玉米皮纖維發酵培養黑曲霉和肉原毛平革菌的產酶分析
2023-12-14 12:45:28劉玉春
食品科學 2023年22期
劉玉春,郭 超,王 超
(國家糧食和物資儲備局科學研究院,北京 100037)
玉米皮是玉米深加工的主要副產物之一,通常用作飼料原料[1],而玉米皮中纖維含量約為70%。與軟木、硬木等其他來源纖維相比,玉米皮纖維(corn bran fiber,CBF)具有獨特的單糖組成和結構特征,即半纖維素質量分數達到68%[2],且支鏈化程度高,木質素含量低[3]。目前,已有將玉米皮作為固態發酵原料用于固態發酵產木聚糖酶等的研究報道[4]。
黑曲霉(Aspergillus niger)是國際公認的GRAS(generally regarded as safe)菌株[5-6],被廣泛用于傳統發酵食品和釀造。A.niger具有強大的蛋白質折疊、修飾及胞外蛋白分泌能力[7],已成為發酵工業的重要出發宿主和細胞工廠。A.niger在酶制劑生產方面應用廣泛,如超過30 種以上的食品酶是由A.niger發酵生產,包括纖維素酶、木聚糖酶、淀粉酶、糖化酶、果膠酶等[8-10]。A.niger能夠以糧油加工副產物等生物質為原料發酵產酶[11],而這些生物質原料的成分和結構也會影響A.niger的胞外酶分泌表達。許堯興等[12]報道培養基組成對A.niger產α-半乳糖苷酶的影響。Afangide等[13]報道玉米皮成分對A.niger發酵產木聚糖酶的影響。目前,隨著基因工程、分子生物學等技術應用于A.niger改造,使A.niger成為微生物研究領域的主要課題之一[14-17]。
肉原毛平革菌(Phanerochaete carnosa)是一種白腐真菌,屬于Phanerochaete sensu stricto,最早從香脂冷杉(Abies balsamea)中分離獲得[18]。P.carnosa具有典型的白腐真菌底物偏好性[19],其糖苷水解酶表達活性與黃孢原毛平革菌(P.chrysosporium)相似。但與大多數白腐菌不同,P.carnosa幾乎完全從軟木中分離獲得[20]。P.carnosa除具有完整的木質纖維素降解酶系編碼基因外,其還具有獨特軟木降解酶基因,如錳過氧化物酶、半纖維素酶和糖蛋白降解酶基因[20]。Jurak等[21]研究P.carnosa利用云杉及白楊木培養基生長的轉錄組差異,發現錳過氧化物酶基因(MnP)為該菌株轉錄豐度最高的基因。Mahajan等[22]對P.carnosa在纖維素和云杉底物上生長的胞外蛋白質組進行研究。
發酵底物的成分和結構能夠影響微生物的生長、胞外酶的分泌表達[23]。國內外針對P.carnosa的研究報道很少,且主要集中在P.carnosa對軟木的降解。目前,鮮見以CBF為碳源發酵培養A.niger和P.carnosa,以及對兩種真菌的蛋白質組進行分析的相關報道。本研究以CBF為碳源,應用前期篩選獲得的A.niger和P.carnosa進行發酵培養,分析發酵液中還原糖含量、蛋白質量濃度、木聚糖酶和α-L-阿拉伯呋喃糖苷酶活性變化規律,并應用無標記蛋白質組技術分析與CBF降解相關胞外酶系的分泌表達情況,旨在為以CBF作原料制備具有半纖維素降解優勢的復合酶制劑、酶法水解CBF制備功能性低聚阿拉伯木聚糖提供基礎,促進玉米皮的增值利用。
1 材料與方法
1.1 材料與試劑
玉米皮由河北廣玉淀粉糖業有限公司提供;A.niger和P.carnosa為本實驗室篩選保存的菌株。
木糖、樺木木聚糖 愛爾蘭Megazyme公司;乙酸乙酯 上海阿拉丁生化科技股份有限公司;3,5-二硝基水楊酸、酒石酸鉀鈉 國藥集團化學試劑有限公司;蛋白質濃度測定試劑盒 北京索萊寶科技有限公司;對硝基苯基-α-L-阿拉伯呋喃糖苷(4-nitrophenylα-L-arabinofuranoside,pNPAf)、對硝基苯酚(p-nitrophenol,pNP)美國Sigma-Aldrich公司。
1.2 儀器與設備
MQD-SIR單層小容量振蕩培養箱 上海旻泉儀器有限公司;SynergvHT酶標儀 美國BioTek公司;電泳儀、凝膠成像儀 美國Bio-Rad公司;1200高效液相色譜儀(配紫外檢測器)美國Agilent公司。
1.3 方法
1.3.1 CBF提取和培養基制備
CBF提取參照Lee等[2]方法,玉米皮干燥粉碎后過60 目篩,粉碎后玉米皮與正己烷混合攪拌30 min,攪拌結束后倒掉上層正己烷,重新添加正己烷攪拌,共重復3 次。取100 g脫脂玉米皮,按1∶15(g/mL)加入蒸餾水,再加入耐高溫α-淀粉酶,95 ℃水浴處理1 h;隨后水浴溫度降至55 ℃,加入糖化酶處理2 h,充分脫除淀粉。再加入中性蛋白酶,55 ℃反應1 h,隨后加熱至100 ℃滅酶5 min;紗布過濾,濾渣用蒸餾水洗滌2 次后干燥。
CBF培養基[24]:蛋白胨1.0 g/L、尿素0.2 g/L、CaCl20.3 g/L、MgSO4·H2O 0.3 g/L、(NH4)2SO44.2 g/L、KH2PO42.0 g/L、0.2%(V/V)微量元素(CoCl20.2 g/L、MnSO4·4H2O 1.6 g/L、ZnSO4·7H2O 1.4 g/L、FeSO4·7H2O 5.0 g/L)、10 g/L玉米皮纖維;121 ℃滅菌20 min后備用。
1.3.2A.niger和P.carnosa培養及發酵上清液蛋白分析
A.niger和P.carnosa經PDA平板活化后,接種于200 mL CBF培養基,30 ℃、200 r/min誘導培養。每隔12 h取發酵液樣品,12 000 r/min離心10 min,取上清液,進行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)分析。
1.3.3 理化指標的測定
1.3.3.1 還原糖含量和蛋白質濃度測定
將不同培養時間的發酵上清液適當稀釋,取200 μL樣品,加入300 μL DNS試劑,沸水浴10 min后冷卻,在540 nm波長處測定吸光度。按照試劑盒說明書采用BCA法測定發酵液蛋白質量濃度。
1.3.3.2 木聚糖酶活性測定
將20 μL 適當稀釋發酵上清液加入180 μL 0.5 g/100 mL預熱的樺木木聚糖底物,水浴5 min;再加入300 μL DNS試劑,沸水煮10 min,冷卻至室溫;在540 nm波長處測定吸光度。每分鐘分解底物生成1 μmol還原糖所需的酶量定義為1 個酶活單位(U)[25]。
1.3.3.3α-L-阿拉伯呋喃糖苷酶活性測定
將20 μL發酵液加入180 μL 2 mmol/L pNPAf底物中,40 ℃反應10 min;加入300 μL 1 mol/L Na2CO3溶液終止反應,在405 nm波長處測定吸光度。以不同濃度pNP溶液與1 mol/L Na2CO3溶液混合,繪制標準曲線,根據標準曲線方程計算酶水解底物后pNP的生成量,每分鐘分解底物生成1 μmol的pNP產物所需的酶量定義為1 個酶活力單位(U)[24]。
1.3.4 CBF單糖和酚酸組成測定
1.3.4.1 單糖組成測定
參照1-苯基-3-甲基-5-吡唑酮(1-phenyl-3-methyl-5-pyrazolone,PMP)(甲醇溶解)柱前衍生-高效液相色譜法[26]。稱取20 mg CBF樣品,加入30 mL水,緩慢加入亞鐵氰化鉀溶液和乙酸鋅溶液各5.0 mL,再加水至80 mL,室溫振蕩1 h后離心,濾紙過濾,定容至100 mL。取200 μL樣品加入0.5 mL 4 mol/L三氟乙酸溶液,在120 ℃水解2 h,氮氣吹干;再分別加入0.3 mol/L NaOH溶液和0.5 mol/L PMP各0.5 mL,70 ℃水浴60 min,冷卻至室溫;再加入0.5 mL 0.3 mol/L HCl溶液,隨后加入0.5 mL氯仿,振蕩搖勻后靜置20 min,棄去下層氯仿層,萃取3 次除去多余的PMP衍生劑,上層水相過膜,進行高效液相色譜分析。混合標準品由葡萄糖、木糖、阿拉伯糖、甘露糖、鼠李糖、葡萄糖醛酸、半乳糖醛酸、半乳糖、巖藻糖和核糖10 種單糖和乳糖內標(均為20 mmol/L)組成。
高效液相色譜參數:色譜柱:SHISEIDO C18柱(4.6 mm×250 mm,5 μm);流動相:0.1 mol/L磷酸鹽緩沖溶液(pH 6.9)-乙腈,體積比為82∶18;檢測波長245 nm;柱溫25 ℃,流速1.0 mL/s。進樣量10 μL。
1.3.4.2 酚酸含量測定
參考Sani 等[27]方法并進行適當調整。稱取200 mg CBF,加入10 mL體積分數80%乙醇溶液渦旋混勻,超聲處理30 min,8 000 r/min離心10 min;取上清液,35 ℃旋轉蒸發除乙醇,殘渣用20 mL純水溶解,轉移至50 mL容器中,用氫氧化鉀溶液調節pH值至7.0,再加入20 mL乙酸乙酯振蕩10 min后取上層酯層,水相再用20 mL乙酸乙酯重新萃取2 次,合并乙酸乙酯相為中性酚;調節水相pH值至2.0,再用20 mL乙酸乙酯萃取3 次,合并乙酸乙酯層為酸性酚,將中性酚和酸性酚合并后35 ℃濃縮至干,殘渣用體積分數60%甲醇溶液定容至10 mL,過0.22 μm濾膜后待測。
高效液相色譜參數:色譜柱:Agilent C18柱(4.6 mm×150 mm,5 μm);流動相A:體積分數1%乙酸溶液;流動相B:乙腈;柱溫35 ℃,流速為1.0 mL/s;檢測波長270 nm;進樣量10 μL。梯度洗脫條件:0~5 min,94%~86% A、6%~14% B;5~8 min,86%~82% A、14%~18% B;8~15 min,82%~80% A、18%~20% B;15~20 min,80%~76% A、20%~24% B;20~30 min,保持96% A、4% B。
1.3.5 無標記蛋白質組學分析
取培養60 h的發酵液,12 000 r/min離心10 min,分離上清液,發酵液蛋白樣品進行酶解處理[28]。蛋白質譜鑒定實驗委托北京諾禾致源科技股份有限公司進行。蛋白質生物信息學使用InterProScan軟件進行基因本體論(Gene Ontology,GO)和結構域IPR功能注釋(包括Pfam、PRINTS、ProDom、ProSite數據庫)。使用ExPASy中的Compute pI/Mw工具評估蛋白質的分子質量(mw)和理論等電點(pI)。
1.4 數據處理與分析
采用GraphPad Prism 5進行處理數據;采用Adobe Illustrator軟件處理圖片。
2 結果與分析
2.1 CBF的單糖和酚酸組成
由表1可知,CBF樣品中含有10 種單糖,木糖、阿拉伯糖、半乳糖、葡萄糖、半乳糖醛酸、葡萄糖醛酸、巖藻糖、甘露糖、鼠李糖、核糖的物質的量比為720.4∶402.5∶130.4∶76.5∶24.6∶20.2∶15.7∶10.8∶10.2∶1。其中木糖(50.97%)和阿拉伯糖(28.47%)的含量最高,阿拉伯糖/木糖為0.56(表1),該結果與已報道研究結果一致,即CBF單糖主要包括阿拉伯糖(29%~36%),半乳糖(4%~6%),葡萄糖(0%~2%),木糖(55%~59%)和葡萄糖醛酸(3%~5%)[29-30]。CBF阿魏酸含量為0.09%,與已報道相比含量較低[31],香豆酸和咖啡酸含量低于檢測限。研究顯示,木質纖維素生物質的主要成分為纖維素(40%~45%)、半纖維素(25%~30%)和木質素(15%~20%)[31],CBF成分結構與一般木質素纖維明顯不同,可作為底物用于發酵制備具有半纖維素降解優勢的復合酶制劑。

表1 CBF的單糖和酚酸組分構成Table 1 Compositions of monosaccharides and phenolic acids in CBF
2.2 A.niger和P.carnosa在CBF培養基中的生長及發酵上清液蛋白分析結果
A.niger和P.carnosa在CBF培養基中均能夠良好生長,發酵培養192 h后,能夠觀察到球狀菌體,發酵液狀態變為澄清(圖1)。

圖1 發酵培養192 h后的CBF培養基Fig.1 CBF-containing medium after 192 h fermentation
圖2為A.niger和P.carnosa發酵過程中不同時間發酵液SDS-PAGE檢測結果。發酵液中蛋白條帶隨發酵培養時間延長呈現明顯變化,其中A.niger在第48~60小時變化最為明顯,而P.carnosa在第36~48小時變化最為明顯。


圖2 不同培養時間A.niger(A)和P.carnosa(B)發酵上清液SDS-PAGE圖譜Fig.2 SDS-PAGE patterns of total extracellular proteins in the fermentation supernatants of A.niger (A) and P.carnosa (B)
2.3 A.niger和P.carnosa發酵液還原糖和蛋白含量
由圖3A可知,A.niger和P.carnosa發酵液中蛋白質量濃度最高分別為1.21 mg/mL(192 h)和1.23 mg/mL(192 h)。發酵培養過程中,A.niger和P.carnosa發酵液中蛋白含量逐步上升,A.niger發酵液蛋白質量濃度由12 h的0.70 mg/mL升高至192 h的1.21 mg/mL;其中在48~60 h出現第1個較快升高的階段,該結果與SDSPAGE檢測結果(圖2A)一致。P.carnosa發酵液蛋白質量濃度由12 h的0.72 mg/mL升高至192 h的1.23 mg/mL,而在36~48 h出現一個迅速升高的階段,即由0.82 mg/mL升高至1.05 mg/mL,該結果與SDS-PAGE檢測結果(圖2B)一致,推測該階段P.carnosa可能快速分泌表達胞外酶以降解CBF。
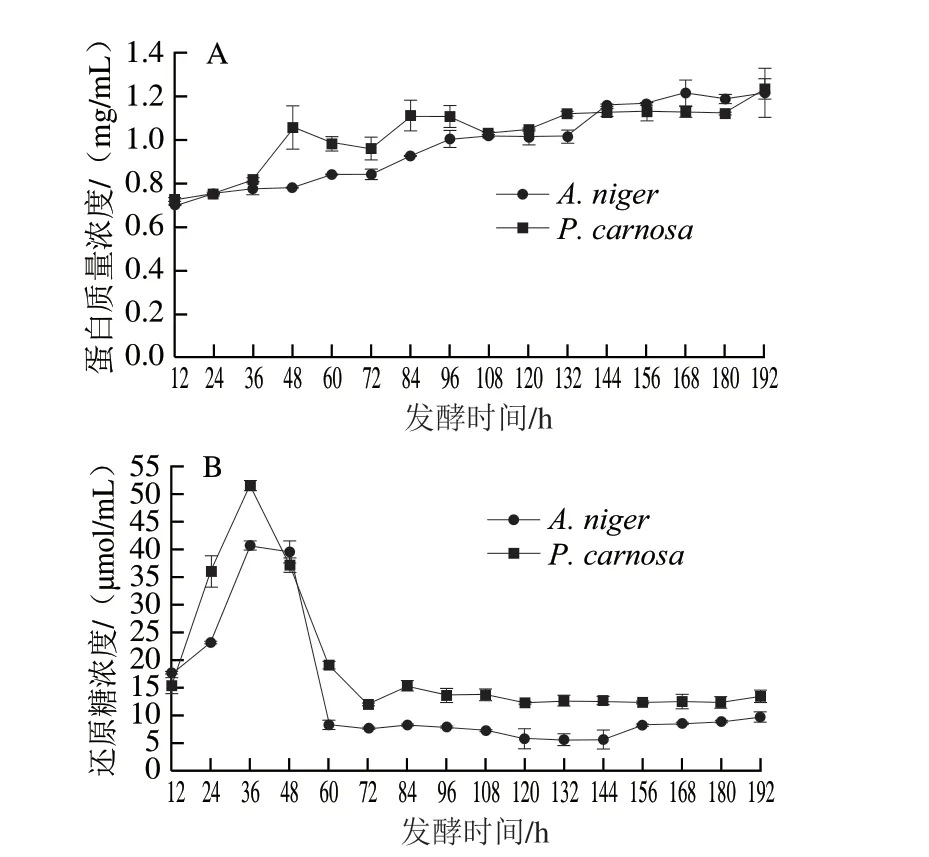
圖3 A.niger和P.carnosa發酵液中蛋白質量濃度(A)和還原糖濃度(B)變化Fig.3 Changes in extracellular protein (A) and reducing sugar concentration (B) in the fermentation broths of A.niger and P.carnosa
由圖3B可知,A.niger和P.carnosa發酵液中還原糖濃度最高為40.52 μmol/L和51.37 μmol/mL,均出現在36 h。A.niger和P.carnosa發酵過程中還原糖濃度變化趨勢相似。A.niger在12~36 h迅速升高,在36 h達到最高為40.52 μmol/mL,在36~60 h又迅速降低至8.18 μmol/mL,60~192 h在5.55~9.61 μmol/mL之間呈較低水平波動。P.carnosa在36 h達到最高(51.37 μmol/mL),之后在36~72 h迅速降低至11.91 μmol/mL,72~192 h在11.91~15.23 μmol/mL之間波動。
2.4 A.niger和P.carnosa發酵液中木聚糖酶和α-L-阿拉伯呋喃糖苷酶活性
由圖4A可知,A.niger和P.carnosa發酵液中木聚糖酶活力最高分別為246.58 U/mL(72 h)和367.21 U/mL(60 h)。A.niger發酵過程中木聚糖酶活力在36~72 h迅速升高,72 h達到最高,72~120 h緩慢下降至163.09 U/mL,而120~156 h又緩慢上升至208.22 U/mL。P.carnosa發酵過程中木聚糖酶活力在24~60 h迅速升高,60~84 h緩慢下降至317.86 U/mL,而84~192 h在312.15~350.16 U/mL間波動。P.carnosa與A.niger發酵液中木聚糖酶活性波動相似,但P.carnosa在各階段木聚糖酶活性均高于A.niger,是其酶活力的1.38~4.76 倍。A.niger發酵液木聚糖酶活性在48~60 h升高最快,而P.carnosa發酵液木聚糖酶活性在36~48 h升高最快,均與發酵液蛋白質量濃度變化呈正相關(圖3),推測兩株菌在此階段開始大量分泌表達木聚糖酶。
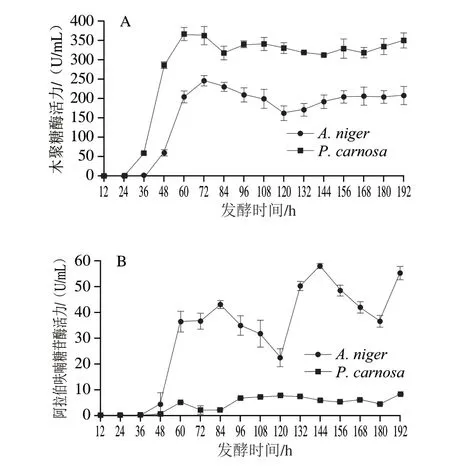
圖4 A.niger和P.carnosa發酵液中木聚糖酶(A)和α-L-阿拉伯呋喃糖苷酶(B)活力變化Fig.4 Changes in xylanase (A) and arabifurosidase (B) activities in the fermentation broths of A.niger and P.carnos
由圖4B可知,A.niger和P.carnosa發酵過程中α-L-阿拉伯呋喃糖苷酶活性變化趨勢差異很大。A.niger和P.carnosa發酵液中α-L-阿拉伯呋喃糖苷酶活力最高分別為57.90 U/mL(144 h)和8.26 U/mL(192 h)。A.niger發酵液α-L-阿拉伯呋喃糖苷酶活性變化很大,在36~84 h迅速升高至42.99 U/mL,84~120 h迅速下降(42.99~22.39 U/mL),形成第1個峰;隨后由120~144 h迅速升高(22.39~57.90 U/mL),144~180 h迅速下降(57.90~36.49 U/mL),形成第2個峰;最后在180~192 h又迅速升高至55.13 U/mL。P.carnosa發酵液α-L-阿拉伯呋喃糖苷酶活性同樣呈波動變化,但其酶活力相對于A.niger發酵液較低,最高僅為8.26 U/mL。發酵過程中發酵液中α-L-阿拉伯呋喃糖苷酶活性波動性變化趨勢可能有兩個原因:首先,阿拉伯糖殘基作為CBF支鏈,其與主鏈存在多種連接方式,而不同α-L-阿拉伯呋喃糖苷酶具有不同催化模式[32-33],因此,推測A.niger可能在不同發酵階段表達具有不同催化模式的α-L-阿拉伯呋喃糖苷酶;其次,CBF的降解過程是α-L-阿拉伯呋喃糖苷酶與木聚糖酶協同作用的過程,因此,兩種真菌可能在生長過程中調控相關α-L-阿拉伯呋喃糖苷酶與木聚糖酶基因的協同表達,結果則表現為發酵液中酶活力的波動現象。
綜合發酵液中還原糖含量和蛋白質量濃度變化,以及木聚糖酶和α-L-阿拉伯呋喃糖苷酶活性變化規律,兩種菌在發酵過程中均存在“分泌胞外酶-酶解CBF-產生低聚糖底物-吸收利用-生長-再分泌胞外酶”的波動現象,而這種通常出現在非連續補料發酵培養過程。
2.5 蛋白質的鑒定及統計分析
無標記蛋白質組分析P.carnosa發酵液共鑒定出109 種蛋白質,包括糖苷水解酶(53/48.62%)、蛋白酶/肽酶(9/8.26%)、脂肪酶/酯酶(14/12.84%)、氧化酶/還原酶(6/5.50%)和假設蛋白(27/24.77%)(圖5、表2)。CBF發酵培養P.carnosa分泌表達的糖苷水解酶比列高于結晶纖維素(46%)和云杉木(29%)培養,而氧化酶/還原酶分泌表達比例(5%)低于結晶纖維素(15%)和云杉木(15%)[22]誘導表達比例(表2),該結果表明CBF與結晶纖維素、云杉木的組成成分和分子結構的差異導致同種菌株會分泌表達具有不同組成的復合酶系。

圖5 CBF發酵培養A.niger(A)和P.carnosa(B)分泌表達蛋白質的功能分類Fig.5 Functional classification of proteins identified in the secretomes of A.niger (A) and P.carnosa (B) grown in CBF-containing medium

表2 不同底物發酵培養A.niger和P.carnosa分泌表達蛋白質的功能分類比較Table 2 Functional classification of proteins identified in the secretomes of A.niger and P.carnosa grown in different media %
A.niger發酵液蛋白質組分析共鑒定出105 種蛋白質,包括糖苷水解酶(82/78.10%)、蛋白酶/肽酶(9/8.57%)、脂肪酶/酯酶(9/8.57%)、氧化酶/還原酶(2/1.9%)和植酸酶(3/2.86%)(圖5、表2)。CBF發酵培養A.niger分泌表達的糖苷水解酶比列高于甘蔗渣(72.28%)和木薯渣(69.57%),氧化酶/還原酶(1.90%)低于甘蔗渣(3.96%)和木薯渣(3.26%)[35]誘導表達比例(表2)。該結果與P.carnosa蛋白質組分析結果一致,表明A.niger分泌表達胞外酶系與發酵底物組成成分和結構相關。
蛋白組分析結果顯示,P.carnosa分泌表達了完整的纖維素降解酶系(表3):10 個內切葡萄糖苷酶(GH5、GH12、GH16、GH45和GH74家族)、4 個纖維二糖酶(GH6和GH7家族)、1 個β-1,3-葡聚糖酶(GH17家族)和5 個β-葡萄糖苷酶(GH3、GH31和GH55家族)。研究顯示,白腐菌降解纖維素需要內切葡聚糖酶、纖維二糖酶和β-葡萄糖苷酶3 種酶協同作用[34],而CBF發酵培養P.carnosa的蛋白質組中能夠檢測到3 類纖維素降解酶。
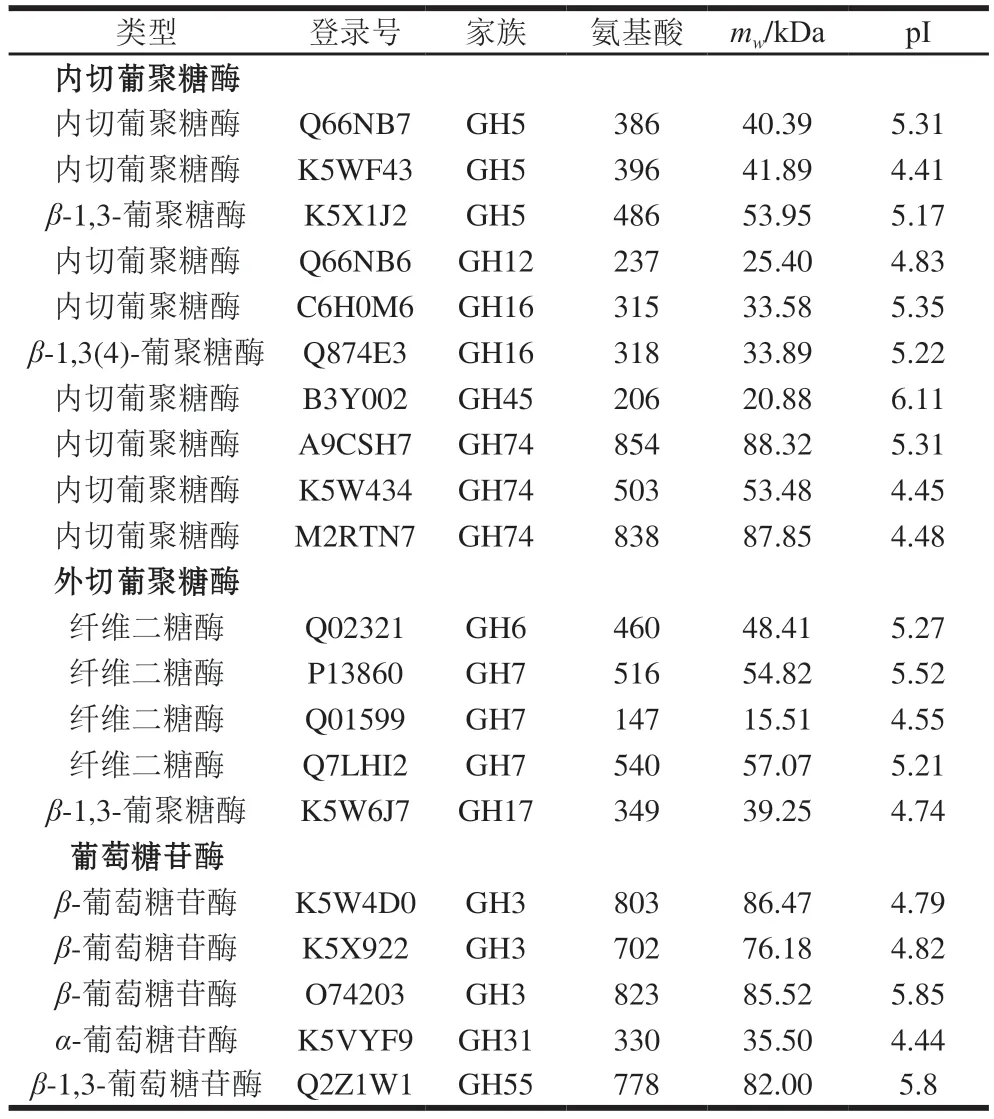
表3 CBF發酵培養P.carnosa分泌表達的纖維素降解酶類Table 3 Cellulose-degrading proteins identified in the secretome of P.carnosa cultured on CBF
P.carnosa能夠分泌表達完整的半纖維素降解酶系(表4,圖6),包括4 個木聚糖酶(GH10和GH11家族)、1 個β-木糖苷酶(GH3家族)、4 個α-L-阿拉伯呋喃糖苷酶(GH43和GH127家族)、3 個乙酰木聚糖酯酶(CE1家族)和1 個葡萄糖醛酸酶(GH115家族)、6 個半乳糖苷酶(GH27、GH35和GH43家族)、2 個甘露聚糖酶(GH5家族)和3 個甘露糖苷酶(GH5、GH47和GH92家族)。
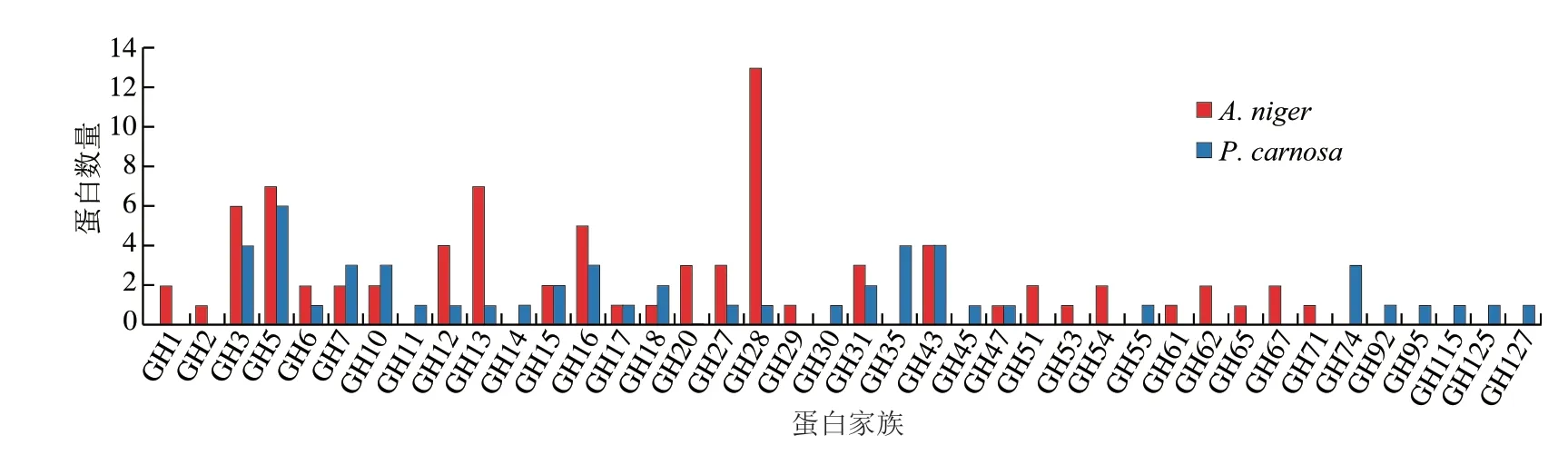
圖6 A.niger和P.carnosa分泌表達糖苷水解酶家族蛋白分布Fig.6 Distribution of glycosyl hydrolase family identified in the secretome of A.niger and P.carnosa grown in CBF-containing medium
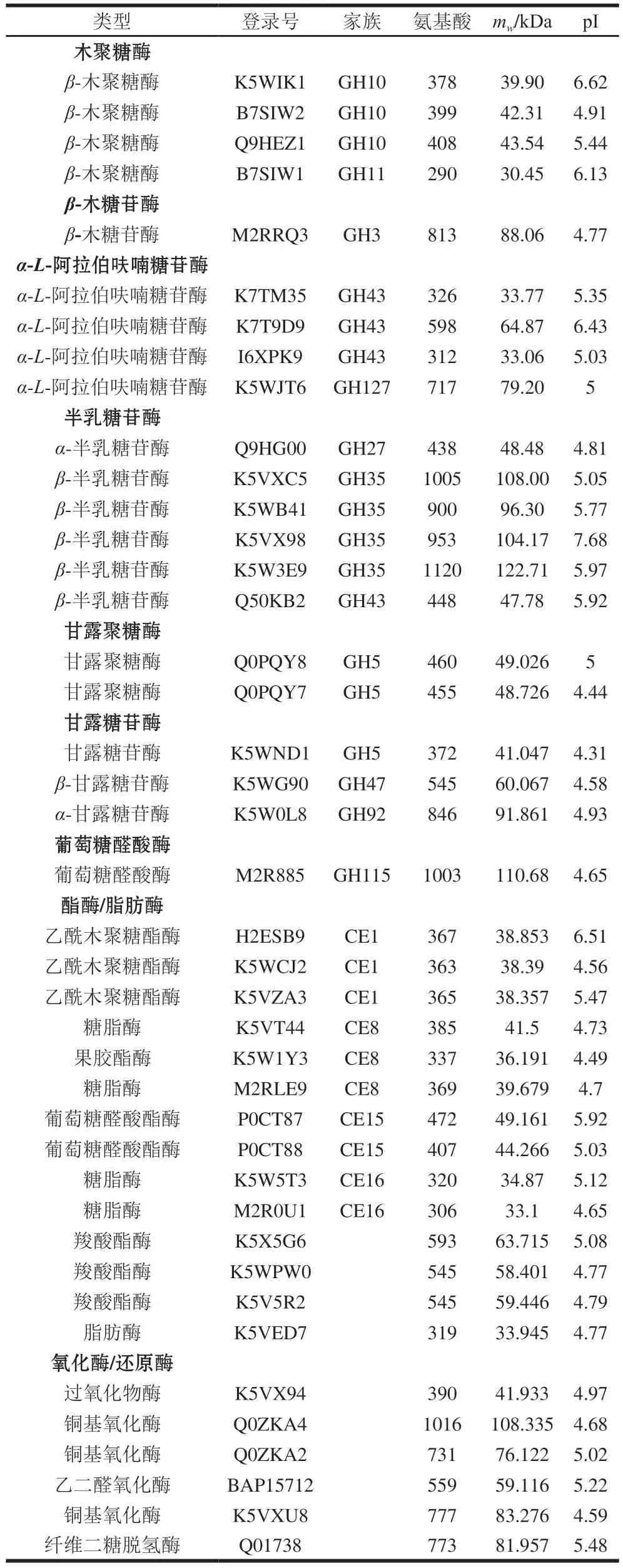
表4 CBF發酵培養P.carnosa分泌表達的半纖維素降解酶類Table 4 Hemicellulose-degrading enzymes identified in the secretome of P.carnosa cultured on CBF
P.carnosa蛋白質組鑒定到6 個氧化/還原酶,包括過氧化物酶、銅基氧化酶、乙二醛氧化酶、纖維二糖脫氫酶(表4),但未檢測到錳過氧化物酶和細胞色素P450單加氧酶。有報道顯示,P.carnosa基因組含有266 個預測為細胞色素P450單加氧酶的編碼基因[20]。在已測序和注釋的木腐擔子菌中,P.carnosa的P450單加氧酶基因數量最多,幾乎是P.chrysosporium的2 倍[20],但本研究顯示,P.carnosa未分泌表達細胞色素P450單加氧酶。該結果與P.carnosa利用云杉及白楊木培養基生長的轉錄組和蛋白質組分析結果[21-22]不同,可能由于CBF與云杉及白楊木的木質素含量和結構存在巨大差異,CBF的木質素含量約為7.8%[36]。纖維素和云杉發酵培養P.carnosa的胞外蛋白質組中能夠檢測到錳過氧化物酶,但不能檢測到纖維二糖脫氫酶的表達[22]。此外,云杉發酵培養P.carnosa還能夠檢測到過氧化物酶和銅基氧化酶[22]。
P.carnosa分泌表達了14 個脂肪酶/酯酶(CE1、CE8、CE15和CE16家族),包括果膠酯酶、糖脂酶、乙酰木聚糖脂酶、羧酸酯酶、葡萄糖醛酸酯酶和脂肪酶(表4),但沒有檢測到阿魏酸酯酶的表達。脂肪酶和酯酶底物特異性廣泛,能夠參與水解半纖維素、纖維素的糖酯類支鏈結構酯鍵,如葡萄糖醛酸酯酶可以水解木質素和木聚糖葡萄糖醛酸支鏈之間的酯鍵[37]。P.carnosa分泌表達了9 個蛋白酶/肽酶,這些蛋白酶可能參與纖維素結合的蛋白水解。
P.carnosa發酵液蛋白質組中檢測到27 個功能未知的假設蛋白(圖5),該結果與前期報道相似[22]。P.carnosa表達較多假設蛋白可能是由于國內外對于該菌種研究報道很少,對該菌株分泌表達蛋白質功能研究不足,但這些蛋白可能在木質纖維素降解中發揮重要作用[38]。因此,蛋白質組分析也是尋找真菌在特定培養條件下表達相關的未知功能蛋白質的重要途徑。
蛋白組分析顯示,A.niger分泌表達完整的纖維素降解酶系(表5),包括12 個內切葡聚糖酶(GH5、GH12、GH16、GH61和GH71家族)、4 個纖維二糖酶(GH6和GH7家族)和8 個β-葡萄糖苷酶(GH1、GH3、GH17和GH31家族)。
A.niger也分泌表達了完整的半纖維素降解酶系(表6),包括2 個木聚糖酶(GH10家族)、2 個木糖苷酶(GH3和GH31家族)、9 個α-阿拉伯呋喃糖苷酶(GH43、GH51、GH54和GH62家族)、1 個乙酰木聚糖酯酶(CE1家族)、1 個阿魏酸酯酶和2 個α-葡萄糖醛酸苷酶(GH67家族)、4 個半乳糖苷酶(GH3和GH27家族)、1 個甘露聚糖酶(GH5家族)、3 個甘露糖苷酶(GH2、GH5和GH47家族)。
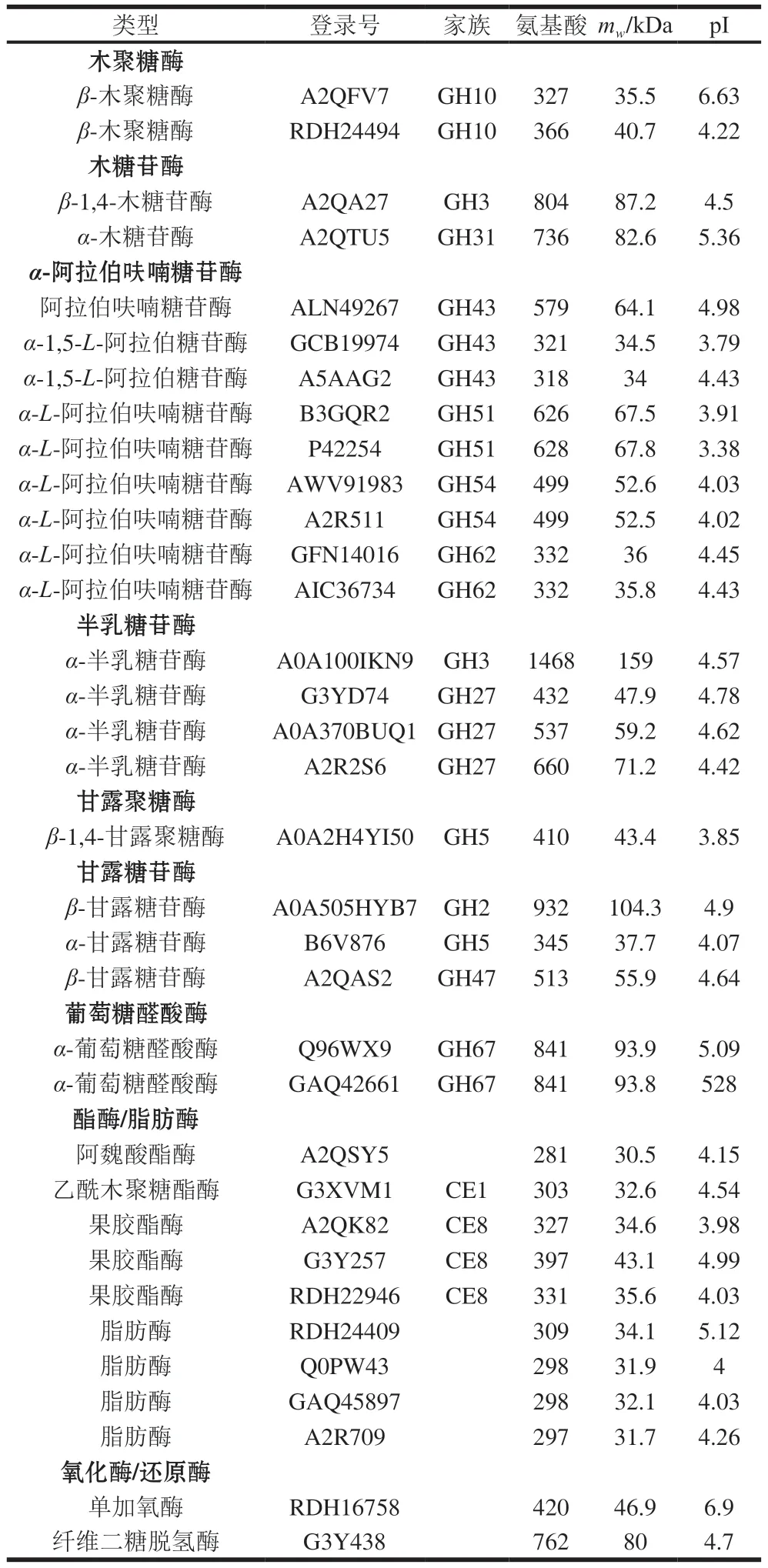
表6 CBF發酵培養A.niger分泌表達的半纖維素降解酶類Table 6 Hemicellulose-degrading enzymes identified in the secretome of A.niger cultured on CBF
A.niger蛋白質組鑒定到2 個氧化酶/還原酶,分別為單加氧酶和纖維二糖脫氫酶。A.niger分泌表達了9 個脂肪酶/酯酶(CE1和CE8家族),包括乙酰木聚糖脂酶、阿魏酸酯酶和果膠酯酶,均為半纖維素主要支鏈降解酶。A.niger分泌表達了9 個蛋白酶/肽酶和3 個植酸酶。與P.carnosa蛋白質組分析結果不同,A.niger發酵液蛋白質組中未檢測到假設蛋白。
如圖6所示,A.niger分泌表達糖苷水解酶的數量遠高于P.carnosa,且糖苷水解酶家族分布也存在很大差異。該結果與已有研究一致,即A.niger具有強大的淀粉糖和非淀粉多糖降解酶系,在分泌表達糖苷水解方面具有優勢[9]。P.carnosa分泌表達氧化酶/還原酶數量高于A.niger(圖5),其在水解利用木質素方面有優勢。盡管兩種真菌具有不同的胞外酶系表達特點,但兩種真菌都能夠利用CBF作為碳源生長。綜上所述,兩種真菌在降解CBF時采取不同協同策略,即分泌表達具有不同組成的胞外酶系,但又具有相似催化功能和效率。
Label-free技術分析揭示了兩種真菌發酵降解CBF所分泌表達的降解酶系,其中也包括功能未知的假設蛋白,這為新的糖苷水解酶的鑒定和發掘提供了方向。此外,蛋白質組分析也揭示了發酵底物組成、結構與微生物分泌表達胞外酶的相關性,該結果可以為設計制備最適合降解CBF的復合酶制劑提供基礎。
3 結論
CBF中木糖(50.97%)和阿拉伯糖(28.47%)的含量最高,阿拉伯糖/木糖為0.56。以CBF為碳源發酵培養A.niger和P.carnosa,并測定發酵液中還原糖濃度、蛋白質量濃度、木聚糖酶和α-L-阿拉伯呋喃糖苷酶活性變化規律。結果顯示,A.niger和P.carnosa發酵過程中均存在“分泌胞外酶-酶解CBF-產生低聚糖底物-吸收利用-生長-再分泌胞外酶”的波動現象。Label-free技術分析顯示,A.niger分泌表達糖苷水解酶的數量遠高于P.carnosa,且糖苷水解酶的家族分布也存在很大差異;P.carnosa分泌表達氧化酶/還原酶數量高于A.niger。兩種真菌都能夠利用CBF作為碳源生長,但分泌表達的不同復合酶又具有相似催化功能和效率,顯示兩種真菌在降解CBF時采取不同的多酶協同策略。Label-free蛋白質組分析揭示了兩種真菌發酵降解CBF所分泌表達的降解酶系,以及發酵底物組成、結構與微生物發酵分泌酶蛋白系的相關性,該結果可為設計制備最適合降解CBF的復合酶制劑提供一定基礎。
