芝麻蛋白Ses i 3的重組表達及致敏性鑒定
2023-12-14 12:45:20姜松松湯鑫磊黃雨彤虞丹丹孟祥忍
食品科學 2023年22期
關鍵詞:小鼠
姜松松,王 濤,湯鑫磊,李 倩,黃雨彤,虞丹丹,孟祥忍,2,*
(1.揚州大學旅游烹飪學院,江蘇 揚州 225127;2.揚州大學中餐繁榮基地,江蘇 揚州 225127)
芝麻過敏是食物過敏的一種,是機體攝入芝麻或芝麻制品引起的一種異常免疫反應。近年來,隨著芝麻及其制品銷量增多,芝麻過敏患者也逐漸增多,在世界范圍內引起廣泛關注。流行病學調查結果顯示,芝麻是中東地區的主要食物過敏原之一,是以色列的第三大食物過敏原[1]。目前,芝麻已經被許多國家列入主要食物過敏原清單,如法國(芝麻過敏發病率為3%)[2]、澳大利亞(芝麻過敏發病率為0.8%)[3]和美國(芝麻過敏發病率為0.49%)[4]等,加拿大明確要求在食品包裝上必須標注芝麻過敏原信息[5]。芝麻是我國常見的油料作物和加工食品原料,在我國食品和餐飲行業中廣泛應用,消費量巨大。目前,我國雖然還沒有準確的芝麻過敏流行病學調查報告,但一項對食物過敏患者過敏原調查結果顯示,芝麻過敏發病率位列第三,僅次于小麥和雞蛋,芝麻是7~17 歲年齡段食物過敏患者最主要的食物過敏原,且發病率逐年升高[6]。此外,研究發現過敏患者攝入極少量的芝麻即可誘發嚴重的過敏臨床癥狀,如蕁麻疹、皮膚水腫瘙癢、腹瀉等,嚴重的甚至可能誘發過敏性休克,導致死亡[7]。2022年,綜合芝麻過敏的發病率、臨床癥狀嚴重程度和潛在危害考慮,聯合國糧食及農業組織和世界衛生組織將其列入新“8大類”食物過敏原之一[8]。由此可見,芝麻過敏已經成為一個不容忽視的公共健康問題。
目前,已經被鑒定出來的芝麻過敏原蛋白有7 種,包括2S貯藏白蛋白(Ses i 1和Ses i 2)、7S類豌豆球蛋白(Ses i 3)、油質蛋白(Ses i 4和Ses i 5)和11S球蛋白(Ses i 6和Ses i 7)[9]。其中,Ses i 3由585 個氨基酸組成,分子質量約為45 kDa,屬于Cupin超家族,具有β-折疊桶的蛋白質超二級結構,由單個肽鏈的3 個單體結合成三角形扁平三聚體,能夠抵抗熱變性和酶消化,可被75%芝麻過敏患者的血清識別,是芝麻主要的過敏原之一[10]。
高純度的芝麻蛋白Ses i 3在致敏性及致敏機理的相關研究中具有不可或缺的地位。然而,從芝麻中提取高純度天然Ses i 3會遇到不可避免的問題。繁雜的提取步驟可能會使蛋白降解損失,同時活性下降。在提純過程中,過敏原蛋白與非過敏原蛋白同時存在,制備高純度單一過敏原蛋白的難度系數較大。此外,天然過敏原蛋白在保存過程中,部分酶可能會將其降解,導致其生物活性降低[11]。這些原因使提取天然過敏原蛋白難以標準化和量產化。相比之下,通過重組技術獲得的食物過敏原蛋白可以較好地規避這些問題。近年來,因重組蛋白具有純度高、產量大及理化和免疫學性質與天然過敏原蛋白相當等特點,研究學者進行了多種食物過敏原蛋白的重組表達研究,并取得較理想的結果[12-13]。
因此,本研究從芝麻中提取總RNA,用反轉錄聚合酶鏈式反應(reverse transcription-polymerase chain reaction,RT-PCR)得到芝麻蛋白Ses i 3基因,構建pET-22b(+)質粒表達載體,轉入BL21(DE3)感受態細胞宿主表達菌中誘導表達,對芝麻過敏原Ses i 3蛋白進行重組表達和純化,并進一步建立BALB/c過敏小鼠模型對其免疫學特性進行鑒定,以期為芝麻過敏蛋白Ses i 3的量產化提供理論依據,同時也為后續芝麻致敏機理和芝麻脫敏研究奠定基礎。
1 材料與方法
1.1 材料與試劑
LB平板組成成分:氯化鈉10 g/L、胰蛋白胨10 g/L、酵母提取物5 g/L,121 ℃滅菌20 min,固體培養基中另加入質量分數1.5%瓊脂粉。
天然芝麻蛋白Ses i 3(純度≥90%),實驗室制備;辣根過氧化物酶(horseradish peroxidase,HRP)-羊抗鼠免疫球蛋白G1(immunoglobulin G1,IgG1)、HRP-羊抗鼠免疫球蛋白G2a(immunoglobulin G2a,IgG2a)、HRP-羊抗鼠免疫球蛋白E(immunoglobulin E,IgE)賽默飛世爾科技(中國)有限公司;白細胞介素-4(interleukin-4,IL-4)、白細胞介素-5(interleukin-5,IL-5)、干擾素-γ(interferon-γ,IFN-γ)、組胺酶聯免疫吸附測試(enzyme-linked immunosorbent assay,ELISA)試劑盒 上海寶曼生物科技有限公司;PrimeScriptTMRT Master Mix、瓊脂糖凝膠回收試劑盒、質粒小量提取試劑盒、限制性內切酶NdeI、XhoI、XbaI 通用生物(安徽)有限公司;HRP anti-6×His tag小鼠單克隆抗體英國Abcam公司。
1.2 儀器與設備
1600凝膠成像分析系統、EPS200蛋白垂直電泳槽上海天能科技有限公司;JY99-IIDN超聲波細胞粉碎機 寧波新芝生物科技股份有限公司;GHP-9160培養箱 上海一恒科學儀器有限公司;TGL-16M臺式高速冷凍離心機 上海盧湘儀離心機儀器有限公司;Multiskan FC酶標儀 賽默飛世爾科技(中國)有限公司。
1.3 方法
1.3.1 pET-22b(+)-Ses i 3表達載體的構建
根據GenBank數據庫中芝麻過敏原Ses i 3蛋白的基因序列設計擴增引物。上游引物:5’-AATTTTGTTTAACTT TAAGAAGGAGATATACATATGGAAAGCAAAGATCCG GAACTGAAACAGTGCAAAC-3’;下游引物:3’-AGCC GGATCTCAGTGGTGGTGGTGGTGGTGCTCGAGGGCA TCTGCGCGACCCTGCTGCTGCTGGCGCGGGCC-5’。
用Trizol法提取芝麻總RNA[14],將芝麻在液氮中充分研磨后,取50 mg加入1 mL的Trizol混勻,隨后轉移至RNase-free離心管中室溫放置5 min。加入0.2 mL氯仿后劇烈搖晃15 s,室溫放置3 min。4 ℃、12 000 r/min離心15 min后,將上清液轉移至新的RNase-free離心管,加入0.5 mL異丙醇,混勻后室溫靜置10 min。4 ℃、12 000 r/min離心10 min,棄上清液,加入1 mL的75%乙醇溶液洗滌沉淀。4 ℃、7 000 r/min離心5 min,棄上清液,放置5 min,加入50 μL無RNase水溶解,-80 ℃保存。
用PrimeScriptTMRT Master Mix試劑進行反轉錄反應,反應體系為5×PrimeScript RT Master Mix 2 μL,RNA≤500 ng,加入無RNase的雙蒸水至10 μL。37 ℃孵育15 min,85 ℃、5 s使反轉錄酶失活。
反應體系:GPV8高保真DNA聚合酶0.5 μL,10×GPV8 buffer 5 μL,10 mmol/L dNTP Mix 1 μL,上下游引物2×2 μL,模板DNA 2 μL,雙蒸水37.5 μL。擴增條件:96 ℃預變性3 min;95 ℃變性15 s,58 ℃退火15 s,72 ℃延伸20 s,23 個循環,72 ℃徹底延伸60 s。用1%瓊脂凝膠糖電泳鑒定PCR產物,將目的條帶割膠回收,用NdeI和XhoI酶切pET-22b(+)載體,用聚合酶連接酶切后的目的基因和載體片段構建重組表達質粒,轉入BL21(DE3)感受態細胞,在含有卡那霉素的LB平板上篩選陽性質粒。將陽性菌落擴大培養,提取質粒進行XbaI和XhoI酶切鑒定,送至安徽通用生物公司進行測序后用BLAST工具對結果進行序列對比分析。
1.3.2 重組芝麻過敏原Ses i 3蛋白的原核表達
將測序正確的載體轉染BL21(DE3)感受態細胞宿主表達菌中擴大培養,當菌液OD600nm值為0.6時,分別加入異丙基硫代半乳糖苷(isopropyl-β-D-thiogalactoside,IPTG)至終濃度為0.2 mmol/L和1 mmol/L,分別在37 ℃和15 ℃以220 r/min培養4 h。將誘導完成的菌液3 000 r/min離心10 min,收集細菌沉淀,隨后使用超聲破菌緩沖液將沉淀破碎,超聲破碎條件為功率300 W,超聲3 s,停3 s,至菌液澄清透亮,此時為全菌蛋白。將超聲破碎的菌液12 000 r/min離心15 min,上清液即為上清蛋白,沉淀為包涵體蛋白。將包涵體蛋白置于包涵體溶解液中4 ℃攪拌至無沉淀,再將溶解好的溶液置于透析袋中,放入尿素濃度依次為6、4、2、1、0 mol/L的磷酸鹽緩沖液(phosphate buffered saline,PBS)中透析,逐步去除蛋白溶液中的尿素,完成包涵體蛋白的溶解復性,十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)分析其表達情況。
1.3.3 重組芝麻過敏原Ses i 3蛋白的純化及復性
將適量的鎳離子樹脂與溶解復性的包涵體蛋白混合[15],4 ℃混勻2 h后,將充分混合作用的溶液加入純化柱中,用清洗緩沖液洗掉雜蛋白,清洗緩沖液為含有50 mmol/L咪唑和尿素、pH 7.4的PBS。隨后再用洗脫緩沖液洗脫目的蛋白,洗脫緩沖液為含有500 mmol/L咪唑和尿素、pH 7.4的PBS,收集洗脫液,即為純化的重組蛋白。
將純化后蛋白復性至PBS中(含有300 mmol/L NaCl、10%丙三醇溶液,pH 7.4),過濾除菌。復性后蛋白進行SDS-PAGE和Western blot檢測。電泳完成后,去除濃縮膠進行轉印,30 V、45 min。使用1×PBST漂洗5 min共3 次,隨后使用5%脫脂奶粉室溫封閉2 h。經1×PBST漂洗3 次后,使用HRP anti-6X His tag小鼠單克隆抗體(1∶2 000)孵育1 h。最后加入增強型化學發光工作液,靜置2 min后拍照顯色。
1.3.4 芝麻過敏BALB/c小鼠模型
24 只BALB/c 小鼠按體質量隨機分成3 組,分別為PBS 組(對照組),天然Ses i 3(native-Ses i 3,N-Ses i 3)組和重組芝麻蛋白Ses i 3(recombinant Ses i 3,R-Ses i 3)組,每組8 只。N-Ses i 3組與R-Ses i 3組小鼠分別在第0、7、14、21和28天經口灌胃1 mg/只N-Ses i 3和R-Ses i 3(含10 μg CT佐劑),PBS組小鼠則經口灌胃等體積PBS(含10 μg CT佐劑)。第42天,每只小鼠進行5 mg蛋白進行大劑量刺激。采樣,進行相關過敏性指標檢測,所有動物實驗研究均經過揚州大學動物倫理委員會批準(倫理號:YXYLL-2023-091)。
1.3.4.1 過敏癥狀的測定
在大劑量刺激后測定小鼠的體溫變化;同時,觀察各組小鼠的臨床癥狀表現并進行評分,具體如下:0 分=無癥狀;1 分=抓撓鼻子和頭部;2 分=活力降低,活動減少;3 分=眼睛與嘴部周圍浮腫,毛發豎立;4 分=刺激后抽搐或靜止,呼吸困難;5 分=死亡[16]。
1.3.4.2 特異性抗體檢測
對小鼠進行眼部內眥采血,1 200 r/min離心15 min取上清液得到血清。采用間接ELISA方法測定各組小鼠血清特異性Ses i 3的IgE、IgG1和IgG2a的水平[17]。
1.3.4.3 細胞因子及組胺檢測
第42天對小鼠進行眼部內眥采血后置于含乙二胺四乙酸二鉀抗凝劑的離心管中,1 200 r/min離心15 min取上清液得到血漿,用于測定小鼠組胺的含量。IL-4、IL-5、IFN-γ、組胺含量檢測參照ELISA試劑盒說明書。
1.4 統計學方法
2 結果與分析
2.1 重組芝麻過敏原Ses i 3蛋白的原核表達
2.1.1 pET-22b(+)-Ses i 3表達載體的構建及測序
提取芝麻總RNA,檢測其RT-PCR擴增產物,如圖1A所示,在1 500~2 000 bp之間有一條明亮條帶,大小與已公布的Ses i 3大小基本相符。將目的條帶與pET-22b(+)酶切后連接,隨后進行酶切鑒定。由圖1B可知,泳道1中5 000~6 000 bp的條帶為載體,1 500~2 000 bp之間條帶為Ses i 3基因。測序和BLAST工具分析顯示與已公布的Ses i 3基因序列相似度100%,表明Ses i 3基因已被成功克隆且已連接到pET-22b(+)表達載體上。圖1C為pET-22b(+)-Ses i 3表達載體示意圖,蛋白末端為6×His標簽。
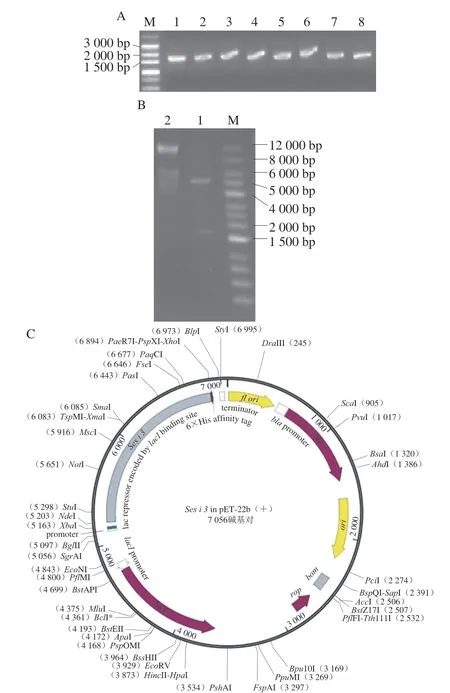
圖1 Ses i 3基因的克隆(A)、pET-22b()-Ses i 3雙酶切鑒定(B)和pET-22b()-Ses i 3圖譜(C)Fig.1 Cloning of Ses i 3 gene (A),identification of pET-22b(+)-Ses i 3 (B) and profile of pET-22b(+)-Ses i 3 (C)
2.1.2 重組芝麻過敏原Ses i 3蛋白的表達及純化
為了探究在不同誘導劑濃度和溫度條件下重組表達載體目的蛋白的表達情況,分別在37 ℃和15 ℃條件下添加不同濃度的IPTG,進行重組蛋白的誘導表達,結果如圖2A所示。泳道1~4表明,在相同溫度下,0.2 mmol/L和1 mmol/L濃度的IPTG均可較好地誘導重組Ses i 3蛋白的表達,且表達量差異不大。鑒于IPTG具有一定的毒性,故選用0.2 mmol/L為IPTG誘導表達的濃度條件。泳道6~13表明,重組芝麻過敏原Ses i 3蛋白以包涵體形式存在,且分子質量約為66 kDa。此外,目的蛋白在37 ℃和15 ℃都是以包涵體的形式存在并且表達量差異不大,因此,選擇大腸桿菌生長常規最適溫度37 ℃作為誘導表達條件。包涵體蛋白溶解透析復性后,用鎳離子吸附柱純化帶有His標簽的重組芝麻過敏原Ses i 3蛋白。由圖2B可見,經過純化后,在66 kDa附近基本沒有出現目的蛋白條帶以外的其他蛋白,因此達到預期純化的效果。包涵體蛋白通過純化后進行復性,圖2C表明,復性的芝麻過敏原Ses i 3蛋白條帶單一,純度較高,無雜蛋白,且復性后表達情況良好。Western blot結果(圖2D)顯示,蛋白與HRP anti-6×His tag小鼠單克隆抗體在大約66 kDa附近有明顯結合,這表明重組Ses i 3蛋白具有一定的活性,可用于后續實驗。
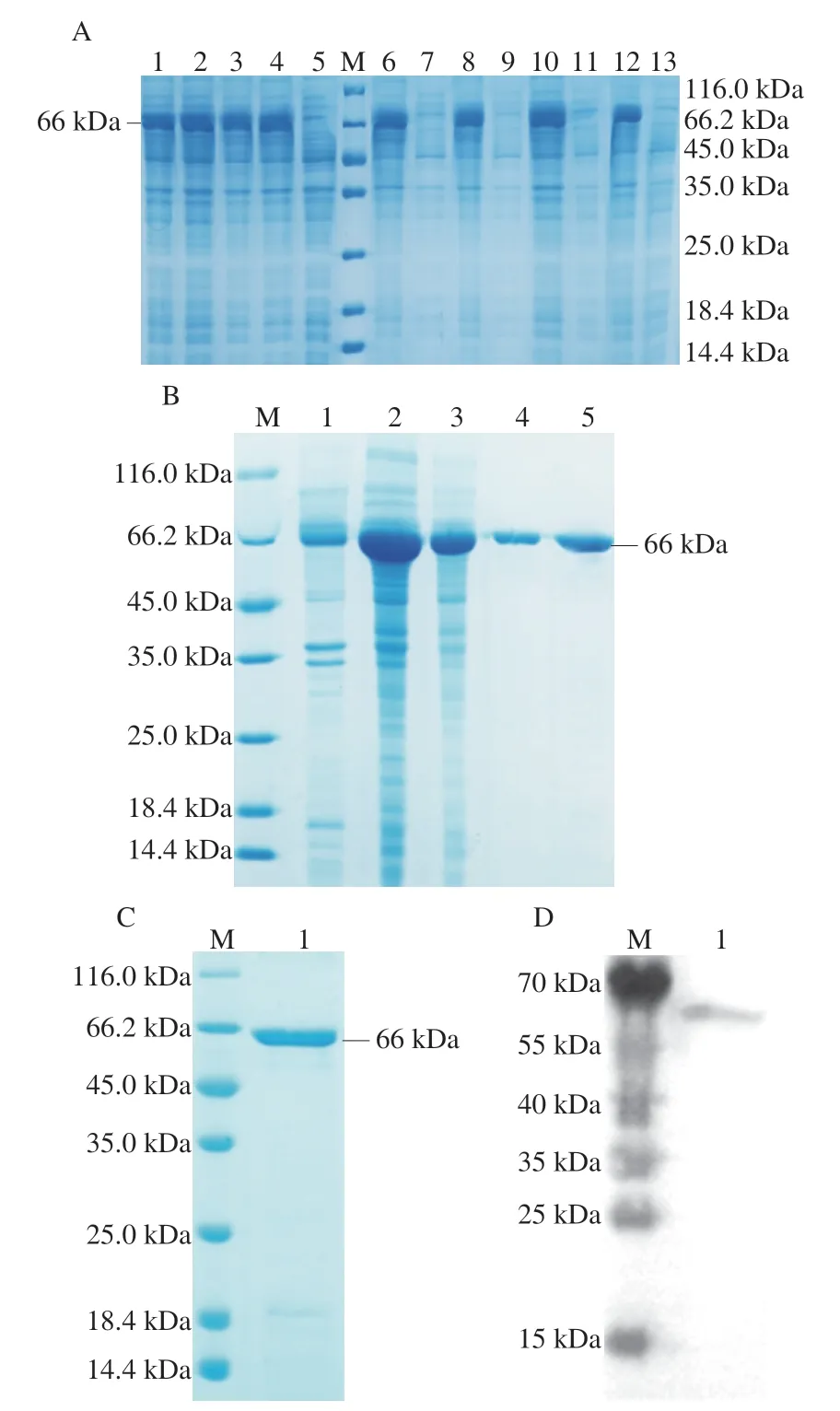
圖2 重組芝麻過敏原Ses i 3蛋白的表達(A)、純化(B)和復性(C)以及Western blot分析(D)Fig.2 Expression (A),purification (B),renaturation (C) and Western blot analysis (D) of recombinant Ses i 3
2.2 芝麻過敏原Ses i 3蛋白的致敏性鑒定
本研究采用BALB/c小鼠模型對R-Ses i 3的致敏性進行評估。如圖3A所示,與PBS組相比,N-Ses i 3和R-Ses i 3組的小鼠經大劑量刺激后,體溫均極顯著下降(P<0.001),R-Ses i 3組的小鼠體溫下降程度略強于N-Ses i 3,但無顯著差異。小鼠過敏癥狀評分如圖3B所示,N-Ses i 3和R-Ses i 3組絕大部分的小鼠表現出較為強烈的過敏癥狀,其中,1 只R-Ses i 3組的小鼠出現刺激后抽搐的癥狀。這表明,N-Ses i 3和R-Ses i 3都引起小鼠產生明顯的食物過敏癥狀。
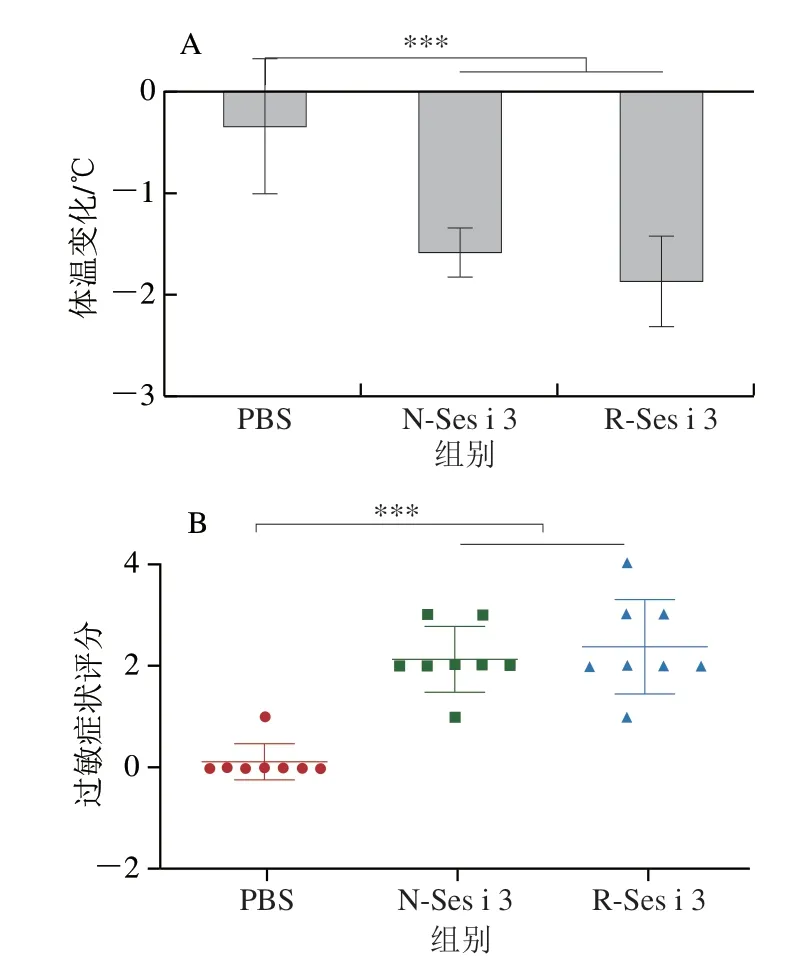
圖3 小鼠體溫變化(A)和過敏癥狀評分(B)Fig.3 Body temperature change (A) and hypersensitivity score (B) of mice
如圖4所示,N-Ses i 3和R-Ses i 3組的小鼠血清中IgE和IgG含量均極顯著高于PBS組(P<0.001),表明N-Ses i 3和R-Ses i 3都具有誘發小鼠芝麻過敏的能力,且在N-Ses i 3和R-Ses i 3組的小鼠血清中IgG1滴度明顯大于IgG2a的滴度,表明在兩組小鼠體內過敏反應均為Th2型。此外,N-Ses i 3和R-Ses i 3組的小鼠血清中特異性抗體的滴度基本相似,但IgG2a有輕微差別。
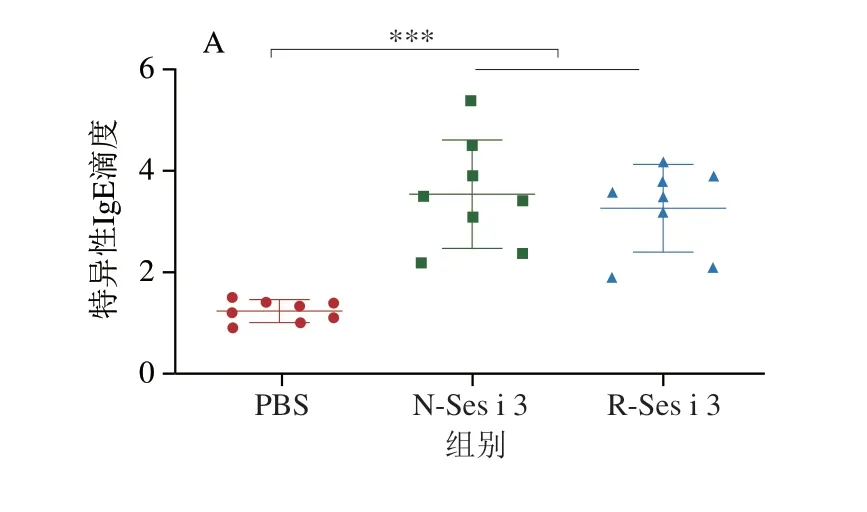
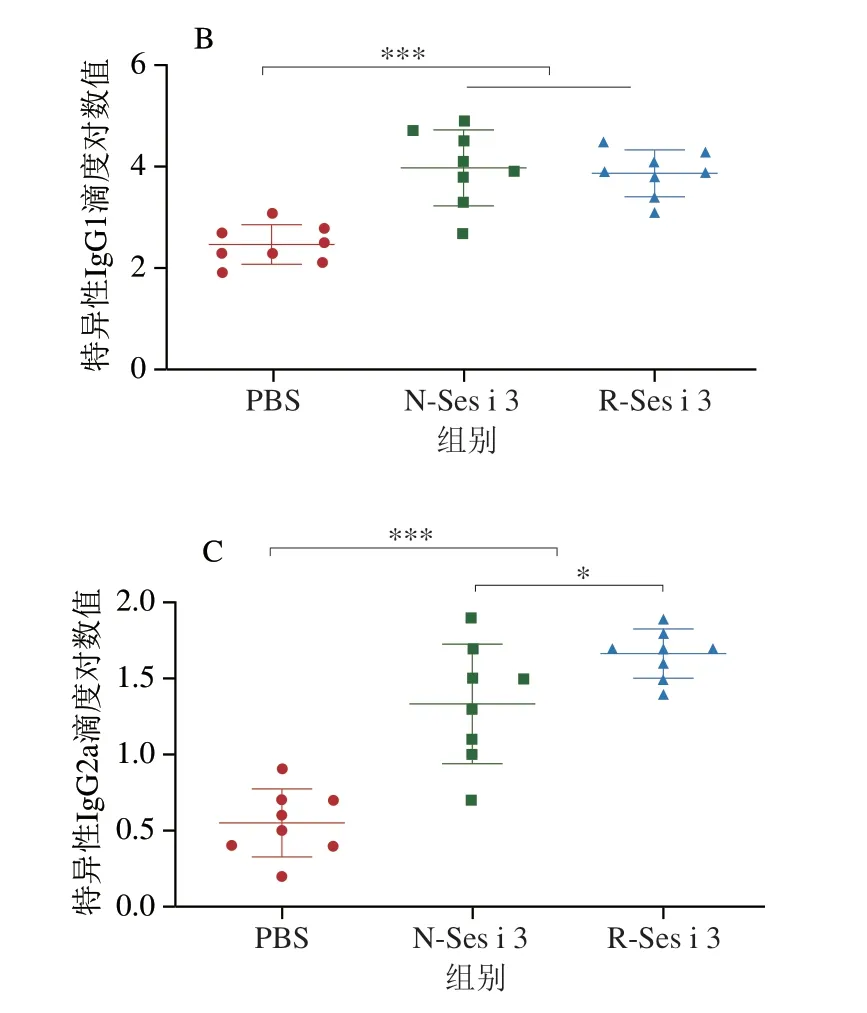
圖4 小鼠血清中特異性IgE(A)、IgG1(B)和IgG2a(C)含量Fig.4 Specific IgE (A),IgG1 (B) and IgG2a (C) contents of mouse serum
如圖5A~C所示,N-Ses i 3和R-Ses i 3組的小鼠血清中Th2型細胞因子IL-4和IL-5均顯著高于PBS組(P<0.05),而Th1型細胞因子IFN-γ與對照組相比無明顯差異,這表明N-Ses i 3和R-Ses i 3都可以通過上調Th2型細胞因子誘發小鼠Th2型過敏反應。此外,N-Ses i 3和R-Ses i 3組小鼠的血漿中組胺含量(圖5D)較PBS組都極顯著升高(P<0.001),這也證明N-Ses i 3和R-Ses i 3誘導小鼠脫顆粒的能力相似。
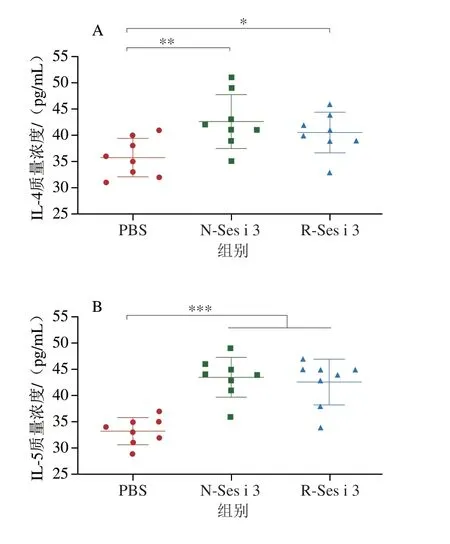
圖5 小鼠血液中的細胞因子和組胺水平Fig.5 Cytokine and histamine levels in mouse serum
3 討論
本研究對芝麻過敏原蛋白Ses i 3進行重組表達并發現其以包涵體形式存在,隨后對包涵體蛋白進行變復性處理,最終得到純度較高的R-Ses i 3。當重組蛋白在細胞中以包涵體形式表達時,雖然純化步驟較可溶性表達繁瑣,但也存在一定的優點,包涵體中的重組蛋白表達較高,變復性后可獲得高純度的蛋白[18-19]。重組表達系統主要有原核表達系統、真核表達系統和無細胞表達系統[20]。而大腸桿菌表達系統作為目前最成熟的表達系統[21],具有培養條件簡單、生長繁殖快、基因操作容易、價格低廉以及快速生產目的蛋白等優點[22],該表達系統在生產花生、魚蝦等多種致敏性較強的食物過敏原重組蛋白中取得較大的進展。如程青麗等[23]利用BL21(DE3)大腸桿菌表達系統,重組牡蠣肌漿鈣結合蛋白,并表明可用重組肌漿鈣結合蛋白替代天然肌漿鈣結合蛋白進行后續致敏性研究。此外,本研究中R-Ses i 3分子質量約為66 kDa,而N-Ses i 3分子質量約為45 kDa,推測可能與pET-22b(+)載體表達的融合標簽及分子間和分子內的靜電斥力有關[24]。本研究進一步采用BALB/c小鼠模型評估R-Ses i 3與N-Ses i 3的致敏性。小鼠過敏模型的建立是評估重組蛋白與天然蛋白致敏性差異的有效方法。史云鳳[25]則采用Rosetta(DE3)大腸桿菌表達系統,成功重組了花生過敏原Ara h 1蛋白,通過建立BALB/c小鼠過敏模型證明重組Ara h 1與天然Ara h 1蛋白具有相似誘發BALB/c小鼠過敏反應的能力。這與本研究中利用BL21(DE3)大腸桿菌表達系統重組表達的R-Ses i 3與N-Ses i 3可以誘發小鼠芝麻過敏反應及脫顆粒的結果一致,可用于后續免疫學相關實驗。
近年來,重組表達的食物過敏原蛋白在食物過敏機制研究及低致敏性食物開發研究上具有良好的應用前景。Sun Lirui等[26]的研究表明,與天然魚類主要過敏原小清蛋白相比,重組小清蛋白具有批量表達靈活、純度高、成本低、周期短等優點,且重組小清蛋白含有天然小清蛋白的大部分結合表位,可用于魚類過敏的診斷及制備低致敏性魚類產品的相關研究。Han Tianjiao等[27]研究表明,重組牡蠣過敏原肌漿鈣結合蛋白具有和天然肌漿鈣結合蛋白相似的理化性質及免疫活性。同時,云肖[28]的研究也表明,重組葡萄牙牡蠣原肌球蛋白與天然葡萄牙牡蠣原肌球蛋白具有相似的結構及免疫學性質。Tian Yang等[29]利用重組Ara h 1蛋白與葡萄糖胺進行糖化和糖基化反應,并證明可有效降低重組Ara h 1蛋白的抗原性,這可能與重組Ara h 1蛋白構象表位的丟失和游離氨基酸殘基修飾其線性表位相關。此外,重組蛋白在定位過敏原蛋白的抗原表位上也具有較大的應用潛力。Fu Linglin等[30]通過重組中國對蝦中的原肌球蛋白和精氨酸激酶,揭示了中國對蝦原肌球蛋白的10 個抗原表位,精氨酸激酶的7 個抗原表位,以及3 個關鍵氨基酸(亮氨酸,天冬氨酸和谷氨酸)。Yang Yang等[31]采用BL21(DE3)大腸桿菌表達系統,重組克氏原螯蝦肌球蛋白輕鏈亞型1蛋白,表明其具有完整的IgE結合活性和結構,可用于進一步的免疫學研究,此外,該研究還利用重組蛋白定義了3 個構象表位區域,并進行空間定位。
4 結論
本研究成功重組表達純化了芝麻過敏原蛋白Ses i 3,并通過動物實驗證實了其與天然Ses i 3蛋白具有相似的免疫學性質,可用于后續芝麻致敏性及致敏機制研究,為未來進一步制備低致敏性芝麻制品奠定了基礎。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
