乳源N-聚糖的結構與功能研究進展
2023-12-14 08:26:24彭秋琦于景華馬曉明劉雷
中國乳品工業 2023年11期
關鍵詞:結構
彭秋琦,于景華,馬曉明,劉雷
(1.天津科技大學食品科學與工程學院 天津 300457;2.宜品乳業(青島)集團有限公司 山東 青島 266555)
0 引 言
母乳是嬰兒最佳的營養食品,能夠為嬰兒提供蛋白質、脂肪、碳水化合物、礦物質及維生素等營養成分,也可提供多種生物活性成分[1],如免疫球蛋白、骨橋蛋白、生長因子、低聚糖等[2]。近年來,乳源低聚糖作為乳中的第三大營養成分,因其豐富的生物學功能引起了人們的注意,模擬母乳低聚糖成分的嬰兒配方奶粉應運而生。目前大多數的研究都集中在游離低聚糖上,而與乳蛋白質結合的聚糖的相關研究較少,如修飾糖蛋白的N-聚糖[3]。N-聚糖的合成源于蛋白質的N-糖基化。糖基化是乳蛋白質最普遍和最復雜的翻譯后修飾之一,是一個非模板驅動的生物合成過程。乳腺上皮作為乳腺的功能單位,是乳蛋白合成和分泌的主要部位[4]。在乳腺中,蛋白質N-糖基化沿著內質網-高爾基體分泌途徑進行。在內質網中,寡糖轉移酶將與多萜醇相連的N-聚糖前體轉移到新生多肽(NX-S/T)的共識序列中的天冬酰胺的酰胺氮殘基上,在高爾基體中,N-聚糖前體被不同的糖苷酶和糖基轉移酶修剪和拉長,從而介導了N-聚糖的成熟[5]。在蛋白質糖基化的過程中,N-糖鏈分子賦予了蛋白質分子多種多樣的生物學功能。因此,為了更好地理解乳源N-聚糖在嬰兒健康中如何發揮作用,本文綜述了乳糖蛋白中N-聚糖的結構特性及不同物種間含量與結構的比較(人乳、牛乳和山羊乳),對乳源N-聚糖的生物學功能及其在嬰兒配方奶粉中的應用及前景加以闡述。
1 乳源N-聚糖的結構
1.1 乳源N-聚糖的組成結構特性
糖基化是乳蛋白質中最重要、最常見的翻譯后修飾類型之一,在糖基轉移酶和糖苷酶的共同調控下,糖鏈通過共價結合的方式連接到蛋白質的特定位點,形成糖蛋白。糖基化通過穩定蛋白質的三維結構,促進細胞間的識別和細胞信號的傳遞,這其中修飾蛋白質的糖鏈發揮了巨大作用[6-7]。據統計,70%的人乳蛋白質均被糖鏈修飾[8],如乳鐵蛋白、分泌型免疫球蛋白(sIgA)、部分脂肪球膜蛋白、κ-酪蛋白等。根據糖鏈在蛋白質上的連接位置不同,大致可分為N-糖鏈和O-糖鏈。N-糖鏈連接在蛋白質肽鏈中天冬酰胺殘基側鏈的酰胺氮上,一般由甘露糖(Man)、N-乙酰氨基葡萄糖(GlcNAc)、半乳糖(Gal)、巖藻糖(Fuc)及唾液酸(N-乙酰神經氨酸Neu5Ac 和N-羥乙酰神經氨酸Neu5Gc)組成。所有的N-聚糖都具有統一的核心結構:五糖核心,即2 個N-乙酰氨基葡萄糖連接3 個甘露糖。根據N-聚糖的分泌途徑和五糖核心外圍分支的不同,N-聚糖可分為高甘露糖型(High mannose)、雜合型(Hybrid)和復合型(Complex)[9],如圖1所示。
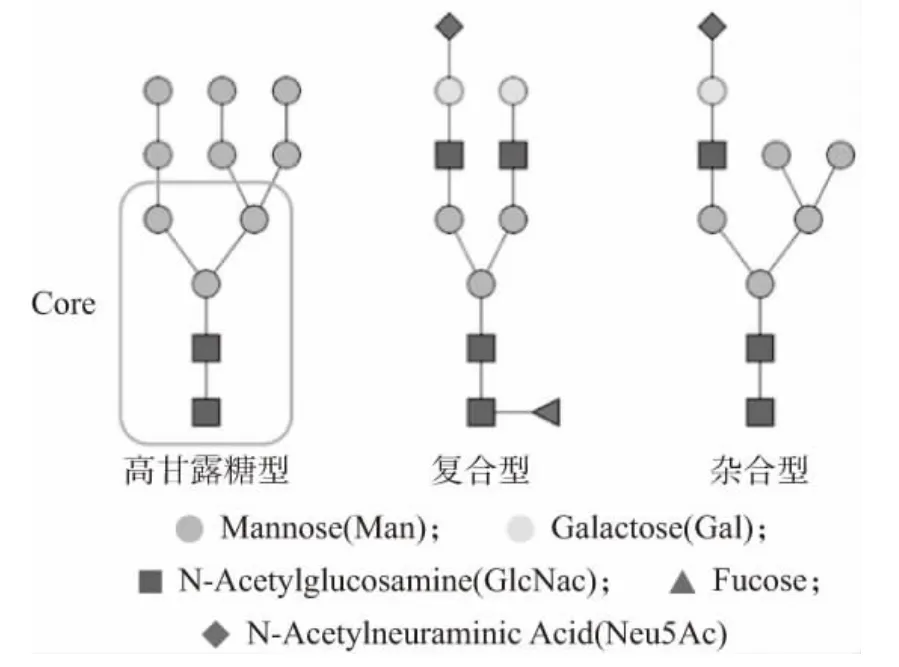
圖1 乳源N-聚糖的3 種主要類型
在核心結構的基礎上,N-聚糖可以被巖藻糖修飾。母乳N-聚糖的巖藻糖基化修飾依賴于編碼巖藻糖基化轉移酶的基因[10]。到目前為止,在人類基因組中已檢測到13 種不同的巖藻糖基轉移酶(FUTs)[11]。FUT2 和FUT3 這2 種基因決定了母親的分泌型狀態(Se)和Lewis 血型抗原(Le),根據FUT2 和FUT3 的活性表達,可將母親群體分為4 組:分泌型Lewis 陽性(Se+Le+)、分泌型Lewis 陰性(Se+Le-)、非分泌型Lewis陽性(Se-Le+)和非分泌型Lewis 陰性(Se-Le-)[12]。不同的基因編碼不同的巖藻糖基轉移酶。FUT8 基因通過編碼α1,6-巖藻糖基轉移酶,將Fuc 殘基轉移到N-糖鏈最內側的GlcNAc 單元;FUT2 Se 基因編碼α1,2-巖藻糖基轉移酶;FUT3 或其他α1,3-基因(FUT4,5,6,7,9)編碼α1,3 或α1,4-巖藻糖基轉移酶[13]。除巖藻糖修飾外,N-聚糖還可以被唾液酸修飾。唾液酸作為糖蛋白N-糖鏈上重要的末端修飾單糖,可通過α-2,3 或α-2,6 鍵連接在端基半乳糖上,構成不同鍵型的唾液酸化N-糖鏈。然而,對于作用在α-2,3 或α-2,6 鍵上的酶的作用機制尚未可知,仍需進一步探究。以上2 種重要的糖基化修飾在嬰兒的生長發育中起著重要作用[14]。
1.2 不同乳源N-聚糖的結構與含量比較
乳源N-聚糖的含量與結構受物種、泌乳階段、胎次、環境等因素影響而發生變化[15]。人乳、牛乳和山羊乳中N-聚糖的結構與含量如表1 所示。

表1 人乳、牛乳、山羊乳中N-聚糖的結構與含量比較
母乳中的糖蛋白具有多種生物學功能。例如,乳鐵蛋白可以抑制病原體,如大腸桿菌、霍亂弧菌和變形鏈球菌;α-乳白蛋白有助于嬰兒吸收鐵和鋅;各種免疫球蛋白可以促進嬰兒免疫系統的發育[23]。作為乳清糖蛋白的重要組成部分,N-聚糖可以保護具有生物活性的蛋白質和多肽不被胃腸道系統降解[24]。LU Y等[15]分別在母親初乳、過渡乳和常乳乳清蛋白中鑒定出20、22 和21 種N-聚糖結構,隨著泌乳期的進行,母乳乳清蛋白中N-糖鏈的巖藻糖基化降低37.5%,唾液酸化增加1.25 倍,與HEGAR B 等[12]的研究結果相符。DALLAS 等[16]在母親常乳乳清蛋白中共鑒定出52種N-聚糖結構,其中84%為巖藻糖基化,47%為唾液酸化(按豐度計算)。母乳N-聚糖結構與含量的差異可能由原料、處理方法、儀器誤差等因素而造成。GOONATILLEKE E 等[25]對不同健康母親在不同哺乳期中的乳蛋白質豐度及個別蛋白質N-糖基化位點進行監測,發現乳鐵蛋白中巖藻糖基化N-聚糖含量在哺乳期間增加。核心巖藻糖可以控制生物活性,如病原體黏附和細胞間信號傳遞[26]。而具有末端巖藻糖的聚糖是Lewis 家族的糖苷,可與病毒和細菌的凝集素受體結合,阻止與上皮細胞的黏附,從而防止病原體侵襲和定殖。另外,分泌型免疫球蛋白A(sIgA)含量雖然在哺乳期中減少,但其中的單唾液酸化和雙唾液酸化聚糖含量升高。sIgA 是保護表面黏膜免受病原體入侵的主要免疫球蛋白。大腸桿菌會導致新生兒腦膜炎和敗血癥[27]。sIgA 中的唾液酸化的多糖與S-菌毛的大腸桿菌結合,以防止其與上皮細胞結合。
在牛乳鐵蛋白的N-聚糖中,分別從初乳、過渡乳和常乳中鑒定出41、33 和22 中N-聚糖結構。與初乳相比,過渡乳和常乳中的總N-糖鏈含量分別降低了1.3 倍和1.7 倍。隨著哺乳時間的延長,復合型和雜合型N-聚糖的含量下降,而高甘露糖型的含量增加。唾液酸化和巖藻糖化的N-糖鏈先增加后減少,它們在過渡乳中的含量分別是初乳和成熟乳的1.8 倍和5.1倍[17]。與母乳相比,牛乳乳清蛋白中的N-聚糖濃度較低,但其糖鏈結構更加復雜,較長的糖鏈結構占總N-鏈聚糖的比例更大[28],巖藻糖基化程度低,唾液酸化程度高[22]。Neu5Gc 的含量也是人乳和牛乳N-聚糖組成的明顯區別之一。人乳N-聚糖的唾液酸主要是Neu5Ac,不含Neu5Gc,而牛乳N-聚糖中二者都包含,這種差異是由于人體中的胞苷酸Neu5Ac 羥化酶在進化過程中發生了突變,而Neu5Gc 的合成只能通過CMP-Neu5Ac 羥化而成,因此人體無法自主合成Neu5Gc[18]。
在山羊乳清蛋白的N-聚糖中,分別鑒定出32、38 和34 種N-聚糖結構。在初乳中的巖藻糖化程度是常乳中的1.66 倍,Neu5Ac 和Neu5Gc 修飾的唾液酸化程度分別是常乳中的3.62 倍和1.69 倍[19]。YUE H等[22]在人、牛和山羊乳乳清蛋白中測得N-聚糖的含量分別為8.8、4.9 和7.9 nmol/mL。與人乳和牛乳相比,山羊乳中巖藻糖基化和唾液酸化低于牛乳,更接近于人乳,這表明羊乳與人乳乳清蛋白中的N-鏈聚糖組成具有同源性[28],山羊乳有望成為嬰兒配方奶粉中人乳N-鏈聚糖的替代品。
2 乳源N-聚糖的生物學功能
乳源N-聚糖具有重要的生物學功能,與乳低聚糖相比,乳中的糖蛋白可能連接多種結構的N-糖鏈,這使其在生命過程中發揮更加多樣的作用。已證實的乳源N-聚糖的作用包括促進腸道有益微生物的定殖、抑制致病菌的生長、抑制病原體的黏附、抗炎、免疫調節、促進大腦發育等。
2.1 作為益生元,促進腸道有益微生物的定殖
腸道內非致病微生物的定殖對嬰兒出生后的健康至關重要。N-聚糖可作為某些特定微生物生長的底物,如雙歧桿菌[29-30]、擬桿菌屬[31]、糞腸球菌屬[32],這些菌通過分泌糖苷酶,將N-聚糖從糖蛋白上釋放下來,使N-聚糖作為碳源或者唯一碳源被微生物利用[33]。KARAV S 等[30]利用內切β-N-乙酰氨基葡萄糖苷酶(EndoBI-1)釋放N-糖鏈,比較了B. infantis EVC001定殖的母乳喂養嬰兒的糞便樣本和缺乏B. infantis EVC001 定殖的對照組嬰兒糞便樣本的N-糖鏈,發現與對照樣品相比,B. infantis EVC001 定殖嬰兒的糞便樣本中N-聚糖的含量要高得多,這說明EndoBI-1在被B. infantis EVC001 定殖的母乳喂養嬰兒中被激活,促進了乳蛋白N-聚糖在腸道中的釋放,從而在嬰兒腸道菌群中完成B. infantis EVC001 的定殖。在一項體外實驗中,人們以乳N-糖蛋白作為選擇性底物富集雙歧桿菌,并促進EndoBI-1 的產生以進行初始去糖基化,發現釋放的N-聚糖促進了細菌的快速生長,超過70%的中性N-聚糖和90%的唾液酸化N-聚糖被嬰兒雙歧桿菌利用[29]。除雙歧桿菌外,人乳中核心巖藻糖基化也有助于新生小鼠腸道中乳酸桿菌的定殖[34]。
N-聚糖的利用程度取決于N-聚糖的存在形式、結構等因素。在存在形式上,與結合在糖蛋白上的糖鏈相比,從糖蛋白上釋放下來的游離形式的糖鏈可以更好地作為芽孢桿菌的碳源使其快速生長[29]。在結構上,人乳N-聚糖比牛乳N-聚糖表現出更強的雙歧活性,這是由于與牛乳N-聚糖相比,人乳N-聚糖具有高水平的短鏈結構,在細胞中更容易被雙歧桿菌利用消耗[20]。此外,N-聚糖結構具有多樣性,它們可以被不同的細菌代謝。TAO J 等[34]在豬乳N-聚糖與其后代腸道菌群之間的關系中發現,高甘露糖型和唾液酸化N-聚糖含量與擬桿菌含量呈正相關,與變形桿菌含量呈負相關。以上研究說明,乳源N-聚糖作為益生元對于促進嬰兒腸道有益微生物的定殖具有重要作用。
2.2 抑制致病菌的生長
目前關于乳源N-聚糖對病原菌的生長抑制研究較少,更多研究集中在N-聚糖對病原體的抗黏附作用。WANG 等[28]通過人乳和牛乳的N -聚糖對食源性致病菌的抗致病性活性比較,發現人乳N-鏈聚糖的抑制作用明顯強于牛乳N-聚糖,這是由于人乳N-聚糖具有高巖藻糖基化結構。研究發現,較高的抗致病活性與巖藻糖基化N -聚糖的水平密切相關[28],并且將巖藻糖從聚糖上移除后活性幾乎消失。這種作用機制尚未可知,因此仍需對這些N-聚糖結構的作用方式進行進一步研究,以了解N-糖鏈的抑菌功能。
2.3 抑制病原體(細菌、病毒等)黏附
細菌、病毒等病原體黏附在宿主細胞上是使其引發疾病的第一步。以細菌為例,它們通過表面的黏附素與宿主細胞上的受體結合,以攝取宿主的營養物質[36]。當細菌與宿主細胞黏附時,大多數細菌的黏附素與受體之間的作用屬于外源凝集素與糖的特異性識別模式。乳中N-糖鏈抑制細菌黏附的機制在于,糖鏈充當細胞表面多糖的類似物,它的存在減少了細菌表面黏附素與宿主細胞受體結合的機會,從而起到抗黏附的作用[37]。雖然N-糖基化使乳蛋白質具有抗菌活性,但糖鏈結合在糖蛋白中會使其抗致病活性受到極大的阻礙,因此只有糖鏈被內切酶釋放下來之后才會發揮更大的抗菌作用[38]。例如,從人乳乳鐵蛋白中純化出的N-聚糖顯著降低了李斯特菌對結腸上皮細胞的侵襲,且乳鐵蛋白經糖苷酶處理后,對腸沙門氏菌和大腸桿菌O157:H7 的活性發生了變化[24],說明腸道中乳蛋白的N-糖鏈不僅在抗體-抗原識別中起信號分子的作用[39],還可以在釋放出來后,作為抵抗細菌入侵的強大屏障。
LU Y 等[15]在人乳、牛乳和山羊乳的游離低聚糖和N-聚糖的抗金黃色葡萄球菌作用的比較中指出,乳N-聚糖的抗黏附作用強于乳低聚糖,且N-聚糖的抗黏附活性依次為人乳>山羊乳>牛乳。這種活性的差異可能是由于不同物種的N-聚糖的結構是不同的,人乳N-聚糖的高抗黏附活性來源于其較高的巖藻糖基化N-聚糖的結構,但是目前尚不清楚這種抗黏附優勢來自一種還是幾種關鍵的巖藻糖基化結構,還是所有結構的協同作用。此外,在乳N-聚糖抑制病原體黏附的過程中,除巖藻糖基化N-聚糖,還有其他的修飾成分也發揮作用,如唾液酸化N-聚糖。研究發現,IgA 的唾液酸化N-糖鏈與病原體之間的特定結合可以保護新生兒免受S-菌毛大腸桿菌感染而引起的敗血癥和腦膜炎[27]。
2.4 抗炎
N-糖基化位于細胞膜表面,具有一定的抗炎作用。炎癥性腸病(IBDs)是由于慢性腸道炎癥導致的腸黏膜損傷[40]。脂多糖(LPS)作為革蘭氏陰性細菌外膜的主要成分,是腸道炎癥最有效的刺激物之一[41]。LPS能夠誘導多種細胞,如巨噬細胞、淋巴細胞、粒細胞、上皮細胞等產生炎癥。腸上皮細胞是阻止病原體進入腸黏膜屏障的重要成分,它通過分泌各種炎癥介質參與先天性和后天性的免疫應答[42]。余秋地[43]以Caco-2細胞為體外模型,LPS 刺激細胞構建炎癥模型,研究了牛乳與人乳N-聚糖對細胞炎癥的影響。結果表明,N-糖基化有助于降低腸道內的過度炎癥反應,人乳N-聚糖比牛乳N-聚糖具有更好的抗炎活性,巖藻糖基化修飾是影響乳中N-聚糖抑制炎癥的關鍵性因素。壞死性小腸結腸炎(NEC)作為一種發病率和死亡率高的炎癥性腸病,主要發生于早產兒 (孕周<37 周)和極低出生體重兒(<1 500 g)中,死亡率高達50%[44]。來自體內實驗表明,乳源核心巖藻糖化N-聚糖對NEC 具有保護作用。研究中大鼠乳汁的核心巖藻糖基化N-聚糖的水平降低,導致結腸促炎因子上調,抗炎因子下調,以及Treg/Th17 細胞的不平衡,導致NEC 后代的腸道炎癥,因此揭示了核心巖藻糖基化N-聚糖作為治療劑的潛力[34]。然而,關于這種糖型如何發揮抗炎作用仍需進一步探索。
2.5 促進大腦發育
乳源N-聚糖對于嬰兒大腦發育的作用主要在于它其中的一種單糖成分——唾液酸。人類中樞神經系統組織的Neu5Ac 濃度高于身體任何其他組織[45]。在整個哺乳期和嬰兒出生時的不同胎齡之間,發現母乳中含有高水平的Neu5Ac,其中21%~28%來自糖蛋白[46]。母乳中的Neu5Ac 對嬰兒的大腦發育和認知具有潛在的重要作用[47]。在新生豬和嚙齒動物中補充結合Neu5Ac,發現Neu5Ac 還可顯著增強學習和記憶能力[48-49]。提供特定的唾液酸化N-糖鏈結構對于Neu5Ac 增強新生兒的大腦發育尤為重要,特別是對于早產兒。因此,未來需要更多的研究來確定哪些結構更有利于Neu5Ac 的釋放。
2.6 調節免疫系統
新生兒的免疫系統是不成熟的,易發生感染和過敏。因此,在生長早期中保證免疫庫的適當平衡對嬰兒的健康發育具有重要意義。腸道微生物群在免疫系統的發育和調節中起著至關重要的作用,微生物群的變化與嬰兒許多慢性疾病的發展有關,如濕疹、哮喘和壞死性小腸結腸炎[37]。研究發現,一些N-聚糖能夠協同調節細胞的增殖與分化,影響T 細胞的活化[41]。母乳中的巖藻糖基化N-聚糖可以幫助腸道乳酸桿菌和雙歧桿菌的定殖,調節Treg/Th17 平衡,從而預防母乳喂養新生兒的腸道炎癥[11、34]。ZHOU 等[50]報道了妊娠期糖尿病(GDM)小鼠乳汁中的N-聚糖會影響其后代卵清蛋白誘導的免疫反應。與正常小鼠相比,GDM 大鼠后代脾B 細胞(CD19+)和T 細胞(CD3+)比例較高,Th1/Th2 細胞(IL-4/干擾素-γ) 和Treg/Th17 細胞(IL-10/IL-17) 動態平衡失衡,這可能與GDM 小鼠乳汁中巖藻糖化和唾液酸化N-糖鏈水平升高有關,因此未來應進一步確定蛋白質N-糖基化對嬰兒免疫系統的影響。
3 在嬰兒食品中的應用與展望
乳源N-聚糖因其豐富的生物學功能被大家廣泛研究。人們可以通過人乳和牛乳N-聚糖的比較,來揭示母乳喂養和嬰兒配方奶粉喂養的嬰兒的健康差異。更重要的是,乳源N-聚糖可作為一種食品配料補充到嬰兒配方奶粉中,通過模擬母乳成分,給嬰兒提供近似于母乳的營養和保護。
牛乳和山羊乳中的N-聚糖是天然聚糖,類似于人乳中的N-聚糖,它們對于嬰兒的生長發育具有重要意義,如巖藻糖基化N-聚糖有望成為嬰幼兒的個性化益生元[11、51]。然而,由于人乳N-聚糖的可獲得性有限,因此牛乳和山羊乳中N-聚糖的富集是當前研究的熱點。乳制品加工中產生的副產物乳清是乳源N-聚糖的潛在來源,每升乳清中約含4.5 g 糖蛋白[52],因此將乳清轉化為天然N-聚糖對乳制品加工具有重要意義。BUNYATRATCHATA 等[51]通過EndoBI-1 酶大規模生產牛乳N-聚糖,并找到其在較低的pH(4.45)、較長的反應時間(475 min)、研究范圍內的中等溫度(52.58 ℃)和較高的酶/蛋白比(1∶333)的條件下產量更高。
嬰兒食品的加工過程中會發生乳蛋白N-糖基化的變化,從而影響產品的特性。XIAO 等[53]在用牛奶制備酸奶的發酵過程中,發現蛋白質N-糖基化模式保持相對不變,表明N-糖基化穩定了蛋白質構象,減少了發酵過程中牛奶中蛋白質的聚集和沉淀。同時,不同的熱處理溫度也會影響N-聚糖的釋放,HTST(高溫短時72 ℃15 s)相較于UHT(超高溫135 ℃3 s)促進了N-聚糖的釋放[51]。此外,由于N-聚糖和低聚糖在組成和結構上的相似性,可以假設乳源N-聚糖也可以被吸收到循環中,到達各種組織靶點,如大腦、腎臟和尿路[54],對研究新生兒腸道菌群和生理結構具有重要意義。
目前,乳源N-聚糖大多數僅用于科學研究,在嬰兒配方奶粉中的應用較少,如何從乳清產品中高效綠色富集N-聚糖仍需進一步研究。此外,乳源N-聚糖作為一種食品配料應用于乳制品中,其在嬰配奶粉中的作用機制、安全性及量效關系尚不清楚,仍需進一步探討。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
