優勢[40]。
化學因子試驗的設置情況見表2。

表2 化學因子試驗的設置情況
3 應對多環境因素變化的室內模擬試驗策略
微藻室內模擬研究概述示意見圖1。
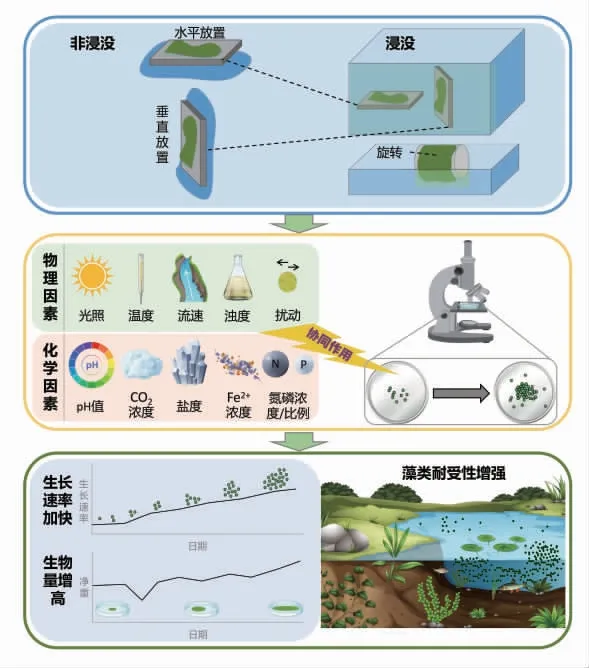
圖1 室內模擬試驗概述示意
目前,單因子模擬研究較多,而缺乏多因子的綜合模擬研究,并且關于溶藻細菌、沉水植物、挺水植物和浮游植物等微生物和水生植物的化感物質[41-42]對微藻影響的室內模擬控制試驗均較少。研究微藻生長、競爭機制可為水體治理富營養化提供新的思路。
(1)不同培養反應器
附著式微藻培養是一種利用固定載體讓微藻附著生長的培養方式,相比懸浮微藻培養,可節省水資源、減少占地面積、提高生物產量和水處理效率。為提高附著式微藻培養的生產力和質量,建議:①利用發光細菌提供散射光,改善微藻生物膜的光照環境;②建立綜合評價體系,對附著微藻培養系統的生產力、化學成分、酶活性等指標進行全面和可比較的評價;③深入研究微藻生物膜內的物質轉移、EPS 分泌、基因表達等過程,為附著微藻培養系統的優化和規模化提供理論依據。
(2)單藻種
室內模擬試驗的未來發展方向主要包括:探討物種與環境因子的關聯性和競爭性;設計高效、低成本的新型光生物反應器;優化適宜工業化應用的藻種;開發微藻在能源、環境、食品和醫藥等領域的應用。此外,室內模擬試驗還可結合分子生物學、遺傳學、代謝組學等技術,深入研究微藻在不同環境壓力下的分子機制,揭示微藻的響應模式和適應策略。
(3)多藻種
因室內模擬試驗存在著不足和挑戰,無法反映真實環境中存在的多種復雜因素和隨機事件的影響;該種試驗通常只觀察和測量部分或表面的微藻特征和指標,無法深入了解其內在的分子機制和調控網絡;只涉及部分或局部的微藻物種和群落,無法反映其在全球或區域尺度上的多樣性和分布以及與其他水生生物之間的相互作用。未來在多環境因子下的室內模擬試驗可從以上不足之處進行改進和突破。
4 結論及展望
通過對藻類室內模擬試驗大量文獻的綜述,總結了物理因素(光照、溫度、流速、濁度和擾動等)和化學因素(pH 值、CO2濃度、鹽度、Fe2+濃度、N,P 濃度及其化學計量比等) 協同作用導致藻類的快速增殖并引起藻華。由于以上環境因子之間存在著復雜的相互作用和協同效應,故影響著微藻的生理代謝、生物化學組成、群落結構和功能。通過綜述多環境因子對微藻生長、競爭影響的室內模擬研究的主要內容、方法和進展,總結出目前存在的問題和不足,并對未來研究方向進行展望:
(1)增加試驗設計的復雜性和實際性,考慮更多的環境因子和其組合效應,模擬更接近自然環境的條件,提高試驗結果的可靠性和適用性。
(2)引入更多的微藻種類和群落類型,探索不同微藻之間的相互作用和競爭機制,揭示微藻群落結構和功能對環境因子變化的響應規律。
(3)利用分子生物學、遺傳學、代謝組學等高通量技術,深入研究微藻在不同環境壓力下的分子機制和調控網絡(如基因表達、信號轉導、代謝調節等),并通過轉錄組、蛋白質組、代謝組等多組學分析,揭示微藻的響應模式和適應策略。
(4)結合數值模擬和人工智能等技術,構建湖泊氣候-水文-水質-藻華機理模型,利用室內模擬試驗數據進行參數化和驗證,量化多環境因子對微藻生長競爭的影響及貢獻率,預測未來氣候變化情景下湖泊微藻群落的變化趨勢和生態效應。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
