稀土元素對L-酪氨酸發酵的影響研究
2023-12-25 22:37:26王碩時唐恩趙春光徐慶陽
中國調味品 2023年12期
王碩 時唐恩 趙春光 徐慶陽

摘要:目前工業上主要采取微生物發酵法生產L-酪氨酸,此工藝存在生產周期長、糖酸轉化率低等問題。因此,該研究以發酵過程中菌體量、L-酪氨酸產量、糖酸轉換率以及耗糖速率為指標,通過單因素實驗和正交實驗探究La3+、Ce3+、Nd3+對L-酪氨酸發酵的影響并最終確定了稀土元素的添加組合,提高了菌體的活力,縮短了延滯期,延長了產酸高峰期。實驗結果表明,向L-酪氨酸發酵培養基中添加80 mg/L La3+、2 mg/L Ce3+、0.3 mg/L Nd3+對發酵有利。發酵耗時35 h,縮短了2.2%;最高菌體量達到80.3,提升了10.8%;產酸量達到55.7 g/L,提升了9.7%;糖酸轉化率為23.4%,提升了6.4%。驗證了添加稀土元素的可行性,為L-酪氨酸大規模工業化生產提供了依據。
關鍵詞:大腸桿菌;L-酪氨酸;稀土元素;耗糖速率
中圖分類號:TS201.3????? 文獻標志碼:A???? 文章編號:1000-9973(2023)12-0001-06
Study on Effect of Rare Earth Elements on Fermentation of L-Tyrosine
WANG Shuo1, SHI Tang-en1, ZHAO Chun-guang2, XU Qing-yang1,3,4*
(1.College of Bioengineering, Tianjin University of Science and Technology, Tianjin 300457, China;
2.Ningxia Eppen Biotech Co., Ltd., Yinchuan 750100, China; 3.National and Local
United Engineering Laboratory of Metabolic Control Fermentation Technology,
Tianjin 300457, China; 4.Tianjin Engineering Laboratory of Efficient and
Green Amino Acid Manufacturing, Tianjin 300457, China)
Abstract: At present, microbial fermentation method is the main method to produce L-tyrosine in industry, which has problems such as long production cycle and low sugar-acid conversion rate. Therefore, in this study, with the bacterial biomass, L-tyrosine yield, sugar-acid conversion rate and sugar consumption rate in the fermentation process as the indexes, the effects of La3+, Ce3+ and Nd3+ on the fermentation of L-tyrosine is explored through single factor experiment and orthogonal experiment. Finally, the addition combination of rare earth elements is determined, which has improved the bacterial vitality, shortened the lag period and extended the peak period of acid production. The results show that adding 80 mg/L La3+, 2 mg/L Ce3+ and 0.3 mg/L Nd3+ into L-tyrosine fermentation medium is beneficial to fermentation. Fermentation time is 35 h, which is shortened by 2.2%. The maximum bacterial biomass reaches 80.3, increasing by 10.8%. Acid yield reaches 55.7 g/L, increasing by 9.7%. The sugar-acid conversion rate is 23.4%, increasing by 6.4%. The feasibility of adding rare earth elements is verified, which has provided a basis for large-scale industrial production of L-tyrosine.
Key words: Escherichia coli; L-tyrosine; rare earth element; sugar consumption rate
L-酪氨酸是一種對人體起著重要作用的芳香族氨基酸[1],屬于三大芳香族氨基酸之一,廣泛應用于化工、食品、醫藥、飼料等行業[2]。L-酪氨酸有助于加快新陳代謝,刺激人體神經系統,輔助治療慢性疾病。除此之外,L-酪氨酸可以作為一種重要的大腦化學物質來幫助調節人體的情緒、食欲以及人體對外界壓力的應激反應等[3],L-酪氨酸還是多種醫藥化工產品的制備原料,在農業中作為肥料增加農作物的抗旱性,在化妝品行業中起到美白和調理皮膚的作用[4]。
稀土元素被發現于18世紀末,包括銅、鈷、欽等17種元素,21世紀初有人開始對稀土元素的醫學作用進行研究[5]。我國是首個將稀土元素應用于農業的國家,并取得了良好的效果。馬文彬等[6]通過研究稀土在畜牧業養殖中的應用發現,稀土有促進動物分泌生長素的作用。除了動、植物外,早在20世紀末,有研究表明稀土元素對微生物的生長也起到一定的作用,王燕等[7]通過研究稀土元素對谷氨酸發酵及谷氨酸脫氫酶的作用,發現稀土元素能促進谷氨酸棒桿菌的生長和酶活力。也有報道表明,稀土配合物對黑曲霉、黃曲霉等真菌的生長有抑制作用。根據已有研究[8],少量稀土可以作為一種生理激活劑,起到激活生物體內生長因子的作用,有關稀土生物效應的研究越來越受到人們的關注。因此,探究稀土元素對大腸桿菌發酵生產L-酪氨酸的作用具有現實意義。
目前生產L-酪氨酸最主要的方法為微生物發酵法[9],該方法原材料廉價,且發酵成本低,原料易獲得,反應所需能耗小,對環境污染很小。在合適的發酵條件下,菌株經過發酵可直接獲得L-酪氨酸。大腸桿菌成為生產L-酪氨酸的主要微生物,主要是由于大腸桿菌具有生長速率快、易于培養的優點,是目前大規模工業化生產的理想菌株之一[10]。現已有眾多科研人員開展了微生物發酵法生產L-酪氨酸的研究[11],但發酵技術仍然不成熟。針對發酵過程中生物量低、生產速率低、糖酸轉化率低等問題,本研究以L-酪氨酸生產菌——大腸桿菌為供試菌株,探究稀土元素對該菌株的生長及其合成產物的影響,通過單因素實驗和正交實驗初步確定了稀土元素的最適添加量以及生產L-酪氨酸的最適培養基的組成成分。
因此,為了緩解L-酪氨酸發酵后期菌體活力下降的問題,本研究以La3+、Ce3+、Nd3+3種稀土元素為基礎,探究稀土元素對L-酪氨酸發酵的影響,并優化其用量,為L-酪氨酸及其他芳香族氨基酸的低成本、高效率工業化生產提供了一定的依據。
1 材料與方法
1.1 菌種
L-酪氨酸生產菌:大腸桿菌TYR-05(PT7-aroGfbr+PT7-tyrAfbrtyrB+PxylF-T7RNAP+ApheLA),天津科技大學代謝工程研究室保藏。
1.2 培養基
1.2.1 種子培養基
葡萄糖25 g/L,酵母粉4 g/L,檸檬酸鹽2 g/L,(NH4)2SO4·7H2O 2.5 g/L,KH2PO4·3H2O 3 g/L,MgSO4·7H2O 2 g/L,FeSO4·7H2O 5 mg/L,MnSO4·7H2O 1 mg/L,VH 0.5 mg/L,VB混0.5 mg/L,微量元素混合液1 mL/L。
1.2.2 發酵培養基
葡萄糖20 g/L,酵母粉4 g/L,(NH4)2SO4 3 g/L,KH2PO4·3H2O 1.5 g/L,檸檬酸鹽1.8 g/L,蛋氨酸1 g/L,苯丙氨酸0.8 g/L,FeSO4·7H2O 30 mg/L,MnSO4·H2O 10 mg/L,VB混0.5 mg/L,微量元素混合液1.5 mL/L(除對稀土元素添加方式進行探究外,其他成分均相同)。
1.3 主要儀器
Biotech-JG/JGY/BG發酵罐 上海保興生物設備工程有限公司;SBA-40ES生物傳感分析儀 濟南延和生物科技有限公司;BT-4電子天平 深圳博途科技有限公司;75UV分光光度計 上海菁華科技儀器有限公司;MiniSpin 離心機 德國Eppendorf 公司;Agilent 1200型高效液相色譜儀、ZW型色譜柱溫箱 安捷倫科技(中國)有限公司;pH 計、Direct-Q3超純水機 美國Mettler Toledo公司。
1.4 培養方法
1.4.1 菌種活化
從-80 ℃冰箱中取出菌株,在超凈臺中于酒精燈火焰旁用接種環取3環于試管斜面上,于32 ℃培養箱中培養12~14 h。
1.4.2 一級種子培養
待菌種活化好后,在超凈臺中于酒精燈火焰旁用接種環取3環于茄形瓶斜面培養基上,于32 ℃培養箱中過夜培養。
1.4.3 二級種子培養
待一級種子活化好后,用滅菌的細菌水洗菌株,在發酵罐補料口于酒精燈火焰旁倒入發酵罐中,于36 ℃、pH 7.0中性環境中培養。
1.4.4 發酵培養
當菌體量OD600 nm達到要求時,按相應要求的接種量放液,同時加入發酵培養基于發酵罐中,加入底糖,流加氨水來調節pH,使其維持在7.0~7.2,調節攪拌軸控制罐內溶氧,調節通風控制罐壓,使罐壓維持在0.04 MPa,發酵溫度36 ℃,發酵周期35 h。
1.5 稀土元素添加控制發酵實驗
在培養基成分和其他發酵條件不變的前提下,分別探究培養基中添加不同添加量的3種稀土元素對L-酪氨酸發酵的影響。
1.6 發酵檢測方法
1.6.1 發酵過程中pH的測定
采用Hamilton pH電極實時檢測,精密pH試紙輔助測定。
1.6.2 發酵過程中殘糖的測定
于罐中取樣,用離心機離心留上清液,將上清液稀釋100倍,用SBA-40ES生物傳感儀測定殘糖含量。
1.6.3 發酵過程中菌體量的測定
每2 h于發酵罐中取樣,吸取原樣稀釋相應倍數,用75UV分光光度計測定菌體量,菌體量可用吸光度OD600 nm表示,其計算方式為:
菌體量=OD600 nm×稀釋倍數。
1.6.4 發酵過程中L-酪氨酸的測定
發酵開始,每2 h取樣,發酵液用pH 10~12的NaOH溶液稀釋一定倍數后,經0.45 μm膜過濾。高效液相色譜測定:層析柱:Phenomenex C18 110 (250 mm×4.6 mm,5 μm);流動相1 mL/min 10%的乙腈;溫度30 ℃;檢測波長230 nm;進樣量20 μL。
2 結果與討論
2.1 不同La3+添加量對L-酪氨酸發酵的影響
在5 L的發酵罐中進行不同La3+添加量的分批發酵,在發酵過程中每2 h測定菌體量(OD600 nm)、L-酪氨酸產量和最終的糖酸轉化率,按照不同濃度配比設計單因素實驗,稀土元素鑭添加量分別為0,40,80,120,160 mg/L[12],從而確定最佳La3+添加量,通過測定發酵過程中L-酪氨酸產量、菌體量以及糖酸轉化率,明確不同La3+添加量對菌體生長與產酸的影響,結果見圖1。
由圖1可知,隨著La3+添加量的逐漸升高,菌體量和L-酪氨酸產量出現先升后降的趨勢。結果表明,當培養基中不添加La3+時,菌體量為68.1,L-酪氨酸產量為51.8 g/L。當La3+添加量為80 mg/L時,菌體量和L-酪氨酸產量分別為75.1和55.7 g/L,相比于空白組,二者分別提高了9.7%和7.1%。La3+添加量在0~80 mg/L時與菌體量和L-酪氨酸產量呈正相關,此時La3+添加量的升高影響了大腸桿菌的代謝能力,增強了菌體本身對培養基中營養物質的吸收,使菌體生長速率加快,最終菌體量得到提高。此時La3+作為一種生理激活劑使代謝流更多地流向產物L-酪氨酸的合成,L-酪氨酸的產量也得到提高。當La3+添加量大于80 mg/L時,L-酪氨酸的產量逐漸下降,這是由于發酵液中的La3+添加量過多,對菌體產生毒害作用。La3+也會直接影響大腸桿菌細胞壁的肽聚糖結構[13],抑制產物合成,且這種抑制作用會隨著La3+添加量的進一步提高而更加顯著。因此,培養基中添加80 mg/L的La3+對L-酪氨酸發酵提升最大。
綜合結果來看,一定濃度的稀土元素鑭會促進菌體生長,而濃度過高反而會起到抑制作用,因此發酵時La3+的最適添加量為80 mg/L。
由表1可知,La3+添加量在80 mg/L時糖酸轉化率最高,為23.4%,相比于空白組提高了8.5%,且不同La3+添加量所對應的耗糖量也是不同的,這是由于La3+添加量為80 mg/L時,La3+作為一種生理激活劑,起到激活生物體內生長因子的作用,增強菌體對培養基中無機氮源的吸收與利用,從而促進菌體生長[14],使菌體消耗的葡萄糖更多地用于代謝產物的合成。可以看出向培養基中添加La3+可以提高L-酪氨酸的生產效率,降低生產成本,從而實現大規模生產。所以,培養基中添加一定量的稀土元素鑭對L-酪氨酸發酵有利。
2.2 不同Ce3+添加量對L-酪氨酸發酵的影響
分別向發酵培養基中添加0,2,4,6,8 mg/L的Ce3+,以菌體量、L-酪氨酸產量以及糖酸轉化率為指標,探究不同Ce3+添加量對發酵的影響,結果見圖2。
由圖2可知,發酵結束時不添加Ce3+的菌體量為67.8,L-酪氨酸產量達到51.4 g/L;當Ce3+添加量為4 mg/L時為最佳添加量,菌體量和L-酪氨酸產量分別為69.3和54.6 g/L,相比于空白組分別提高了2.2%和5.9%。Ce3+添加量在0~4 mg/L時,隨著Ce3+添加量的升高,菌體量和L-酪氨酸產量也隨之增大。這是由于對數生長期的大腸桿菌細胞對Ce3+的敏感性較強,Ce3+可以增加大腸桿菌細胞膜的通透性[15],而且通透性增加的程度會隨著Ce3+添加量的增加而加劇。當Ce3+添加量大于4 mg/L時,菌體量和L-酪氨酸產量也隨之下降,這是因為Ce3+同時參與了阻礙細胞生理代謝相關的酶促反應,對菌體本身產生毒害作用,加速菌體的衰亡[15]。
在Ce3+添加量為4 mg/L時,L-酪氨酸產量高于傳統發酵方法,細胞膜通透性的增加會提高產物外排的能力,解除產物對關鍵酶的反饋抑制,L-酪氨酸得以積累。因此,在培養基中添加適量的Ce3+會增強菌株的產酸能力,進而提高發酵的糖酸轉化率。
由表2可知,沒有添加Ce3+的培養基糖酸轉化率僅為20.3%,而添加了Ce3+的實驗組表現較好,菌株達到了最佳的生長狀態,糖酸轉化率達到了22.9%,相對于空白組提升了11.4%,且在0~4 mg/L時糖酸轉化率隨著Ce3+添加量的升高而升高。
2.3 不同Nd3+添加量對L-酪氨酸發酵的影響
通過單因素實驗,在發酵中后期分別選擇0,0.3,0.6,0.9,1.2 mg/L的Nd3+進行L-酪氨酸發酵實驗,測定發酵結束后的菌體量、L-酪氨酸產量和糖酸轉化率,通過對比確定最適的Nd3+添加量,結果見圖3。
由圖3可知,空白組即不添加Nd3+的菌體量為73.4,L-酪氨酸產量為52.1 g/L;Nd3+添加量為0.6 mg/L時,菌體量和L-酪氨酸產量均達到最大,分別為75.1和54.3 g/L,相比于空白組分別提升了2.3%和4.1%。添加Nd3+后,菌體量和L-酪氨酸產量隨著Nd3+添加量的增加而增大,Nd3+能夠延長菌體的穩定期,提高菌體的活力,降低發酵后期菌體裂解的風險,同時能夠促進輔酶Q及細胞DNA和RNA的生物合成,并能調節細菌對維生素等生長因子的吸收與消耗,從而促進細菌的增殖[16]。而過多的Nd3+會與細菌DNA發生作用,使DNA斷裂,使其不能完成基本的代謝活性,菌體活力下降,降低L-酪氨酸的產量[17]。因此,選擇0.6 mg/L Nd3+為最適添加量。
由表3可知,Nd3+添加量為0.6 mg/L時糖酸轉化率最大,為22.5%,相比于不添加Nd3+提高了6.2%。隨著Nd3+添加量的增加,細胞膜的物質運輸得以加強,細菌對營養物質的吸收能力增強,糖酸轉化率也隨之增加。
2.4 正交實驗優化
通過上述單因素實驗,本研究確定了La3+、Ce3+、Nd3+的最佳添加量分別為80,4,0.6 mg/L。采用菌體量和L-酪氨酸產量為考核指標,以L9(34)為正交實驗表設計三因素三水平正交實驗(見表4),A為La3+添加量,B為Ce3+添加量,C為Nd3+添加量,探究發酵中的菌體量和L-酪氨酸產量情況。菌體量正交實驗結果見表5,方差分析見表6;L-酪氨酸產量正交實驗結果見表7,方差分析見表8。
結合菌體量的正交實驗結果和極差(R)分析可知,3種因素對于菌體量而言,影響順序為A>B>C,即La3+>Ce3+>Nd3+,通過比較3個因素的均值大小,即最大均值對應最佳添加量,得到最佳添加組合為A2B1C1,即La3+最佳添加量為80 mg/L,Ce3+最佳添加量為2 mg/L,Nd3+最佳添加量為0.3 mg/L。
綜合產量的正交實驗結果和極差(R)分析可知,3種因素對于產量而言,影響順序為C>A>B,即Nd3+>La3+>Ce3+;由均值結果可知,最佳添加量組合為A2B2C3,即La3+添加量為80 mg/L,Ce3+添加量為4 mg/L,Nd3+添加量為0.9 mg/L。
本研究通過對菌體量和L-酪氨酸產量的正交實驗結果分析可知,La3+和Ce3+添加量對發酵菌體量和L-酪氨酸產量均有極顯著影響,而Nd3+添加量僅對菌體量有顯著影響,為了達到既保證菌體量又促進L-酪氨酸生產的目的,選擇La3+添加量80 mg/L、Ce3+添加量2 mg/L、Nd3+添加量0.3 mg/L為稀土元素的最佳添加量。
2.5 最適稀土元素添加量與常規發酵的對比驗證
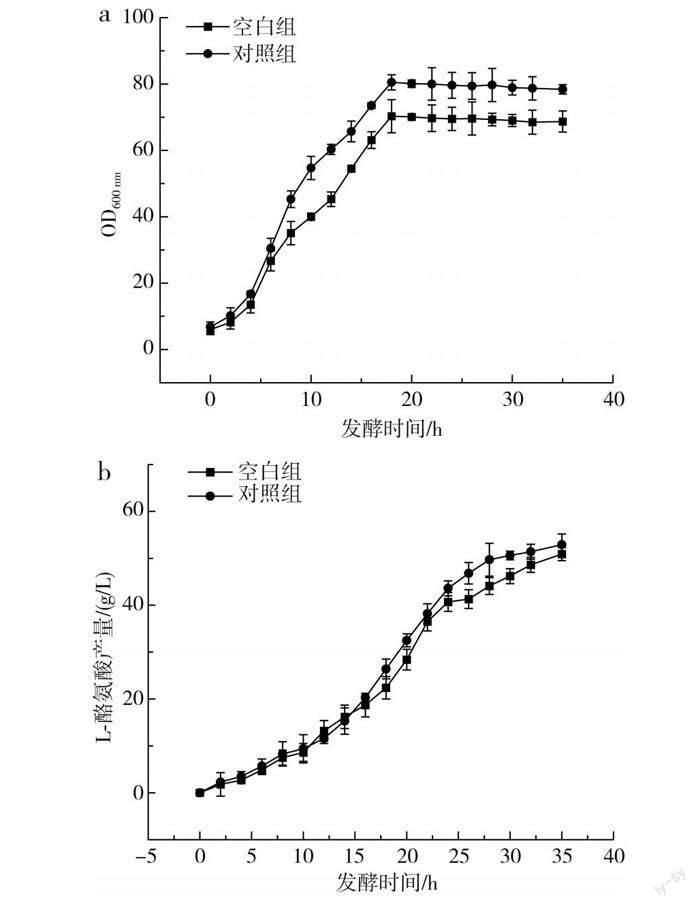
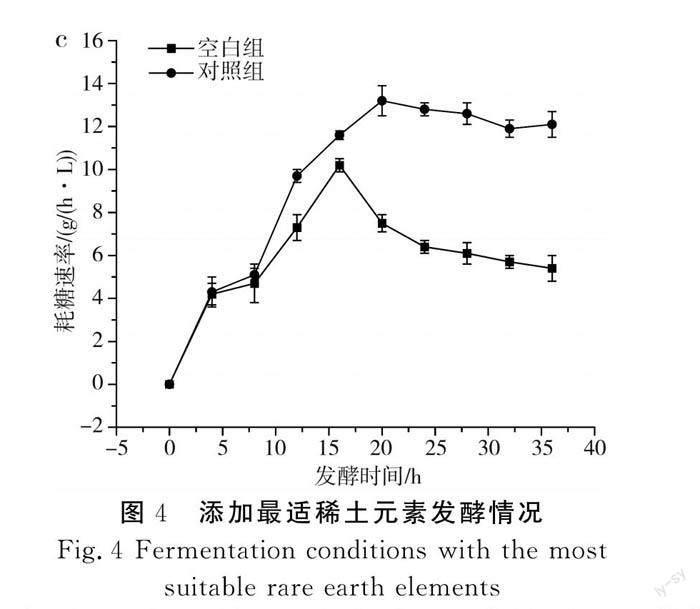
傳統的L-酪氨酸發酵過程中,發酵中期L-酪氨酸的產酸速率達到最大,但持續時間較短,發酵后期隨著時間的延長,L-酪氨酸的產酸速率出現快速下降的趨勢,分析原因可能是發酵后期菌體逐漸衰亡,對培養基中營養物質的吸收能力減弱。本研究通過單因素實驗確定了3種不同稀土元素的最優添加量,并通過正交實驗得到了最佳添加量組合,為了驗證實驗的可行性,在其他培養基條件一致的情況下,以不添加稀土元素的培養基作為對照組,以La3+、Ce3+、Nd3+分別添加80,2,0.3 mg/L為實驗組,進一步探究稀土元素及添加量的優化效果,結果見圖4。
由圖4可知,添加了最適稀土元素的對照組菌體量、產量和耗糖速率均大于空白組,其中對照組菌體量和產量分別達到75.6和52.9 g/L,相對于空白組提高了8.1%和3.8%;對照組在20 h時耗糖速率最大,為13.2 g/(h·L),而空白組最大耗糖速率出現在16 h時,為10.2 g/(h·L)。在對照組的發酵過程中,菌體生長旺盛,菌體活力高,耗糖速率大,產酸多,因此糖酸轉化率高于空白組。由此可見,稀土元素的添加提高了菌體本身對葡萄糖的利用率,解決了L-酪氨酸發酵后期菌體活力不足、對營養物質吸收能力弱的問題,為L-酪氨酸大規模生產提供了有利的依據。
2.6 不同稀土元素添加方式對L-酪氨酸發酵的影響
根據上述2.4正交實驗結果得出稀土元素添加量的最佳組合為La3+添加量80 mg/L、Ce3+添加量2 mg/L、Nd3+添加量0.3 mg/L。根據文獻[17]報道,發酵過程一次性補加稀土元素會對菌體產生抑制,為了解決這一問題,本實驗采用表9中3種發酵方式進行L-酪氨酸生產,并對其發酵過程進行監測,結果見圖5。
由圖5可知,采用傳統發酵方式生產L-酪氨酸時,最大菌體量為71.6,L-酪氨酸產量為50.3 g/L。采用一次性補加發酵方式的最大菌體量為76.5,L-酪氨酸產量為53.2 g/L,相比于傳統發酵分別提高了6.4%和5.5%。采用流加發酵方式的最大菌體量為80.3,L-酪氨酸產量為55.7 g/L,相比于傳統發酵分別提高了10.8%和9.7%。可能是在向發酵底物中加入稀土元素后,稀土元素能夠改變細胞膜的通透性,增強了細胞主動運輸或被動擴散的能力,加快了菌體對營養物質的吸收,提高了菌體自身的活力,加快了菌體生長,從而提高了L-酪氨酸的產酸速率和產酸量,但一次性補加稀土元素會在發酵初期菌體量較少時對菌體產生一定的抑制作用,延長菌體的延滯期,從而降低菌體的生長速率[18]。
由上述結果可知,采用流加稀土元素發酵的方式對L-酪氨酸發酵起到了促進作用,分析原因為加強了大腸桿菌的磷酸葡萄糖轉移酶系統,使菌體對葡萄糖的攝取能力提高;同時磷酸烯醇式丙酮酸合成酶是L-酪氨酸中心代謝途徑的關鍵酶[19],而稀土元素作為一種生理激活劑增加了此酶的活性,使葡萄糖更多地流向產物,從而促進了L-酪氨酸產量的提高。
3 結論
在大腸桿菌發酵法產L-酪氨酸的研究中,其產酸量與糖酸轉化率是大規模生產需要控制的重要指標,本研究通過單因素實驗分別確定了最佳La3+添加量為80 mg/L,最佳Ce3+添加量為4 mg/L,最佳Nd3+添加量為0.6 mg/L;通過正交實驗得出3種不同稀土元素的最佳組合為La3+80 mg/L、Ce3+2 mg/L、Nd3+0.3 mg/L;最終對稀土元素的添加方式進行優化,發現采用流加補料對提升L-酪氨酸發酵的菌體量、產酸量、糖酸轉化率影響最顯著,且采用發酵各項指標均達到最優。在此條件下,最大菌體量為80.3,與傳統發酵方式相比提高了10.8%;產酸量達到55.7 g/L,相比傳統發酵方式提高了9.7%;糖酸轉化率提高至23.4%。大腸桿菌發酵過程中容易被環境以及營養條件所影響,本研究通過實驗得知La3+能增強菌體對營養物質的攝取能力;Ce3+能增加細胞膜的通透性;Nd3+能延長菌體的穩定期,提高菌體的活力。通過調整發酵培養基成分,添加一定濃度的稀土元素,從而提高其產酸能力,對于大規模生產L-酪氨酸具有一定的參考價值。
參考文獻:
[1]BONGAERTS J, KRANER M, MULLER U, et al. Metabolic engineering for microbial production of aromatic amino acids and derived compounds[J].Engineering,2001,3(4):289-300.
[2]LUTKE-EVERSLOH T, STEPHANOPOULOS T G. L-tyrosine production by deregulated strains of? Escherichia coli[J].Applied Microbioly Biotechnology,2007,75(1):103-105.
[3]李國華,熊海波,陳寧,等.木糖對L-酪氨酸發酵的影響[J].中國調味品,2020,45(8):6-9,15.
[4]李國華.L-酪氨酸的發酵過程控制和提取工藝研究[D].天津:天津科技大學,2021.
[5]阮南,阮北,侯建革,等.稀土元素在谷氨酸發酵中的應用研究[J].中國稀土學報,2005(3):382-384.
[6]馬文彬,尹昕,朱秀雄,等.氯化釤對雌性大白鼠內分泌腺結構和功能的影響[J].中國稀土學報,1990(2):162-165.
[7]王燕,楊平平,宋香,等.稀土元素對谷氨酸發酵產酸及其谷氨酸脫氫酶的影響[J].食品與發酵工業,2004(9):33-36.
[8]紀云晶,栗建林.我國稀土某些生物學效應的研究概況[J].衛生毒理學雜志,2000(1):23-28.
[9]肖國安,王先兵,焦慶才,等.L-酪氨酸酶法轉化制備方法:中國,CN103224972A[P].2013-07-31.
[10]王欽.代謝工程改造大腸桿菌積累L-酪氨酸與發酵條件優化[D].無錫:江南大學,2019.
[11]姚元鋒,趙廣榮.L-酪氨酸代謝工程研究進展[J].食品與發酵工業,2013,39(5):132-137.
[12]張穎.稀土元素鑭對Corynebacterium pekinense發酵賴氨酸的影響[J].安徽農業科學,2012,40(10):5741-5742,5810.
[13]柴瑞娟,李露,王玉良.La和Ce對兩種細菌抗性的影響及其細胞壁的紅外光譜分析[J].稀土,2014,35(5):50-54.
[14]阮南,安家彥.稀土元素對檸檬酸發酵的影響[J].河北化工,2010,33(5):2-4.
[15]陳愛美,施慶珊,馮勁,等.Ce3+誘導大腸桿菌細胞膜通透性增加[C]//第八屆中國抗菌產業發展大會論文集,全國衛生產業企業管理協會抗菌產業分會,2012:149-153.
[16]袁素萍,高熙,米新宇,等.鏑對大腸桿菌微量元素吸收行為影響研究[J].現代儀器與醫療,2019,25(3):10-14.
[17]湯中勛.稀土元素對栓菌產漆酶的影響及其酶的固定化[D].蕪湖:安徽工程大學,2021.
[18]汪承潤,陳華波,楊帆,等.稀土鑭離子對大腸桿菌基因組DNA的影響[J].癌變·畸變·突變,2006(2):116-118.
[19]KIM S C, MIN B E, HWANG H G, et al. Pathway optimization by re-design of untranslated regions for L-tyrosine production in Escherichia coli[J].Scientific Reports,2015,5:13853.
收稿日期:2023-06-25
基金項目:寧夏回族自治區科技計劃項目(2021BDE92007);山東省重點研發計劃(2021ZDSYS10)
作者簡介:王碩(1999—),男,碩士研究生,研究方向:代謝控制發酵。
*通信作者:徐慶陽(1980—),男,副研究員,博士,研究方向:代謝控制發酵。
