基于文獻計量學的農業種植環境中抗生素抗性基因的研究進展
2023-12-20 01:30:44趙震雄張宇軒王麗平劉瑞志
環境科學研究 2023年12期
趙震雄,張宇軒,王麗平,劉瑞志,湯 利
1. 云南農業大學資源與環境學院,云南 昆明 650201
2. 中國環境科學研究院,國家環境保護河口與海岸帶環境重點實驗室,北京 100012
近年來,具有廣泛傳播性的有害生物污染成為全球關注的焦點. 抗生素作為一種新污染物,具有生物毒性、環境持久性、生物累積性等特征[1-2]. 2022 年5 月,《新污染物治理行動方案》將抗生素列為重點管控新污染物,需采取源頭禁限、過程減排及末端治理的全過程環境風險管控措施. 與抗生素自身危害相比,抗生素抗性基因(antibiotic resistance genes,ARGs)的危害更為嚴重,能夠通過基因的水平轉移打破物種間的限制,實現跨物種傳播[3]. 研究證實,若抗生素耐藥性問題得不到解決,預計到2050 年全球每年因抗生素耐藥性問題死亡的人數將達到1000 萬人,造成經濟損失100 萬億美元[4]. 近年來,因農藥、獸藥、化肥等的大量使用,以ARGs 為代表的新型生物性污染呈現出復雜性和多元化特征,作為新的環境污染問題在農業環境中日益突出. 諸多研究表明,ARGs 會隨著農產品等通過食物鏈進入畜禽動物或人體內,引發的抗生素耐藥性問題已發展成為生態安全和公共健康的嚴重威脅,但目前關于農業環境中ARGs 的系統研究報道仍明顯不足. 因此,亟需深入了解農業環境中ARGs 的污染現狀及環境風險,以期為未來農業環境中抗生素及ARGs 的管控提供理論依據.
1 基于文獻計量學的農業環境中ARGs 的研究熱點分析
采用文獻計量學方法,基于CNKI 和Web of Science 數據庫,系統分析了近5 年來農業環境中ARGs 的研究熱點. 檢索日期為2022 年7 月22 日,設定檢索主題為“Agriculture”或“Agricultural”中精煉檢索“Antibiotic Resistance Genes”,去重后分別得到CNKI 的183 篇和Web of Science 的115 篇論文. 利用CiteSpace[5]對檢索到的文獻關鍵詞進行聚類統計,并繪制Timeline 圖譜(見圖1). 基于CNKI〔見圖1(a)〕和Web of Science〔見圖1(b)〕兩種檢索方式,聚類模塊值(Q值)分別為0.8443 和0.7326,聚類平均輪廓值(S值)分別為0.9608 和0.9008,表明CiteSpace 能夠對農業環境中ARGs 的數據集準確歸類. 關鍵詞的整體網絡Density 值分別為0.0140 和0.0216,表明目前農業環境中ARGs 的研究較少且不聚焦. 進一步分析Timeline 圖譜,結果表明農業環境中ARGs 的研究主題隨著時間發生明顯變化. 具體而言,2017 年,相關研究主要集中于抗生素及其ARGs 在水和土壤環境中的相關性及時空變化規律. 2019 年起,關于抗生素及ARGs 的研究則更多聚焦于水庫、河流及海水養殖等人為干擾較多的環境. 截止到2022 年,農業種植環境中ARGs 的研究逐漸發展為一個新的研究熱點,主要聚焦于探究有機肥、畜禽糞污和生產生活廢水對農業種植環境中ARGs 生物積累的影響及其環境風險,但相關研究報道仍明顯不足.
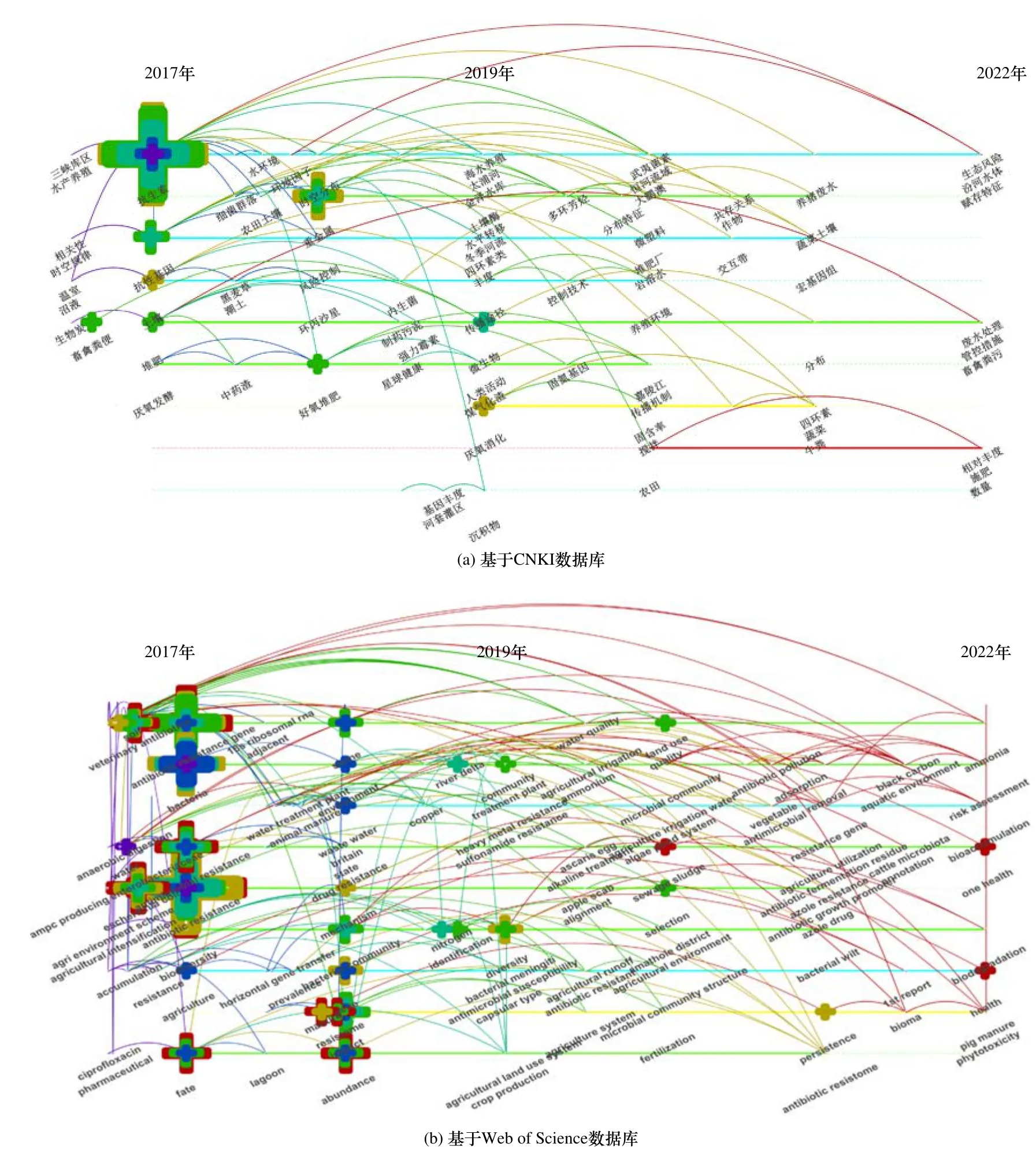
圖1 基于CNKI 和Web of Science 數據庫的農業環境中ARGs 相關研究文獻的CiteSpace 分析Fig.1 CiteSpace analysis of ARGs research literatures in agricultural environment based on CNKI and Web of Science
進一步通過Bibliometrix[6]基于Louvain 算法得到近5 年農業環境中ARGs 的研究主題演化〔見圖2(a)〕,由2017—2019 的7 個主題演變到2020—2022 年的10 個主題,表明該領域ARGs 的相關研究開始趨于多元化和深入化. 其中,“Agriculture”和“Diversity”兩個主題分化為“Biological-control”“Identification” “Agriculture”“Diversity”等多個研究主題,推測與近年來農業生產中利用生物多樣性和土壤修復達到提高產量的目的相關. 然而,當前環境中依然殘留著大量的抗生素,張煥軍等[7]發現湖泊和河流水體主要以溶解性較強的磺胺類和化學性質相對穩定的四環素類為主,沉積物和土壤中則以性質穩定的喹諾酮類、大環內酯類和四環素類抗生素為主[8-9]. 更為重要的是,農業土壤環境中ARGs 的多樣性和耐藥性隨著時間推移仍在不斷發展,主題“Soil”和“Fate”逐步演化為“Diversity”“Resistome”和“Treatment plant”等. 通過Vosviwer[10]計算并繪制農業環境中ARGs 研究主題的相關性熱圖〔見圖2(b)〕,結果表明,微生物群落、耐藥菌、重金屬、MGEs 和糞肥均與農業種植環境中ARGs 的研究緊密相關,加之“Soil”和“Bacteria”兩個主題分化出多個研究主題〔見圖2(a)〕,表明土壤中細菌是ARGs 的主要載體.農業種植環境逐漸成為抗生素耐藥性發展的重要屬地,需要進一步深入了解農業種植環境中ARGs 的來源、遷移、影響因素及管控措施等.
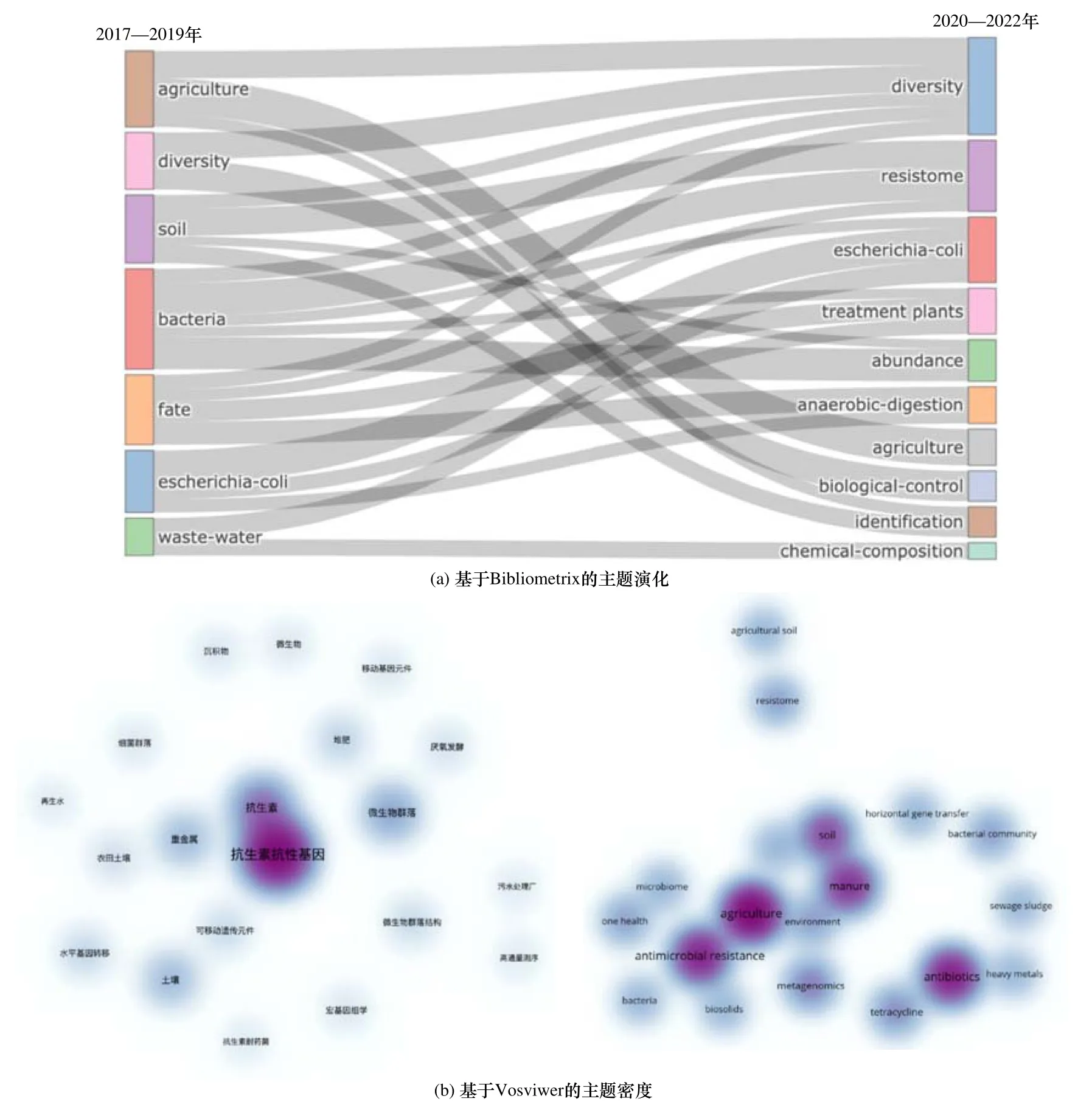
圖2 基于Bibliometrix 主題演化和Vosviwer 主題密度的農業環境ARGs 的研究概況Fig.2 The general research overview of ARGs in agricultural environment based on Bibliometrix theme evolution and Vosviwer theme density
2 農業種植環境中ARGs 的污染來源及賦存特征
2.1 ARGs 的污染來源
ARGs 一般是由于細菌對抗生素產生耐藥性所致,主要分為內源抗性、適應性抗性和獲得性抗性.內源抗性是指細菌本身的固有屬性,Chen 等[3]在人為干擾較少的土壤中檢測到完全不同于現代抗性組的β-內酰胺類和氟喹諾酮類ARGs,劉燕等[11]在貢嘎山受人為干擾較小的極高海拔區(2948~3651 m)土壤中發現了132 種ARGs 和10 種MGEs. 然而,內源性抗性產生的ARGs 豐度顯著低于受人為干擾較多的土壤環境,細菌的適應性抗性和獲得性抗性是ARGs 在環境中持續富集的主要原因.
適應性抗性是指環境背景值變化或抗生素亞抑制水平導致基因和或蛋白質表達水平暫時改變,促進細菌產生適應性耐藥性. 獲得性耐藥性是指在細菌的繁殖生長過程中,通過對環境中外源遺傳物質行整合產生獲得性耐藥性. 據統計,2013 年我國36 種化學類抗生素使用總量約162000 t[2],其中四環素類、磺胺類、喹諾酮類、大環內酯類和β-內酰胺類抗生素使用范圍廣且用量大,分別占使用總量的5%、7%、17%、26%和21%,廣泛應用于農業、畜牧業和醫藥行業. 基于Vosviwer 的主題密度分析表明ARGs 與抗生素和微生物群落密切相關〔見圖2(b)〕,因此,農業環境中抗生素耐藥性的發展并不僅是微生物自身所分泌物質導致,而是在農業環境中抗生素增加的背景條件下,細菌更容易產生適應性抗性和獲得性抗性,導致ARGs 持續累積并多元化發展.
農業種植活動中施肥、施藥、灌溉等生產方式[12]極大促進了ARGs 的富集及耐藥微生物的發展[2,13],農業種植環境面臨著嚴峻的ARGs 挑戰. 在我國主要農業產區(見表1),土壤種植環境中ARGs 平均豐度為2.49×1010~3.09×1010copies/g(絕對豐度)和5.7×10—3~2.39×10—1(相對豐度)[14-20],遠高于水環境中ARGs 的豐度[21]. 通過共現網絡分析(見圖3)發現,農業種植環境中ARGs 與獸用抗生素密切相關,“Bacteria”和“Abundance”是ARGs、抗生素耐藥性和獸用抗生素間的重要樞紐.

表1 不同農業種植環境中的ARGs 豐度Table 1 Abundance of ARGs in different agricultural planting environment
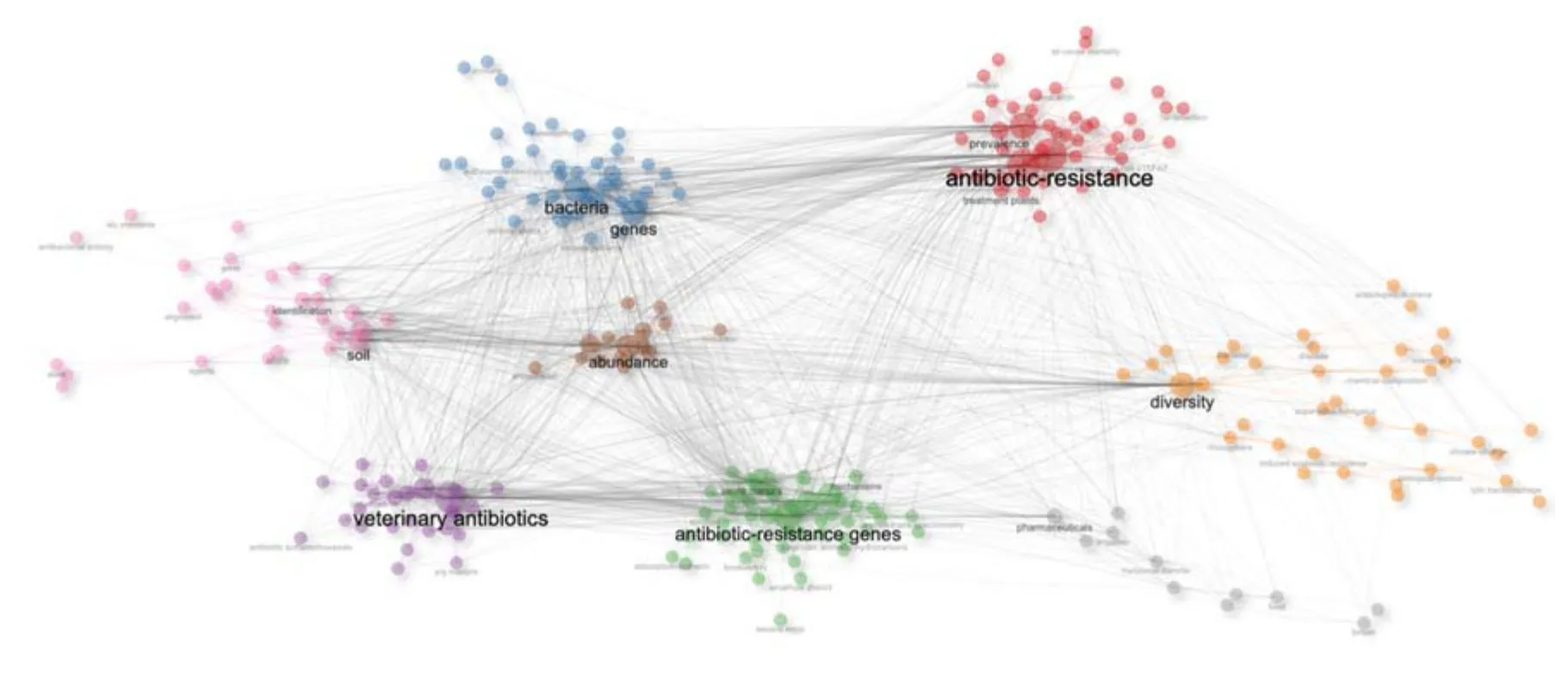
圖3 農業環境中ARGs 相關研究的關鍵詞共現網絡圖Fig.3 Co-occurrence network of keywords in ARGs related research in agricultural environment
研究表明,40%~90%的抗生素不能被生物完全吸收[22-24],僅2013 年被人類和動物排泄的抗生素就高達54000 t[2]. 糞肥糞水或農家肥等的施用導致大量生物殘體和代謝產物進入種植環境,抗生素及其ARGs 會伴隨該過程同步進入土壤,細菌隨即對其產生適應性抗性和獲得性抗性,從而加速ARGs 的發展. 施用糞肥[25]和長期堆放糞肥[26]土壤中的ARGs相對豐度分別為5.02×10—4~3.00×10—3和2.94×10—2~1.55×10—1,施用有機肥土壤中的噬菌體攜帶的ARGs總豐度顯著高于不施肥和施用化肥的土壤[27],連續施用兩年糞水的土壤中ermF和ermB的污染指數顯著增加[28]. 石禮虎等[29]研究發現,與空白對照組相比,施用抗生素菌渣(高/低施用量實驗組) 能夠顯著提高ARGs 的檢出率和檢出豐度. 殺菌劑添加也會促進土壤中ARGs 豐度的升高. 張厚樸等[30]利用三唑酮、百菌清、嘧菌酯和多菌靈四種殺菌劑處理設施土壤60 天后,ARGs 總豐度是未添加殺菌劑土壤的1.02~1.98 倍,是森林土壤的1.12~2.16 倍. 此外,人類生產生活廢水,尤其是畜禽養殖、醫藥行業廢水等通常會經污水處理廠處理后排放到自然環境或再生利用,但現有污水處理設施對抗生素及ARGs 的去除效果極其有限. 農業種植灌溉對水資源需求量很大,抗生素及其ARGs 會隨河湖水體、地下水、再生水灌溉等進入農田土壤,極大程度上促進了農業種植土壤環境中ARGs 的富集和發展(見圖4).
2.2 ARGs 的賦存特征
細菌受到抗生素脅迫時,會啟動主動防御機制增強抗生素耐藥性[31],增加細胞分泌物和代謝水平,表達更多的外排泵基因,促進質粒、轉座子和整合子等可移動遺傳元件(Mobile Genetic Elements,MGEs)的產生和釋放[32]. ARGs 在環境中可分為胞內ARGs(Intracellular ARGs,iARGs)和胞外ARGs(Extracellular ARGs,eARGs). 研究表明,約有60%和40%的ARGs分別存在于DNA[25]和MGEs 中[33-34],Zou 等[25]研究發現MGEs 和iARGs 的相對豐度存在強相關性. 由此推知,iARGs 容易被細菌體內的遺傳物質(Intracellular DNA,iDNA)和MGEs 吸收和轉導,而eARGs 則容易隨著細菌在構建生物膜的過程中排出體外,進而被環境中游離的DNA(Extracellular DNA,eDNA)和MGEs捕獲[35].
農業種植土壤中的微生物能夠通過分泌核酸、多糖、蛋白質和糖醛酸等胞外聚合物(extracellular polymeric substances,EPS) 以適應環境中的抗生素、除草劑和重金屬等帶來的壓力[36]. EPS 中富含大量的DNase 酶,能夠水解ARGs[31]和eDNA[37],因此在理論上土壤中EPS 對于ARGs 的發展是阻礙的. 然而,在人為影響較大的土壤中,抗生素、重金屬等污染物大量存在,這些污染物能夠被EPS 吸收并與DNase 酶的特征位點結合[37],導致DNase 酶的活性降低,從而使得ARGs 免受DNase 酶的降解并長期存在. Fan 等[38]研究發現,厭氧氨氧化過程中產生的EPS 中的蛋白會分別在GLU-307、hs-191、ASP-318和tr-32 等多個位點與紅霉素和磺胺甲惡唑結合,在EPS 對抗生素的長期馴化過程中致使ARGs 產生和累積.
此外,EPS 能夠吸收環境中的eDNA、eARGs 和MGEs,且土壤環境中的EPS 比水環境中更穩定.Pathan 等[39]研究發現土壤中eDNA 含量是水體中的3~4 倍,Wang 等[40]研究發現,EPS 內部ARGs 絕對豐度比相應的游離ARGs 高0.2~4.6 個數量級,且DNA轉化體含量證明與EPS 相關的ARGs 轉化效率比游離ARGs 高3.3~236.3 倍. 由此表明,農業種植環境中EPS 和eDNA 能夠充當ARGs 增殖的潛在遺傳物質來源和載體[33],導致ARGs 在農業種植環境中廣泛分布并持續累積[41].
3 農業種植環境中ARGs 的遷移及風險
與常規的化學污染物不同,ARGs 是具有遺傳信息的DNA 片段,位于染色體上的ARGs 能通過染色體的自我復制遺傳在細菌親代之間垂直傳播,發生垂直基因轉移(vertical gene transfer, VGT). 更多研究表明,ARGs 的潛在生態健康風險普遍是由跨物種的基因水平遷移(horizontal gene transfer,HGT) 造成的.ARGs 相關的基因盒、質粒、轉座子和整合子等MGEs廣泛存在于細菌中,基因盒和整合子捕獲ARGs,質粒進行接合和轉移ARGs,轉座子和插入序列轉移ARGs,不同生物間可以利用這些MGEs 作載體,通過轉化(細菌-環境介質-吸收遺傳物質)、接合(細菌-菌毛介質-傳遞遺傳物質)、轉導(細菌-噬菌體-傳遞遺傳物質)等方式使ARGs 在細菌間相互傳播,最終導致抗生素耐藥性通過基因水平遷移的方式廣泛傳播.所以,ARGs 在細菌群落間的基因水平遷移和垂直基因轉移是導致ARGs 造成環境和健康風險的兩個重要驅動力.
研究表明,iARGs 和eARGs 的HGT 過程并不相同. Zou 等[25]研究發現,iARGs 與質粒上的整合子(R=0.981,P<0.001)與轉座子(R=0.908,P<0.001)的相關性遠高于eARGs 與質粒上的整合子(R=0.591,P<0.05)與轉座子(R=0.779,P<0.05) 的相關性,表明iARGs 主要通過細菌間iDNA 的接合、轉導及MGEs發生水平遷移,而eARGs 則主要通過eDNA 的自然轉化[34]和MGEs 進行水平轉移[35],進而證明ARGs的HGT 主要依賴于DNA 的遺傳特性和MGEs 的跨物種遷移能力. 然而,并非所有的DNA 都能成功插入細菌的基因中,這主要取決于外源DNA 鏈和染色體DNA 之間是否存在同源區域. 例如,將DNA 整合到革蘭氏陰性桿菌基因組的過程中,相較于其他DNA,同源DNA 更容易被轉化,其轉化率高達109倍[42]. 研究表明,eDNA 的長度為20~200 bp,更容易插入細菌中進一步表達[35],當eDNA 與受體基因組同源的DNA 相連時,整合轉化的概率能夠增加105倍[42]. Ye 等[34]研究發現,eDNA 的自然轉化不需要特定的蛋白質,能夠通過維持銅綠假單胞菌生物膜結構[43]誘導細菌富集氨基糖苷類ARGs[44],影響攜帶ARGs 的質粒在不同細菌間的自然轉化頻率[45],從而加速MGEs 和ARGs 被其他細菌轉化的進程. 此外,土壤中穩定的EPS 不僅可以長時間保持微生物間的物質交換相對靜止,還能通過自身組分連接兩個細胞的菌毛,從而更加有利于細菌之間的單向質粒交換和基因交換過程[40]. 這些結果意味著土壤環境中EPS的穩定存在、eDNA 的自然轉化和MGEs 的遷移可能是農業種植環境中ARGs 進行HGT 的主要方式之一.
農業生產活動導致抗生素及ARGs 通過糞肥、糞水、灌溉污水等多種方式進入農田土壤環境,人類可以通過多種途徑接觸農業種植環境中的ARGs 并在體內蓄積,細菌群落及基因的水平和垂直轉移是導致ARGs 造成環境和健康風險的兩個重要驅動力. 抗生素及ARGs 進入土壤會直接影響土壤微生物和酶活性,導致耐藥菌產生和增殖,改變土壤微生物群落結構,破壞土壤生態系統平衡. 一旦土著微生物獲得ARGs,因其具有良好的環境適應性,會以超過親代菌株的效率擴散,給土壤種植環境的生態安全帶來更大威脅. 隨著施肥施藥時間不斷延長,ARGs 不斷累積并在土壤不同層級遷移[26]. 農田土壤中的抗生素及ARGs 能夠通過淋溶[46]、滲透等方式進入到地下水環境[47],并通過地表或地下徑流等方式進入河流、湖泊等水環境中,導致ARGs 在不同環境中交叉污染,從而促進多重耐藥基因的產生(見圖4),造成更為嚴重的環境風險.
植物的莖葉、果實等可食用部分是抗生素耐藥性從農業種植環境向人類傳播擴散的主要途徑[16],植物根際土壤中的ARGs 可以向植物體內組織遷移[48].研究表明,施用有機肥導致生菜葉際ARGs 豐度是傳統生菜中的8 倍[49],長期施用雞糞會顯著增加玉米葉表ARGs 豐度,ARGs 種類多達124 種[1],超市購買的蔬菜(沙拉、芝麻菜、香菜等)中同樣檢出了攜帶四環素類ARGs 的可進行自主轉移的質粒[50]. 由此表明,土壤中的ARGs 能夠通過農作物從農業種植環境向人類傳播擴散[49],對人類健康造成較大風險.
4 農業種植土壤中ARGs 的影響因素
4.1 不同農用地利用模式及土壤性質對ARGs 的影響
農作物種植模式和種植作物類型均會影響ARGs的賦存狀況[19]. 例如,在設施菜地、大田和糧田等不同的農地利用方式下,設施菜地ARGs 豐度顯著高于大田及糧田. 王佳佳等[51-52]研究發現,溫室土壤中tet類抗性基因和intI-1基因的豐度及菜地中ARGs 豐度均顯著高于大田土壤(P<0.05),推測與有機肥施用量高、環境密閉、環境溫度高等因素相關,最終導致ARGs 在不同種植模式下的累積差異. 此外,黃福義等[19]研究不同種植作物時發現,同樣施肥方式下種植不同作物會導致ARGs 豐度呈現顯著差異(P>0.05),其中香蕉種植土壤環境中ARGs 的絕對豐度(1.41×1010copies/g)最高,其次為花生(1.28×1010copies/g)、水稻(1.10×1010copies/g)、甘蔗(7.96×109copies/g) 和柑橘(6.47×109copies/g),推測由于不同作物通過根系生長和滲出過程改變土壤理化性質,并富集不同類型細菌群落,從而直接或間接改變ARGs 的賦存和演變特征[53-54]. 此外,秸稈還田同樣與ARGs 的發展存在一定關系. Zhang 等[55]研究了秸稈還田對稻田土壤中磺胺類抗生素(SMX)及其ARGs 的影響,結果表明,對于抗生素,秸稈還田可以通過共代謝作用加速SMX的降解;對于ARGs,秸稈還田作用10 天時,intI-1和sul1基因豐度顯著高于空白對照組,但當作用60 天后,intI-1和sul1基因豐度則顯著低于空白對照組,其中溶解性有機碳及變形菌門和放線菌門細菌在ARGs 消除中發揮了積極作用.
土壤自身性質對ARGs 有重要影響,如土壤類型、土壤pH、水分等. Wang 等[56]研究發現,潮土和鹽堿土ARGs 的多樣性和相對豐度比褐土更高,且潮土中ARGs 的相對豐度與MGEs 的相對豐度具有很強的相關性. 土壤pH 的升高會增加抗生素在土壤中的移動性,從而增加抗生素耐藥遷移風險. 土壤水分則能影響土壤間隙度、吸附系數和離子電位等. 有機質分解的中間產物[41](如鏈球霉素和青霉素)則會對微生物群落和ARGs 的賦存產生影響,如碳、氮、磷作為生物合成DNA 及生長代謝的基本營養元素,其含量與土壤中的ARGs 密切相關. Qian 等[57]研究表明,土壤中的氮會導致部分酶的功能富集和ARGs 的轉移;Sun 等[58]研究發現,通過施加植物和微生物所需的氮會增加土壤和蔬菜中blaTEM、cmlA、sul1和tnpA-4的豐度,表明氮形態的改變及其含量的增加能夠促進ARGs 的富集和轉移[57-60]. Meta 分析證實,ARGs豐度與土壤碳和磷濃度呈顯著正相關(P<0.05)[61],且ARGs 豐度與有效磷濃度的相關性強于ARGs 豐度與總磷濃度的相關性. 另一項研究[62]指出,當土壤中的營養元素含量發生變化時,會產生暫時性的營養饑餓效應,細菌通過養分的饑餓來感受環境中營養的豐富度,從而調節自身生長,但饑餓反應受到干擾會導致細菌DNA 損傷誘導反應(SOS 反應)發生,促進細菌耐藥性和ARGs 產生(見圖4). 例如,當細菌受到氨基酸饑餓信號刺激時會誘導relA 和spoT 基因的表達,增加了鳥苷四磷酸分子(ppGpp)的含量,從而抑制肽聚糖、DNA 等相關物質合成[63]. 有研究[64]指出,在營養缺乏的條件下,ppGpp 通過調節轉錄、翻譯水平和細胞周期,改變細胞的新陳代謝和生理狀態,有利于增強細菌的耐藥性. Rodionov 等[65]研究發現,ppGpp 的積累可以使得大腸桿菌對青霉素的敏感性降低. 因此,農業種植土壤環境中,不合理施肥及養分變化均會對細菌造成選擇壓力[66],增加細菌對抗生素的敏感性或脅迫效應,促進ARGs 的產生、累積和擴散.
4.2 抗生素及其他污染物對ARGs 的影響
抗生素可以通過有機肥、糞肥和糞水施用等方式進入土壤環境,隨即發生吸附、解析、轉化、固定等環境過程,從而長期存在于土壤中. 當土壤中抗生素殘留達到了細菌的亞抑制水平,會產生兩種反應:①它能夠激活細菌的雙組分信號調節系統[62](twocomponent regulatory systems,TCSs)和抗毒素系統[67](toxin-antitoxin system,TAs),提高抗生素耐藥性. TCSs通過磷酸化介導調節下游基因表達,作用于細菌細胞膜的表面修飾、通透性改變和生物膜形成[62];而TAs能夠通過抑制多聚磷酸酶(PPX)增強鳥苷酸分子累積和偶聯,誘導細菌生長停滯促進耐藥性形成[64].②抗生素殘留會直接抑制細菌的DNA 復制和修復、蛋白質及生物膜的合成,從而引發細菌SOS 反應,如喹諾酮類、氨基糖苷類、四環素類和氯霉素類抗生素都可以誘導大腸桿菌和霍亂弧菌中細胞內的SOS 反應[68]. 正常情況下,SOS 反應由recA和lexA基因調節,lexA基因表達的LexA 蛋白會以二聚體形式阻礙RNA 聚合酶與DNA 啟動子結合,抑制SOS 相關基因的表達,使得基因隨機突變率降低,每條染色體每代突變率僅為10—8~10—6[69]. 但是環境或菌群的不穩定可能引發細菌的DNA 損傷,進而產生更多單鏈DNA(ssDNA);recA基因表達的RecA 蛋白會在ATP存在下與ssDNA 結合形成復合物[70],并作為信號分子作用于LexA 蛋白,使之發生自催化水解,降低對DNA 修復、突變以及細胞分裂等相關蛋白的轉錄抑制作用. 細菌便會發生易出錯的DNA 跨損傷合成,導致細菌基因的突變頻率提高10~10000 倍,并誘導細菌產生抗藥性表型的相關基因[71-72]. 例如,Escherichia coli的DNA 修復機制中sulA基因暴露于抗生素等殺菌性物質時,阻礙DNA 損傷修復導致基因組不穩定,從而促進細菌產生ARGs[73].
另有研究[37]顯示,在有機污染物和重金屬等污染物共存的條件下,亞抑制濃度的抗生素更容易誘導細菌的SOS 反應相關基因表達,從而促進多重耐藥ARGs 富集. 有機污染物和重金屬對ARGs 的促進作用主要表現在以下幾方面:①有機污染物和金屬離子都能促進細菌的MGEs 產生. 例如,細菌對微塑料的降解中MGEs 會伴隨著大量有機物降解基因表達而增加[74];重金屬使得細菌胞內胞外滲透壓增加,MGEs 會伴隨著外排泵系統大量表達而增加[75]. ②有機污染物(如十二烷基磺酸鈉[37]和取代芳香烴[31])和重金屬離子(Cu、Ag、Cr 和Zn)[75]都能夠與生物膜中的一些物質結合改變細菌生物膜的通透性,在細菌體內誘導協同抗性[66]. ③部分有機污染物能夠促進ARGs 在細菌體內轉化. 例如,0.5 μmol/L 的取代芳香族化合物會促使ARGs 到大腸桿菌COLIK-12 的轉化效率增加1.57~2.29 倍[31]. 此外,環境中重金屬和有機污染物還會降低EPS 的滲透性和DNase 酶的活性[76],阻礙DNase 酶對ARGs 降解,促進ARGs 跨越EPS 滲透性屏障進入土壤中[77],從而促進ARGs 的富集和遷移. 因此,伴隨著施肥、用藥、灌溉等進入到農業種植環境中的多種污染物均會促進ARGs 的富集和遷移.
4.3 農業種植環境中鹽漬化對ARGs 的影響
農業種植過程中,人們時常會忽略不同作物生長所需的養分條件及土壤環境背景現狀,盲目施肥施藥以達到作物高產的目的,使得農田土壤中積累了大量無機和有機污染物等,加劇了土壤鹽漬化,限制了養分利用,從而可能加劇ARGs 污染及水平遷移的風險. Xu 等[76]研究表明,在含Al 的高鹽土壤和含Ne的中性鹽土中ARGs 和MGEs 的絕對豐度和相對豐度均顯著增加,且鹽脅迫條件下利于刺激細菌合成蛋白質,增加EPS 的分泌,促進ARGs 在土壤環境中的持久性[78]. 例如,泛生菌(Pantoeasp.)、芽孢桿菌(Bacillussp.)、放線菌(Actinomycetessp.)、根瘤菌(Rhizobiumsp.)、節桿菌(Arthrobactersp.)和假單胞菌(Pseudomonassp.) 等均會在鹽脅迫下增加EPS 的分泌[79]. 鹽脅迫會導致細菌分泌的EPS 中的多糖含量增加,不僅會促進EPS 對ARGs 和MGEs 的吸附能力[80],還會抑制細菌對ARGs 的降解.
雖然研究表明EPS 中的DNase 酶能夠與ARGs[31]和eDNA[37]相互作用將其水解破壞,但農業種植土壤中由于過量施肥施藥帶來的有機污染物和金屬離子會與EPS 相互纏繞,導致DNase 酶的三級結構和活性位點發生改變[37],最終導致DNase 酶對ARGs 的降解受阻. 例如,EPS 中攜帶的eDNA 能夠與無機陽離子和磷酸鹽形成緊密的結合鍵能[33],十二烷基磺酸鈉[37]、取代芳香烴[31]和抗生素[38]等物質均能與EPS、eDNA 和質粒結合.
另有研究指出,鹽漬化會抑制尿素水解和硝化過程,限制土壤生物和微生物對營養元素的利用效率,導致土壤中游離氨積累[81]. 土壤中游離氨能夠進入細菌細胞,與磷脂雙分子層中的脂肪酸殘基發生反應形成氨酸鹽,促進細菌細胞膜上的脂質雙層蛋白質通道更易于開放[82-83],導致細菌細胞膜通透性增加,細胞內外的物質交換增加,這一過程可能促進細菌釋放iARGs 和MGEs 或從環境中吸收eARGs 和MGEs,從而加速農業種植環境中的ARGs 的水平遷移過程.綜上所述,農業種植環境中的土壤鹽漬化問題可能促進著ARGs 在農業種植環境中水平遷移和環境持久性[43],進一步加劇了ARGs 的環境風險.
5 ARGs 的削減與管控
目前,ARGs 管控策略主要集中在減少和限制抗生素的使用. 例如,臨床上積極推廣和應用疫苗,在劑量和療程上優化抗生素的使用,畜禽養殖中通過強化養殖管理、優化飼料配方及疫苗接種減少抗生素的使用. 由于抗生素類藥物的化學性質穩定且溶解性強,導致其在環境中持久性及遷移性強. 對于早已面臨較高水平ARGs 污染且長期種植的土壤,源頭管控措施已難以滿足ARGs 的削減需求. 畜禽養殖中,大量抗生素會隨畜禽糞便進入環境,堆肥及高溫發酵可以有效減少糞便中的抗生素,Yue 等[54]研究發現,高溫堆肥主要通過降解抗生素促進ARGs 的削減. ARGs 的控制與抗生素有所不同,李厚禹等[84]研究發現,雖然好氧-厭氧兩相堆肥能有效去除ARGs,但strB、ermB、sul1和sul2等ARGs 和潛在耐藥致病菌仍存在增殖現象. 姜欣然等[85]發現,在高溫堆肥過程中aadA、sul2、mcr-1和oqxB的消減率分別為89.39%、97.99%、99.89%和99.81%,intI-1基因的消減率高于80%,但大多數ARGs 的相對豐度表現出先降低后升高的趨勢. 另有研究[48,86]指出,豬糞生物脫氮堆肥處理后,產物中部分抗生素、ARGs 及人畜共患病病原菌的濃度/豐度均會有所降低. 綜上,雖然堆肥工藝能夠減少部分ARGs 的豐度,但并不能將ARGs 降低到零風險水平,甚至導致部分ARGs 在堆肥區發生多樣性變化并長期存在[26].
另有研究利用外源添加生物炭降低土壤中ARGs的污染水平. Wan 等[87]研究發現,ARGs 豐度與有機碳含量間呈顯著負相關,進一步研究表明生物碳能夠吸附并促進土壤中抗生素的降解,從而減少細菌耐藥性產生機率和遷移效率. 段曼莉等[88]研究表明,蘋果樹枝制備的生物炭可以增加土壤對土霉素的吸附能力,降低土壤、生菜葉和根系中土霉素和ARGs 的累積和遷移[49]. 需要注意的是,生物碳并不能有效控制所有的ARGs,Shi 等[89]發現生物炭雖然能夠降低土壤中tetW、sul2和ermB的豐度,但它顯著增加了tetX和intI-1的豐度. Zhang 等[55]發現秸稈還田可以通過增加土壤有機碳促進微生物群落的共代謝作用加速磺胺類抗生素的降解,但第90 天土壤中sul1、intI-1和sul3的豐度為第10 天的1~1.5 倍. 此外,噬菌體療法也是控制抗生素耐藥性的新方法,生物炭-噬菌體協同作用可以提高土壤和生菜內生細菌群落的結構多樣性和功能穩定性,阻控ARGs 或耐藥菌的遷移,促進微生物對有機污染物的生物強化降解[90].
綜上,目前農業種植環境中控制ARGs 的方法主要包括兩個方面:一是通過堆肥發酵等高溫熱解方式減少抗生素和耐藥菌,從而實現對ARGs 的初步削減;二是通過活性炭等外源物質添加降低污染物有效性,增強土壤微生物穩定性,以此達到對土壤中ARGs 的進一步控制[91-92],但兩種方式均不能實現所有ARGs的有效去除,且去除效果并不顯著,甚至造成部分ARGs 的累積. 因此,對于控制農業種植環境中的ARGs 管控可以從以下幾方面著手:一是優化有機肥的生產工藝,控制其中的抗生素及ARGs 殘留;二是優化養分和肥料配方,減少重金屬及其他污染物進入土壤環境;三是針對ARGs 特性,開發能夠對其進行高效吸附和降解的新材料.
6 結論與展望
6.1 結論
a) 2017—2022 年的文獻計量結果表明,農業種植環境已發展成為抗生素耐藥性發展的新興歸屬地,面臨較為嚴重的ARGs 污染問題,但相關研究卻相對較少且不聚焦.
b) 施肥、施藥、灌溉等農業生產方式極大促進了抗生素耐藥性的發展,細菌的適應性和獲得性抗性是ARGs 在環境中持續富集的主要原因,ARGs 的遷移主要歸因于MGEs 的水平轉移、細菌間iDNA 的轉導、eDNA 的自然轉化和EPS 的物質交換等過程,導致ARGs 在農業種植環境中廣泛分布.
c) 不同農地利用模式下,土壤理化性質、營養元素、抗生素及重金屬等均會影響ARGs 的賦存特征,農業生產引發的土壤鹽漬化和污染物富集問題會增加細菌對抗生素的脅迫效應和敏感性,加速ARGs 在環境中的累積和遷移.
d) 農業種植環境中的ARGs 可以通過食物鏈等多種途徑進入生物體內并不斷蓄積,細菌群落及基因的水平和垂直轉移是導致ARGs 風險累積的重要驅動力,對生態環境和人體健康造成嚴重威脅.
e) 堆肥發酵和外源有機碳添加是控制農業種植環境中抗生素耐藥性的主要方式,但其作用效果有限,仍需進一步研究新技術新方法,實現對種植環境中早已大量累積的ARGs 的有效控制.
6.2 展望
農田土壤是農業環境中抗生素耐藥性傳播和擴散的重要載體,存在較高的環境風險和人體健康風險,必須采取有效措施降低ARGs 和耐藥菌污染風險. 要實現此目標,建議從以下方面著手:
a) 重視抗生素的源頭削減和控制,加強生產管理及對生產廢水和廢棄物的管理.
b) 深入探討不同種植環境中MGEs、iARGs 和eARGs 的賦存特征,并探究其組成成分對ARGs 積累和遷移風險的影響機制.
c) 利用科學方法進一步優化養分及肥料配比,削弱污染物和養分變化對微生物的脅迫效應,降低微生物適應過程中ARGs 產生和累積的概率.
d) 進一步開展農業環境中的ARGs 的演變特征研究,構建其在環境介質中轉移的數學模型,進一步制定并完善ARGs 從土壤環境到人類病原體細菌傳播的風險評估準則.
猜你喜歡
今日農業(2022年1期)2022-11-16 21:20:05
今日農業(2022年3期)2022-11-16 13:13:50
今日農業(2022年2期)2022-11-16 12:29:47
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
今日農業(2021年14期)2021-11-25 23:57:29
中老年保健(2021年12期)2021-08-24 03:30:40
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科技傳播(2019年22期)2020-01-14 03:06:54
