小麥穗長QTL鑒定及其遺傳分析
2023-12-28 07:25:50姚琦馥周界光王健陳黃鑫楊瑤瑤劉倩閆磊王瑛周景忠崔鳳娟蔣云馬建
中國農業科學 2023年24期
關鍵詞:效應
姚琦馥,周界光,王健,陳黃鑫,楊瑤瑤,劉倩,閆磊,王瑛,周景忠,崔鳳娟,蔣云,馬建
小麥穗長QTL鑒定及其遺傳分析
姚琦馥1,周界光2,王健2,陳黃鑫2,楊瑤瑤2,劉倩2,閆磊2,王瑛2,周景忠3,崔鳳娟3,蔣云4,馬建2
1銅仁學院農林工程與規劃學院/貴州省梵凈山地區生物多樣性保護與利用重點實驗室,貴州銅仁 554300;2四川農業大學小麥研究所,成都 611130;3通遼市農牧科學研究所,內蒙古通遼 028015;4四川省農業科學院生物技術核技術研究所,成都 610000
【目的】穗長在決定小麥穗的構造和產量潛力方面具有重要作用。挖掘具有育種利用價值的小麥穗長數量性狀位點(quantitative trait loci,QTL),并解析其遺傳效應,為分子標記輔助育種提供理論依據。【方法】以自然突變體和川農16構建的198份F6代重組自交系(recombinant inbred lines,RIL)群體(MC群體)作為研究材料,于2020—2021和2021—2022年生長季,在四川溫江區、崇州市和雅安市(2021WJ、2022WJ、2021CZ、2022CZ和2021YA)進行試驗,對5個環境下的穗長進行表型鑒定。利用基于16K SNP芯片構建的高質量遺傳連鎖圖譜對穗長性狀位點進行定位。另外,根據穗長主效QTL側翼標記的基因型分析主效位點對產量相關性狀的遺傳效應,從而評估其對產量提升的潛力。【結果】共鑒定到14個控制穗長發育的QTL,主要分布在1A(1個)、1B(1個)、2B(1個)、3D(3個)、4A(1個)、4D(2個)、5A(1個)、5B(1個)、7A(1個)、7B(1個)和7D(1個)染色體。其中,在4個環境及最佳線性無偏預測(best linear unbiased prediction,BLUP)值中被檢測到,可解釋6.46%—20.12%的表型變異率,定位于1A染色體側翼標記—間,被視為主效QTL。的正效應位點來源于親本。在多環境QTL分析結果中也檢測到,表明其受環境影響較小,為主效且穩定表達的QTL。的效應在2個具有不同遺傳背景的驗證群體中得到進一步驗證。除旗葉長無顯著變化以外,攜帶正效應位點株系的每穗籽粒數(12.68%)、每穗粒重(14.99%)、千粒重(5.79%)、旗葉寬(2.94%)和小穗數(1.48%)顯著增加,花期(0.61%)顯著提前,而株高(-6.47%)和有效分蘗數(-36.11%)顯著減少。【結論】在1A染色體定位到1個主效且穩定的穗長位點。正效應位點顯著提高穗粒數、穗粒重、千粒重和小穗數,具有一定的育種價值。
小麥;16K SNP芯片;QTL;穗長;產量
0 引言
【研究意義】小麥作為主要的谷類作物之一,對全球的糧食安全有至關重要的作用[1]。全球可耕地面積的持續減少和人口的增加導致對小麥的需求劇增[2-4]。因此,提高小麥單產成為保障糧食安全、滿足人民谷物需求的主要措施。整個麥穗著生小麥的花序,所以穗作為小麥的繁殖器官,是小麥植株的重要組成部分[5-6],穗長是重要的穗部性狀,與單位面積穗粒數密切相關,對小麥產量提高有積極作用[7-9]。因此,挖掘穗長相關的基因/QTL,并研究穗長與農藝性狀的遺傳關系,可為小麥現代分子育種提供參考[10]。【前人研究進展】迄今為止,在小麥上已經鑒定到許多控制穗長的基因/QTL,Fan等[1]檢測到26個和穗長相關的QTL,其中,、和是在試驗環境中穩定的QTL,其中主效QTL和(478.65—541.29 Mb)分別可解釋7.99%— 16.58%和13.09%—30.43%的表型變異。Li等[2]用分別種植在5個不同生態環境中的雙親群體川麥42/科成麥1和川麥42/川農16進行QTL定位,共鑒定到34個穗密度和穗長共定位的QTL,其中,6個主效QTL至少在4個生態環境中被檢測到,BLUP值解釋7.13%—33.60%的表型變異。這些主效QTL均定位在5A和6A染色體,分別命名為和,這兩個QTL對株高、粒長和千粒重有一因多效的作用。Xu等[11]利用2個骨干親本碧螞4號/百農矮抗58構建的包含248個株系的RIL群體為材料,分別在染色體2D、3A、3B、3D、4A、4D、5A、5B、5D、7B和7D中檢測到18個穗長相關的QTL,可解釋2.04%—22.31%的表型變異。其中,2個穩定的主效QTL(13.25—36.89Mb)和506.65—524.73 Mb在4個生態環境中都可以檢測到,其增效位點來自于碧螞4號,其中可解釋11.03%—22.31%的表型變異,可解釋4.65%—8.38%的表型變異。陳黃鑫等[12]以矮蘭麥和野生二粒小麥LM001構建的F8代重組自交系(recombinant inbred lines,RIL)群體為材料,共檢測到17個與穗長相關的QTL,分布在2A、2B、3A、4A、4B、5A和6B染色體上,(—)在5個環境中被檢測到,可解釋10.41%—16.29%的表型變異,為穩定表達的主效位點,其加性效應來源于父本LM001。Ji等[13]以13F10和川麥42雜交構建的F8代RIL群體為材料,檢測到1個與穗長相關的主效QTL(516.60—521.27 Mb),可解釋7.88%—26.60%的表型變異,其早前研究表明5A染色體上控制穗長的主效位點也是春化作用基因位點。Xiong等[14]利用和LX987構建的包括400個單株的RIL群體為材料,在3A、4A、5B、6A、6B和7D染色體上共檢測到9個控制穗長的QTL,可解釋3.00%—22.00%的表型變異,其中染色體6A上的是穩定的主效QTL(—)。姚儉昕等[15]以小偃81和西農1376構建的包含120個株系的F9:10RIL群體為材料,共檢測到2個控制穗長的QTL(—)和(—),可分別解釋18.34%—22.51%和9.57%— 14.94%的表型變異,這兩個主效QTL的增效等位基因均來源于小偃81。其中在基因組上的物理距離為20.77—22.45 Mb,此位點和為同一個基因,而Chai等[16]以Y8679/J411構建的包含191個株系的RIL群體為材料,檢測到2個位于2D染色體與株高和穗長相關的主效QTL和,分別解釋30.94%—40.63%和31.31%—41.95%的表型變異。其增效位點都來自于親本J411。并證明此位點為同時控制株高和穗長的一致位點。Xu等[17]以人工合成異源六倍體小麥TAA10和重新合成異源六倍體小麥XX329雜交構建的包括198個單株的F7:8RIL群體為材料,共檢測到7個與穗長相關的QTL,其中5個是環境穩定的QTL,可解釋7.22%—11.59%的表型變異,其中穩定的主效QTL位點、和LOD值為6.19—12.89,可解釋21.77%—33.29%的表型變異,其增效位點來自于TAA10。和分別解釋14.75%和7.36%的表型變異,其增效位點來自于XX329。Ma等[18]以南大2419和望水白構建的包含136個株系的RIL群體株系間隨機交配構成的永久F2群體為材料,檢測到1個穗長相關的主效QTL(—),解釋29.70%— 36.30%的表型變異。該QTL在永久F2群體中也可解釋31.40%的表型變異。望水白在染色體區間的等位基因增加穗長、小穗數和不育小穗數,減少穗密度,通過選擇育種將會產生大穗的材料。水志杰等[19]以西農389×人工合成小麥材料KU98衍生的F7:8RIL群體為試驗材料,基于小麥55K SNP芯片對該群體進行基因分型,對小麥穗長和穗寬性狀進行了QTL定位,在1A、2D、3A、5A和7B染色體上共檢測到10個與穗長性狀相關的QTL。許多研究在不同染色體上鑒定出穗長穩定的QTL。Yao等[20]報道大約有350個穗長相關的QTL被鑒定,其中一些主效QTL分布在1D、2A、2B、2D、3A、3D、4A、4B、5A、5B、5D、6A、6B、6D、7A、7B和7D。【本研究切入點】小麥21條染色體上都有影響穗長的基因/QTL[1-2, 11-20]。但由于不同定位群體得出的結論有一定差異,且同時鑒定到的主效QTL數量很少,使得這些QTL在實際生產中的應用十分有限。因此,發掘優異的穗長基因并解析其遺傳機制,有利于小麥產量的提升。【擬解決的關鍵問題】本研究以穗長較長的自然變異株為母本、小麥品種川農16(CN16)為父本構建的F6代RIL群體MC為材料,利用基于16K SNP芯片技術構建的高密度遺傳圖譜對穗長性狀進行遺傳定位。結合產量相關性狀,分析穗長主效QTL對產量的潛在影響,為分子育種提供理論依據。
1 材料與方法
1.1 試驗材料
采用自然突變體和小麥品種CN16雜交構建的含有198個單株的F6代RIL群體MC為研究對象。具有多小花、長穗、多小穗等特點,是一個自然突變的優異材料。CN16小麥品種具有合適的株型。此外,2個具有不同遺傳背景的F2分離群體/20828(包含218個單株)和/蜀麥969(包含178個單株)作為驗證群體用于主效QTL效應的驗證。所有材料均由四川農業大學小麥研究所搜集并提供。
1.2 試驗設計
將MC群體及親本分別于2021、2022年種植于崇州(2021CZ和2022CZ)、溫江(2021WJ和2022WJ)和雅安(2021YA)共5個環境。采用單籽粒播種法,按照行長0.75 m、株距0.1 m、行間距0.3 m,將每個株系進行單行播種。種植MC群體的5個環境均進行2次重復試驗。/20828和/蜀麥969于2021年種植于崇州。這兩個驗證群體的種植方式與MC群體的相同。所有種植材料的田間管理根據當地氣候進行田間常規管理。
1.3 表型鑒定
當小麥生長至成熟期時,在5個環境對MC群體的穗長表型進行調查。單株主穗的基部到頂部(不包含芒)的長度為穗長。對每個株系選取至少4株長勢一致的單株進行表型測量,4個單株的平均值被認定為該株系的穗長。MC群體的株高、穗長、有效分蘗數、千粒重、開花期、每穗粒重、旗葉長、旗葉寬的最佳線性無偏預測(BLUP)值已被用于相關性分析[21-22],而本研究對調控穗長的遺傳位點進行鑒定和分析。此外,驗證群體/20828和/蜀麥969的穗長表型調查方式與MC群體的相同。
1.4 遺傳圖譜構建和QTL定位
利用Zhou等[22]通過16K SNP芯片構建的遺傳連鎖圖譜對控制穗長發育的位點進行遺傳定位。運用IciMapping 4.2軟件中完備區間作圖法對單個環境的QTL進行檢測,參數設置為Step=0.1 cM、PIN=0.001和LOD≥2.5。同時,運用IciMapping 4.2軟件對QTL進行多環境分析,參數設置為Step=0.1 cM、PIN=0.001和LOD≥5。按照國際遺傳命名規則(https://wheat.pw. usda.gov/ggpages/wgc/98/Intro.htm)對檢測到的QTL命名。利用WheatOmics v1.0公布的中國春參考基因組(CS RefSeq v2.1)獲得QTL側翼標記序列的物理位置。
1.5 主效QTL驗證
基于QTL的定位結果,將與主效QTL連鎖的SNP標記轉化為kompetitive allele specific PCR(KASP)標記。具體的轉化方法參考Zhou等[22]的描述。KASP標記包含2條特異引物(5′-GAAGGTGACCAAGTT CATGCTGCAACATGTATGTCCGACCTa-3′和5′-GA AGGTCGGAGTCAACGGATTGCAACATGTATGTCCGACCTg-3′)與1條通用引物(5′-GCAAATGGTGC GTTGATGGT-3′),KASP引物的合成由北京擎科生物科技有限公司完成。利用開發的KASP引物對/蜀麥969和/20828 F2分離群體的每個單株進行基因型分型。最后,對攜帶正效應位點和不攜帶正效應位點的純合單株的穗長進行統計分析,而基因型為雜合類型的單株不參與統計分析。
1.6 統計分析
穗長表型數據的多環境方差分析(analysis of variance,ANOVA)通過IciMapping 4.2軟件獲得。使用SAS 9.4軟件對穗長的BLUP值和廣義遺傳力(2)進行計算。用Excel 2019對穗長表型數據的平均值進行計算。運用Origin 2021軟件繪制穗長的頻率分布圖。利用IBM SPSS Statistics 20進行穗長表型數據的描述性統計分析、獨立樣本檢驗和顯著性分析。
2 結果
2.1 表型分析
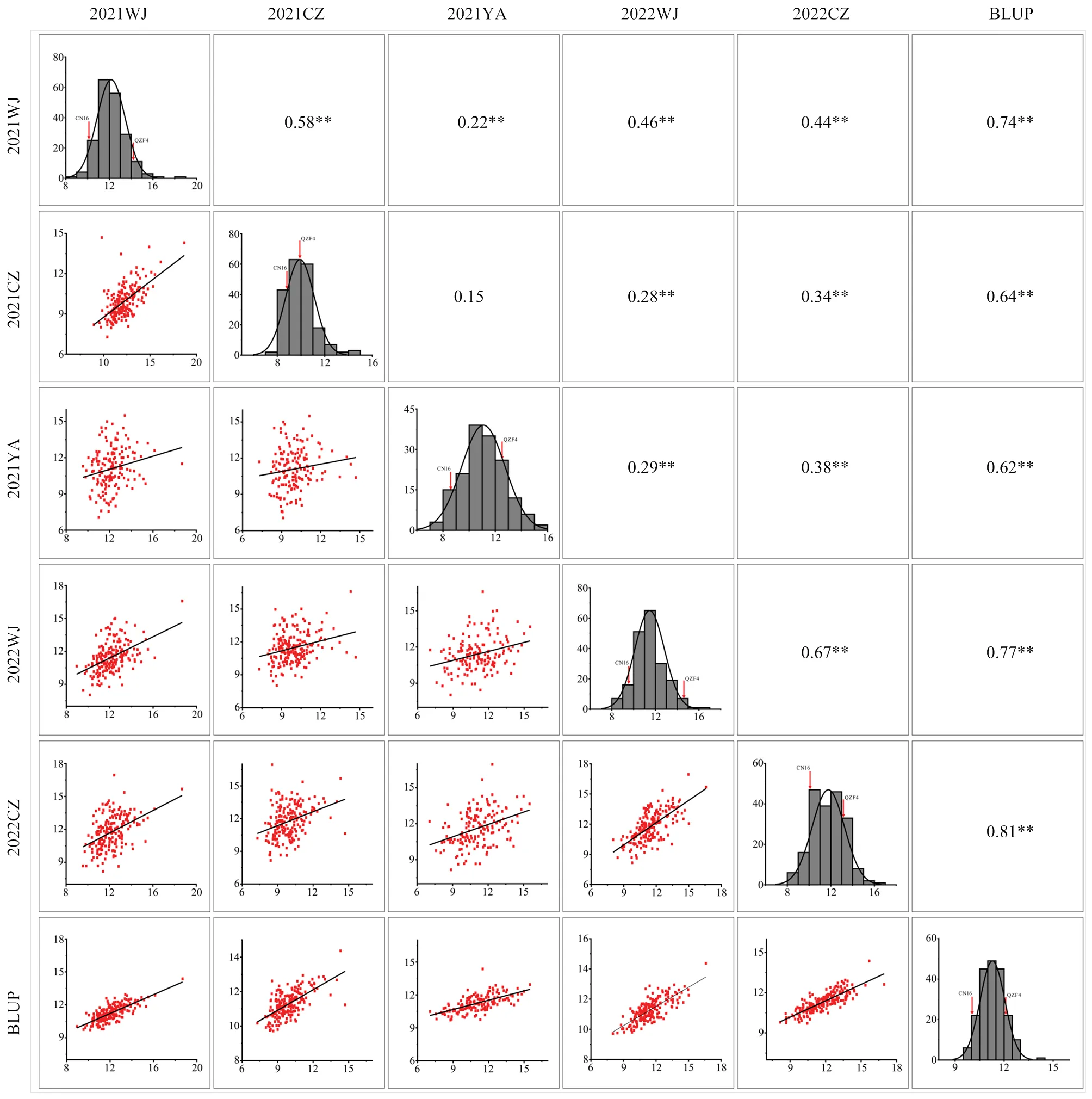
通過對5個種植環境下的MC群體穗長表型值進行鑒定。親本的穗長均顯著高于親本CN16(表1和圖1,<0.05)。MC群體穗長2為0.70,表明其主要受遺傳因子影響。RIL中穗長表型值范圍為7.05—18.68 cm,在不同環境中都存在超親分離現象,且其頻率分布呈近似正態分布(表1和圖2),表明MC群體穗長性狀具有典型的數量遺傳特點。ANOVA分析表明環境、基因型和環境?×?基因型互作對多環境的穗長有顯著影響(表2)。
2.2 相關性分析
根據BLUP值對MC群體在不同環境的穗長表型進行了相關性分析,其相關系數介于0.15—0.81(圖2)。除2021CZ和2021YA之間不存在顯著相關外,其余環境之間均表現出極顯著的正相關(<0.01)。

表1 MC群體親本及其RIL穗長的表型分布
WJ:溫江;CZ:崇州;YA:雅安;BLUP:最佳線性無偏預測;N:無重復值;*和**:在0.05和0.01水平差異顯著。下同
WJ: Wenjiang; CZ: Chongzhou; YA: Ya’an; BLUP: Phenotype values based on the best linear unbiased prediction;N: no duplicate values; * and **: Significant difference at level 0.05 and 0.01. the same as below

表2 MC群體穗長的方差分析

bar=2 cm
同時評估了MC群體穗長和其他產量相關性狀的相關性(表3)。結果顯示,穗長與小穗數、每穗籽粒數和每穗粒重之間存在極顯著的正相關(<0.01),相關系數分別為0.43、0.64和0.50。穗長和有效分蘗數之間存在極顯著的負相關(<0.01),相關系數為-0.45。穗長與株高、千粒重、旗葉長、旗葉寬和開花期之間無顯著相關性。
2.3 QTL分析
根據Zhou等[22]構建的遺傳連鎖圖譜和5個環境的穗長表型數據,共鑒定到14個控制穗長發育的QTL,主要分布在1A(1個)、1B(1個)、2B(1個)、3D(3個)、4A(1個)、4D(2個)、5A(1個)、5B(1個)、7A(1個)、7B(1個)和7D(1個)染色體(表4)。其中,在4個環境及BLUP值中被檢測到,可解釋6.46%—20.12%的表型變異率,定位于1A染色體側翼標記和之間,被視為主效QTL。的正效應位點來源于突變體親本。其余13個QTL均在2個環境及以下被檢測到,最高可解釋8.40%的表型變異率,都被視為微效QTL。多環境QTL分析結果中同樣檢測到,且可解釋表型變異率大于10%,表明其受環境影響較小,為主效且穩定的QTL(表5)。

表3 MC群體穗長與產量相關性狀的相關性
2.4 穗長主效QSl.sau.1A遺傳分析
根據穗長主效側翼標記的基因型,將MC群體分為攜帶增效位點的株系和不攜帶增效位點的株系,并對這2類株系進行分析(圖3)。結果顯示,除2021YA之外的所有環境中,攜帶正效應位點株系的穗長均極顯著高于沒有攜帶正效應位點的株系,最高能顯著提高11.53%的穗長(圖3)。
**:在0.01水平差異顯著。下同 **: Significant difference at level 0.01. the same as below
2.5 QSl.sau.1A位點的驗證
為了驗證的遺傳效應,將其連鎖的SNP標記轉化為KASP標記,序列見材料方法1.5部分。KASP引物在/20828和/蜀麥969群體中的基因型分型結果顯示,在/20828和/蜀麥969群體中,攜帶正效應位點的單株分別有56株和30株,不攜帶正效應位點的單株分別有55株和52株(圖4)。通過對這些單株所對應的穗長進行統計分析,發現在/20828和/蜀麥969群體中攜帶正效應位點的單株比不攜帶正效應位點的單株分別極顯著增加7.42%和9.99%的穗長(<0.01,圖4)。結果表明,的確是一個具有顯著增加穗長效應的主效QTL。
2.6 穗長主效QTL QSl.sau.1A對產量相關性狀的影響
進一步根據穗長主效QTL側翼標記的基因型分析其對產量相關性狀的影響。除旗葉長無顯著變化外,攜帶正效應位點株系的每穗籽粒數(12.68%)、每穗粒重(14.99%)、千粒重(5.79%)和旗葉寬(2.94%)極顯著提高,開花期(0.61%)極顯著提前,小穗數(1.48%)顯著增加,而株高(-6.47%)和有效分蘗數(-36.11%)極顯著減少(圖5)。暗示正效應位點對于產量可能有著積極的影響,具有較大的育種價值。

表4 MC群體穗長相關的QTL

Chr.:染色體;+和?:攜帶和不攜帶QSl.sau.1A正效應位點的株系;n:株系數。下同

表5 MC群體穗長相關的多環境QTL
LOD(A):加性和顯性效應的閾值;LOD(AbyE):環境對加性和顯性效應影響的閾值;(A):加性和顯性效應的表型變異率;(AbyE):環境對加性和顯性效應影響的表型變異率;--:與表4中的穗長QTL未對應上的QTL
LOD(A): Logarithm of the odds of additive and dominant effects; LOD(AbyE): Logarithm of the odds of the influence of environment on additive and dominant effects;(A): Phenotypic variation explained of additive and dominant effects;(AbyE): Phenotypic variation explained of environmental impact on additive and dominant effects; --: The QTL not corresponding to the spike length QTL in Table 4

+和?:攜帶和不攜帶對應QSl.sau.1A正效應位點的單株。下同
3 討論
3.1 與前人研究的比較分析
在CS RefSeq v2.1中檢索本研究和前人研究中穗長QTL的側翼標記序列,通過比對其物理位置來確定QTL是否重疊[23-24]。水志杰等[19]檢測到1個與穗長性狀相關的主效QTL,位于1A染色體標記區間—(51.96— 54.49 Mb)。Kumar等[8]在1A染色體短臂檢測到1 個與穗長性狀相關的位于標記區間—(23.57—67.92 Mb)。本研究鑒定到1個控制穗長發育的主效定位于1A染色體上的側翼標記—(1.21—11.43 Mb)。其物理區間無重疊,可能為新的QTL,由于不同材料間遺傳背景的差異,所以需對進行候選基因克隆來進一步確定其是否為新的穩定遺傳的主效QTL。

*:在0.05水平差異顯著;a—i:QSl.sau.1A對于其他農藝性狀的影響
3.2 穗長與產量相關性狀的關系
穗長是受多基因控制的小麥重要農藝性狀。與產量三要素畝穗數、穗粒數和粒重之間存在一定關系[25],深入研究小麥穗長性狀對提高小麥產量有重要意義[26-27]。此外,小麥為異源六倍體生物,基因組大,遺傳關系復雜,很難闡釋清楚小麥產量相關性狀的遺傳關系。增加穗長而未影響穗密度需提高可育穗粒數來增加產量。因此,在育種過程中要協調穗長與其他產量性狀間的關系,才能達到更大的收獲指數[28]。基于此,本研究分別用種植在5個生態環境的MC群體評價穗長和產量相關性狀間的相關性。正效應位點可顯著增加每穗籽粒數、每穗粒重、千粒重、小穗數和旗葉寬,促進開花期提前,顯著減少有效分蘗數和株高。李濤等[29]研究表明,當同時聚合和時,在川麥42×川農16 RIL群體中株高、穗長和千粒重分別增加13.10%、8.00%和7.50%,對穗粒數基本沒有影響;而在川麥42×川麥39 RIL群體中,株高和千粒重分別增加7.40%和4.70%,而穗長和穗粒數基本不受影響。CUI等[30]基于2個定位群體的條件QTL分析株高與其組成成分穗長和節間長之間的遺傳關系,結果表明,在QTL水平上,穗長對株高的貢獻最小。唐華蘋等[10]基于穗長的條件QTL結果,利用與穗長主效位點、和緊密連鎖的側翼標記基因型分析它們對其他農藝性狀的影響,表明對于株高、穗莖長、每穗小穗數和千粒重沒有顯著影響,對千粒重有顯著影響,對株高和穗莖長有極顯著影響。Ji等[13]研究發現穗長與籽粒長呈極顯著正相關(<0.01),與穗密度呈極顯著負相關(<0.01)。以上研究表明,穗長與產量性狀之間的關系在不同背景下表現并不一致,暗示正效應位點對于產量提高可能有著積極的影響,具有潛在的育種價值。為了能真正評價該位點的育種效應,后期應該設置小區試驗進一步評價是否對單位面積穗數、千粒重以及單位面積產量有實際效應,以明確其真實效應。
3.3 QSl.sau.1A區間內候選基因預測
基于CS RefSeq v2.1,的側翼標記1和的區間大小為10.22 Mb,區間內共包含414個基因,其中,高可信基因202個。根據202個高可信基因的功能注釋,預測了8個(、、、、、、、)可能影響穗長發育的基因。
4 結論
在1A染色體定位到1個主效且穩定地控制穗長發育的QTL,定位于側翼標記—(1.21—11.43 Mb)。正效應位點除對旗葉長無顯著影響外,可顯著增加每穗粒數、每穗粒重、千粒重、小穗數和旗葉寬,促進開花期提前,顯著減少有效分蘗數和株高。其中,的側翼標記和的區間大小為10.22 Mb,區間內預測了8個(、、、、、、、)可能影響穗長發育的基因。
[1] Fan X L, Chi F, Ji J, Zhang W, Zhao X Q, Liu J J, Meng D Y, Tong Y P, Wang T, Li J M. Dissection of pleiotropic QTL regions controlling wheat spike characteristics under different nitrogen treatments using traditional and conditional QTL mapping. Frontiers in Plant Science, 2019, 10: 187.
[2] Li T, Deng G B, Su Y, Yang Z, Tang Y Y, Wang J H, Qiu X B, Pu X, Li J, Liu Z H, Zhang H L, Liang J J, Yang W Y, Yu M Q, Wei Y M, Long H. Identifcation and validation of two major QTLs for spike compactness and length in bread wheat (L.) showing pleiotropic effects on yield-related traits. Theoretical and Applied Genetics, 2021, 134: 3625-3641.
[3] Butterworth M H, Semenov M A, Barnes A, Moran D, West J S, Fitt B D L. North-South divide: contrasting impacts of climate change on crop yields in Scotland and England. Journal of the Royal Society Interface, 2010, 7(42): 123-130.
[4] Su Z Q, Jin S J, Lu Y, Zhang G R, Chao S, Bai G H. Single nucleotide polymorphism tightly linked to a major QTL on chromosome 7A for both kernel length and kernel weight in wheat. Molecular Breeding, 2016, 36(2): 15.
[5] Faris J D, Zhang Z C, Garvin D F, Xu S S. Molecular and comparative mapping of genes governing spike compactness from wild emmer wheat. Molecular Genetics and Genomics, 2014, 289(4): 641-651.
[6] Koppolu R, Schnurbusch T. Developmental pathways for shaping spike inflorescence architecture in barley and wheat. Journal of Integrative Plant Biology, 2019, 61(3): 278-295.
[7] Jantasuriyarat C, Vales M I, Watson C J W, Riera- Lizarazu O. Identification and mapping of genetic loci affecting the free-threshing habit and spike compactness in wheat (L.). Theoretical and Applied Genetics, 2004, 108(2): 261-273.
[8] Kumar N, Kulwal P L, Balyan H S, Gupta P K. QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat. Molecular Breeding, 2007, 19(2): 163-177.
[9] Wu X Y, Cheng R R, Xue S L, Kong Z X, Wan H S, Li G Q, Huang Y L, Jia H Y, Jia J Z, Zhang L X, Ma Z Q. Precise mapping of a quantitative trait locus interval for spike length and grain weight in bread wheat (L.). Molecular Breeding,2014, 33(1): 129-138.
[10] 唐華蘋, 陳黃鑫, 李聰, 茍璐璐, 譚翠, 牟楊, 唐力為, 蘭秀錦, 魏育明, 馬建. 基于55K SNP芯片的普通小麥穗長非條件和條件QTL分析. 中國農業科學, 2022, 55(8): 1492-1502.doi: 10.3864/j. issn.0578-1752.2022.08.002.
Tang H P, Chen H X, Li C, Gou L L, Tan C, Mu Y, Tang L W, Lan X J, Wei Y M, Ma J. Unconditional and Conditional QTL analysis of wheat spike length in common wheat based on 55K SNP array. Scientia Agricultura Sinica, 2022, 55(8): 1492-1502. doi: 10. 3864/j.issn.0578-1752.2022.08.002.(in Chinese)
[11] Xu X, Li X J, Zhang D H , Zhao J S, Jiang X L, Sun H L, Ru Z G. Identification and validation of QTLs for kernel number per spike and spike length in two founder genotypes of wheat. BMC Plant Biology, 2022, 22(1): 146.
[12] 陳黃鑫, 李聰, 吳坤燕, 王岳, 牟楊, 唐華蘋, 唐力為, 蘭秀錦, 馬建. 四倍體小麥株高和穗長性狀的QTL定位及其遺傳效應分析. 麥類作物學報, 2022, 42(7): 799-807.
Chen H X, Li C, Wu K Y, Wang Y, Mu Y, Tang H P, Tang L W, Lan X J, Ma J. Detection of QTLs for plant height and spike length in tetraploid wheat and analysis of their genetic effect. Journal of Triticeae Crops, 2022, 42(7): 799-807. (in Chinese)
[13] Ji G S, Xu Z B, Fan X L, Zhou Q, Yu Q, Liu X F, Liao S M, Feng B, Wang T. Identification of a major and stable QTL on chromosome 5A confers spike length in wheat (L.). Molecular Breeding, 2021, 41(9): 56.
[14] Xiong H C, Li Y T, Guo H J, Xie Y D, Zhao L S, Gu J Y, Zhao S R, Ding Y P, Liu L X. Genetic mapping by integration of 55K SNP array and KASP markers reveals candidate genes for important agronomic traits in hexaploid wheat. Frontiers in Plant Science, 2021, 12: 628478.
[15] 姚儉昕, 張傳量, 宋曉朋, 許小宛, 邢永鋒, 呂棟云, 宋鵬博, 楊孟于, 孫道杰. 基于90K芯片的小麥穗長和旗葉長QTL分析. 麥類作物學報, 2020, 40(11): 1283-1289.
Yao J X, Zhang C L, Song X P, Xu X W, Xing Y F, Lü D Y, Song P B, Yang M Y, Sun D J. QTL analysis of wheat spike length and flag leaf length based on 90k SNP assay. Journal of Triticeae Crops, 2020, 40(11): 1283-1289. (in Chinese)
[16] Chai L L, Chen Z Y, Bian R L, Zhai H J, Cheng X J, Peng H R, Yao Y Y, Hu Z R, Xin M M, Guo W L, Sun Q X, Zhao A J, Ni Z F. Dissection of two quantitative trait loci with pleiotropic effects on plant height and spike length linked in coupling phase on the short arm of chromosome 2D of common wheat (L.). Theoretical and Applied Genetics, 2018, 131(12): 2621-2637.
[17] Xu H W, Zhang R Q, Wang M M, Li L H, Yan L, Wang Z, Zhu J, Chen X Y, Zhao A J, Su Z Q, Xing J W, Sun Q X, Ni Z F. Identification and characterization of QTL for spike morphological traits, plant height and heading date derived from the D genome of natural and resynthetic allohexaploid wheat. Theoretical and Applied Genetics, 2022, 135(2): 389-403.
[18] Ma Z Q, Zhao D M, Zhang C Q, Zhang Z Z, Xue S L, Lin F, Kong Z X, Tian D G, Luo Q Y. Molecular genetic analysis of five spike-related traits in wheat using RIL and immortalized F2populations. Molecular Genetics and Genomics, 2007, 277(1): 31-42.
[19] 水志杰, 安沛沛, 劉天相, 吳洪啟, 劉樂, 史雪, 王中華. 利用人工合成小麥RIL群體進行小麥穗長和穗寬性狀的QTL分析. 麥類作物學報, 2020, 40(6): 656-664.
Shui Z J, An P P, Liu T X, Wu H Q, Liu L, Shi X, Wang Z H. QTL analysis of spike length and width using RIL population of synthetic wheat. Journal of Triticeae Crops, 2020, 40(6): 656-664. (in Chinese)
[20] Yao H N, Xie Q, Xue S L, Luo J, Lu J K, Kong Z X, Wang Y P, Zhai W L, Lu N, Wei R, Yang Y, Han Y Z, Zhang Y, Jia H Y, Ma Z Q.on chromosome 7D of wheat (L.) regulates both head length and spikelet number. Theoretical and Applied Genetics, 2019, 132(6): 1789-1797.
[21] 姚琦馥, 陳黃鑫, 周界光, 馬瑞瑩, 鄧亮, 譚陳芯雨, 宋靖涵, 呂季娟, 馬建. 基于16K SNP芯片的小麥株高QTL鑒定及其遺傳分析. 中國農業科學, 2023, 56(12): 2237-2248.doi: 10.3864/j.issn.0578- 1752.2023.12.001.
Yao Q F, Chen H X, Zhou J G, Ma R Y, Deng L, Tan C X Y, Song J H, Lü J J, Ma J. QTL identification and genetic analysis of plant height in wheat based on 16K SNP array. Scientia Agricultura Sinica, 2023, 56(12): 2237-2248. doi: 10. 3864/j.issn.0578-1752.2023. 12.001.(in Chinese)
[22] Zhou J G, Li W, Yang Y Y, Xie X L, Liu J J, Liu Y L, Tang H P, Deng M, Xu Q, Jiang Q F, Chen G Y, Qi P F, Jiang Y F, Chen G D, He Y J, Ren Y P, Tang L W, Gou L L, Zheng Y L, Wei Y M, Ma J. A promising QTLlikely superior tofor the number of spikelets per spike of wheat shows no adverse effects on yield-related traits. Theoretical and Applied Genetics, 2023, 136(9): 181.
[23] INTERNATIONAL WHEAT GENOME SEQUENCING CONSORTIUM(IWGSC). Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science, 2018, 361(6403): eaar7191.
[24] Ma S W, Wang M, Wu J H, Guo W L, Chen Y M, Li G W, Wang Y P, Shi W M, Xia G M, Fu D L, Kang Z S, Ni F. WheatOmics: A platform combining multiple omics data to accelerate functional genomics studies in wheat. Molecular Plant, 2021, 14(12): 1965-1968.
[25] Wang M X, Lu J, Liu R, Li Y F, Ao D H, Wu Y, Zhang L. Identification and validation of a major quantitative trait locus for spike length and compactness in the wheat (L.) line chuanyu12D7. Frontiers in plant science, 2023, 14: 1186183.
[26] Li W L, Nelson J C, Chu C Y, Shi L H, Huang S H, Liu D J. Chromosomal locations and genetic relationships of tiller and spike characters in wheat. Euphytica, 2002, 125(3): 357-366.
[27] 王夢可, 趙德輝, 曾占奎, 陳鵬, 張雷宜, 蘭彩霞, 劉瑞芳, 王春平. 小麥穗長性狀基因的發掘與標記開發. 西北農林科技大學學報(自然科學版), 2023, 51(2): 11-21.
Wang M K, Zhao D H, Zeng Z K, Chen P, Zhang L Y, Lan C X, Liu R F, Wang C P. Gene detection and marker development of spike length traits in wheat. Journal of Northwest A&F University (Natural Science Edition), 2023, 51(2): 11-21. (in Chinese)
[28] Zhang X Y, Jia H Y, Li T A, Wu J Z, Nagarajan R, Lei L, Powers C, KAN C C, Hua W, Liu Z Y, ChEN C, Carver B F, Yan L L.modifies spike architecture and enhances grain yield in wheat. Science, 2022, 376(6589): 180-183.
[29] 李濤, 陸炳, 李俊, 鄧光兵, 張海莉, 梁俊俊, 余懋群, 楊武云, 龍海. 2個小麥株高QTL位點驗證及其對產量相關性狀的效應分析. 西南農業學報, 2019, 32(3): 476-483.
Li T, Lu B, Li J, Deng G B, Zhang H L, Liang J J, Yu M Q, Yang W Y, Long H. Validation of two plant height QTLs and their effects on yield-related traits in common wheat. Southwest China Journal of Agricultural Sciences, 2019, 32(3): 476-483. (in Chinese)
[30] Cui F, LI J, Ding A M, Zhao C H, Wang L, Wang X Q, Li S S, Bao Y G, Li X F, Feng D S, Kong L R, Wang H G. Conditional QTL mapping for plant height with respect to the length of the spike and internode in two mapping populations of wheat. Theoretical and Applied Genetics, 2011, 122(8): 1517-1536.
Identification and Genetic Analysis of QTL for Spike Length in Wheat

1College of Agroforestry Engineering and Planning, Tongren University/Guizhou Key Laboratory of Biodiversity Conservation and Utilization in the Fanjing Mountain Region, Tongren 554300, Guizhou;2Triticeae Research Institute, Sichuan Agricultural University, Chengdu 611130;3Tongliao Institute of Agriculture and Animal Husbandry Sciences, Tongliao 028015, Inner Mongolia;4Institute of Biotechnology and Nuclear Technology Research, Sichuan Academy of Agricultural Sciences, Chengdu 610000
【Objective】Spike length (SL) plays an important role in determining spike structure and yield potential of wheat. Quantitative trait loci (QTL) for spike length were excavated and their genetic effects were further analyzed to provide theoretical basis for molecular breeding.【Method】This study consisted of a population of 198 F6recombinant inbred lines (RIL) derived from the cross between the natural mutantand the cultivar Chuannong 16 (MC population). The MC population and its parents were planted in five different environments including Wenjiang in 2021 and 2022 (2021WJ and 2022WJ); Chongzhou in 2021 and 2022 (2021CZ and 2022CZ); and Ya’an in 2021 (2021YA) for spike length measurement. The 16K SNP chip-based constructed high-quality and high-density genetic linkage maps were used to map QTL for spike length. Additionally, the genotype of the flanking markers for the major spike length QTL was used to analyze its genetic effect on yield-related traits and thus to evaluate its potentiality for yield improvement.【Result】A total of 14 QTL for spike length were identified and they were mainly distributed on chromosomes 1A (one), 1B (one), 2B (one), 3D (three), 4A (one), 4D (two), 5A (one), 5B (one), 7A (one), 7B (one), and 7D (one). Among them,was detected in four environments and the best linear unbiased prediction (BLUP) value, explained 6.46% to 20.12% of the phenotypic variation, and thus was regarded as a major QTL. The positive allele atcame from the parental line. QTL analysis across multiple environments also detected, indicating it exhibits minimal environmental influence and represents a major and stably expressed QTL. The effect ofwas successfully verified in two populations with different genetic backgrounds. Genetic effects analysis showed that the positive allele ofshowed a significant effect on improving grain number per spike (12.68%), grain weight per spike (14.99%), 1000-grain weight (5.79%), flag leaf width (2.94%), spikelet number (1.48%), and flowering date (0.61%), and a significant effect of reducing plant height (-6.47%) and effective tiller number (-36.11%).【Conclusion】A major and stably expressed spike length QTL,, was detected on chromosome 1A. Its positive allele significantly increased grain number per spike, grain weight per spike, thousand grain weight, and spikelet number per spike, indicating its great breeding value.
wheat; 16K SNP array; QTL; spike length; yield

10.3864/j.issn.0578-1752.2023.24.002
2023-08-11;
2023-10-10
貴州省科技計劃基礎研究項目(黔科合基礎-ZK[2021]一般131)、銅仁市科技計劃基礎研究項目(銅市科研〔2023〕36號)
姚琦馥,E-mail:yaoqifu@126.com。周界光,E-mail:351062153@qq.com。姚琦馥和周界光為同等貢獻作者。通信作者馬建,E-mail:jianma@sicau.edu.cn。通信作者蔣云,E-mail:m13438880787@163.com
(責任編輯 李莉)
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
科學大眾(2020年17期)2020-10-27 02:49:10
紅土地(2018年11期)2018-12-19 05:10:56
意林·全彩Color(2018年9期)2018-11-13 22:49:38
中學物理·高中(2016年12期)2017-04-22 11:53:03
中國衛生(2016年4期)2016-11-12 13:24:14
中國衛生(2014年4期)2014-12-06 05:57:14
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
