杜仲葉際可培養真菌群落組成及產IAA菌株篩選
2024-01-01 00:00:00齊英華趙煜峰宋可云王艷玲冉青松邵秋雨董醇波韓燕峰
貴州大學學報(自然科學版) 2024年4期


摘要:杜仲作為我國特有、貴州廣泛分布的藥用植物,其葉際微生物對其生長具有重要意義。為揭示人工栽培和野生杜仲葉際真菌群落組成、多樣性及其功能,本研究采用傳統分離培養法,對杜仲葉附生及內生真菌群落組成及多樣性進行分析,基于FUNGuild數據庫對其進行功能注釋,并對其產IAA能力進行評估。本試驗共分離獲得真菌2門42屬226株。其中,從野生杜仲葉中分離獲得154株,主要包括青霉屬Penicillium(25.32%)等19個優勢屬;從人工栽培杜仲葉中分離獲得72株,主要包括曲霉屬Aspergillus(16.67%)等19個優勢屬。功能注釋結果顯示:除未定義功能類群,真菌類群主要涉及動物病原菌群(12.79%)、植物病原菌群-木腐菌群(13.95%)和內生菌群-附生菌群-植物病菌群(24.42%)。結果表明人工栽培種和野生種的真菌物種組成差別較大,且共有菌株比例較低。本研究還分離到多個產吲哚-3-乙酸(IAA)菌株。研究結果將為后續精準調控杜仲葉微生物組以增強植物健康、促進植物生長以及功能菌株的開發利用。
關鍵詞:植物微生物;葉際真菌;多樣性;生態功能;吲哚乙酸
中圖分類號:Q939文獻標志碼:A杜仲(Eucommia ulmoides Oliver)又名膠木,屬于杜仲科杜仲屬的落葉喬木植物,主要分布于中國甘肅、陜西等省份,是中國特有的藥用和經濟樹種,在《神農本草經》中被列為上品,富含綠原酸、黃酮、多糖等多種有效成分,通常使用其干燥的樹皮和葉子入藥,具有降壓、增強免疫力、降血糖等藥理作用[1]。有研究指出,杜仲的各組織中微生物物種多度高且數量龐大。這些微生物對杜仲的健康、藥用成分的合成和積累及產質量具有重要影響。楊娟等研究指出,杜仲樹皮中的某些藥理活性成分與真菌類群呈明顯的正相關[2]。DONG等則對杜仲樹皮中的細菌群落組成及功能進行了進一步探究,發現其中大部分核心細菌菌群與樹皮的藥理活性成分高度相關[3]。張青青等通過研究杜仲種子內的微生物群落組成和多樣性,發現杜仲種子內攜帶多種有益功能菌和潛在致病菌[4]。這些微生物對植物的生長發育和藥用價值具有重要影響。
葉片相關微生物,包括上下葉表面和葉組織內部的微生物被統稱為葉際微生物。由于葉片作為植物光合作用和物質生產的主要器官,葉際微生物能通過直接影響葉片功能和壽命從而在植物生產力和適應性方面發揮重要作用。但大多數杜仲微生物研究集中在根、樹皮和種子等組織部位,缺乏對葉際微生物的關注,少量相關研究則偏重內生菌中的細菌,而對附生菌和內生真菌缺乏了解,這阻礙了對葉際微生物群落的認識[5-7]。因此,本文以杜仲葉為研究對象,對其葉內生及附生真菌多樣性進行研究。
有研究指出,環境因素對葉際微生物群落結構具有重要影響[8]。魏亞情發現葉際真菌群落表現出顯著的生物地理格局,且葉際真菌群落的主要驅動因子包括年均溫、年均降水量等氣候因素[9]。另外,沉水植物葉際微生物的群落結構與周圍環境如溫度、光照等也有一定關系[10-11]。在年均溫、年均降水量、溫度、光照及土壤pH等環境因素方面,野生和人工種植條件存在較大差異,可能導致葉際微生物多樣性及豐度不同。因此,本研究對野生和人工種植杜仲葉際真菌群落進行了差異分析。
葉際微生物賦予宿主有益功能的重要方式為通過合成一種或多種植物激素影響植物激素水平。據報道,吲哚-3-乙酸(IAA)是影響杜仲葉際微生物群落的兩種關鍵植物激素之一,在調控其群落組成、多樣性和功能上發揮重要作用[12]。IAA是一種典型的植物生長促進激素,已被廣泛報道可由葉際微生物合成產生。如,潘明凱研究發現柑橘葉際微生物克雷伯氏菌Klebsiella表現出較強的IAA分泌能力[13]。梁淑雅等從苦草根、葉際附生細菌中篩選得到11株產IAA菌[14]。劉元等研究發現,懷山藥內生真菌銳頂鐮刀菌具有較強的IAA分泌能力,可能對宿主植物具有一定的促生作用[15]。葉際微生物,尤其是葉際真菌與植物的健康和生長密切相關,并且作為一種新興的微生物資源,在農業和醫藥等領域的生產應用方面具有巨大的發展空間[16]。
綜上所述,本研究通過傳統分離培養、形態鑒定以及分子鑒定等方法對杜仲葉附生及葉內生真菌群落組成及差異、多樣性進行探究,并對杜仲葉際真菌的生態功能進行注釋。同時,篩選出能夠產IAA的具有促生長活性的菌株,以期為今后杜仲的栽培管理提供重要信息,并積累具有開發利用價值的功能菌株資源。
1材料與方法
1.1樣品的采集與處理
從貴州大學西校區(26°25′N,106°40′E)選取4棵健康的野生杜仲樹和4棵健康的人工種植杜仲植株,剪取健康無病斑的新鮮葉片若干,編號后置于無菌保鮮袋中,放入4 ℃冰箱保存備用。
稱取采集的野生和人工種植的杜仲葉片各2 g分別置于燒杯中,依次用無菌蒸餾水對杜仲葉片進行沖洗,除去葉片表面的塵土顆粒,備用。
本研究各試驗樣本為野生杜仲葉附生菌株樣本(WP)、野生杜仲葉內生菌株樣本(WE)、人工栽培杜仲葉附生菌株樣本(PP)、人工栽培杜仲葉內生菌株樣本(PE)。
1.2葉附生和內生真菌的分離、純化及保種
1.2.1葉附生真菌的分離
葉面附生微生物的分離采用平板稀釋法[17]。將處理好的2 g葉片,放入盛有50 mL滅菌水的三角瓶中,放入振蕩器上,恒溫25 ℃下,以130 r/min振蕩30 min,即制成菌懸液母液,然后用無菌水稀釋成10-1、10-2梯度的菌懸液,分別取10-1、10-2兩個梯度的菌懸液200 μL均勻涂布于兩種培養基平板,每個梯度重復三次,然后置于25~28 ℃培養箱中培養3~5天。
1.2.2葉內生真菌的分離
將葉片組織從上述三角瓶中取出,在無菌操作臺上,用70%酒精處理2~3 s,然后用3.5%的次氯酸鈉溶液處理3~4 min,再用無菌水漂洗4~5次,每次處理1 min以上,然后用滅菌濾紙吸干組織表面水分。將最后一次洗滌液按100 μL/皿涂布于各分離培養基平板,并置于恒溫培養箱中,數天后若未發現培養皿中有菌落生長,則證明葉面滅菌操作合格,滅菌徹底,若皿中有菌落出現,則重復滅菌操作[17]。
內生微生物的分離采用勻漿平板稀釋分離法[18]。將表面消毒的葉片組織用無菌剪刀剪碎,避免用手接觸葉表,放入滅好菌的研缽中,加入無菌水定容至10 mL,研磨至漿狀,得到10-1稀釋倍數的樣品懸浮液,繼而將樣品懸浮液稀釋成10-2倍的稀釋液,由此得到100、10-1、10-2倍三個梯度的稀釋液,然后從每個梯度的稀釋液中各取200 μL涂布于備好的馬鈴薯葡萄糖培養基(PDA)平皿上,各梯度均設置三個平行,置于25~28 ℃培養箱中培養5~7天。
1.2.3菌株的純化及保種
待菌絲長出后,用接種針挑取適量菌絲轉接到新的PDA平板上,獲得純培養菌株。待菌株完成逐步純化后將其接入裝有30%甘油的凍存管中,放于4 ℃冰箱臨時保藏,或-80 ℃超低溫冰箱中進行長期保藏[19]。
1.3菌株的形態學鑒定
將分離得到的真菌菌株接種在PDA平板上,在25 ℃恒溫箱培養7~14天后,觀察、記錄其菌落特征,如顏色、形態、質地,并測量菌落直徑[20]。制片后在普通光學顯微鏡下觀察其菌絲形態,測量并記錄其孢子形狀、大小及分生孢子梗的形態和排列方式等。根據記錄的數據,查閱真菌學專業文獻資料,對比菌種特征,將菌株鑒定到屬。
1.4菌株的分子鑒定
用滅菌刀片從PDA平板上取少量真菌菌株的菌絲和孢子于離心管中,并加適量研磨珠于細胞破碎儀進行研磨。按照真菌基因組DNA提取試劑盒(北京百泰克生物技術有限公司)的操作說明進行真菌DNA的提取,備用。PCR擴增所需引物均來自上海生工生物術有限公司,分別為ITS1(5’-TCCGTAGGTGAACCTGCGG-3’)和ITS4(5’-TCCTCCGCTTATTGATATGC-3’)。
PCR反應體系為25 μL:Master Mix 12.5 μL,引物ITS1和引物ITS4分別為1 μL,DNA模板2 μL,ddH2O 8.5 μL。
PCR反應條件:94 ℃預變性 5 min,1個循環;94 ℃變性1 min;50 ℃退火1 min,72 ℃延伸1 min;進行35個循環,最后72 ℃延伸10 min。
PCR產物的檢測:以DNA Marker DL2000為大小參照,分別將擴增產物與對照放入Parafilm薄膜和上樣緩沖液中進行混合,在加入GenRed的0.8%的瓊脂糖凝膠上進行點樣,然后電泳40 min。在紫外光下,若熒光染料標記的DNA條帶清晰,且與預期大小一致,則說明擴增成功。
DNA測序:將擴增產物送至昆明碩擎生物科技有限公司,并對待鑒定菌株的ITS序列進行序列測定,經BioEdit人工校對去除引物及雙峰序列。登陸NCBI進行BLAST,在GenBank核酸序列數據庫中進行同源序列搜索,BLAST結果相似度為99%以上的,確定為同一種,結合形態學特征進行種屬鑒定[21-22]。
1.5產IAA菌株的篩選
將待測菌株接種到150 mL PDB液體培養基(馬鈴薯200 g,葡萄糖20 g,加蒸餾水至1 L,自然 pH,121 ℃ 滅菌 20 min)上進行培養(250 mL 錐形瓶,28 ℃,150 r/min)7 d,用移液槍吸取200 μL發酵液置白色坩堝內。1∶1加入標準比色液,每株菌試驗設置3個平行;同時設置一組空白對照(未接種的PDB培養液)和一組陽性對照(按1∶1將200 μL吲哚-3-乙酸標準液(0.9 mg /L)與標準比色液混合),在暗處靜置30 min后,觀察其顏色變化。顏色不變為陰性,代表菌株不產IAA;顏色變成粉紅或紅色則為陽性,代表菌株產IAA。
1.6數據處理及分析
1.6.1葉際真菌的分離頻率
本研究以分離頻率(IF)界定可培養真菌群落的優勢類群,將分離頻率在10%以上的屬定義為優勢屬。分離頻率的計算公式為[23]
分離頻率IF(%)=P/N×100%
式中:P為某一真菌分離株的總數;N為所有真菌分離株的總數。
1.6.2葉際真菌的群落組成與差異分析及生態功能注釋統計分離得到的真菌在屬水平上的分離頻率,對其進行群落組成及差異分析。通過FUNGuilds數據庫獲得已鑒定的真菌類群的營養型及生態功能信息。通過Omicstudio、BioLadder-生物信息在線分析和派森諾基因云繪圖平臺分別繪制杜仲葉際真菌群落生態功能和組成的百分比柱狀圖、主成分分析圖、比例韋恩圖。
1.6.3葉際真菌的α多樣性分析
本研究結合 Shannon-wiener與 Margalef 指數對各樣本進行多樣性分析,涉及的指數計算方法如下:
Shannon-wiener 多樣性指數H=-Σ(Pi×ln(Pi))
Margalef 豐富度指數R=(S-1)/ln N
式中:Pi為第i種真菌菌株數與所有真菌分離株總數的比值;S為分離得到的真菌種類數;N為所有真菌分離株的總數。
2結果與分析
2.1野生和人工栽培杜仲葉際真菌群落組成及差異分析本研究以野生和人工栽培杜仲葉為樣本,分離得到可培養真菌2門42屬75種226株。不同來源的杜仲葉真菌群落的優勢類群差異顯著且各不相同。
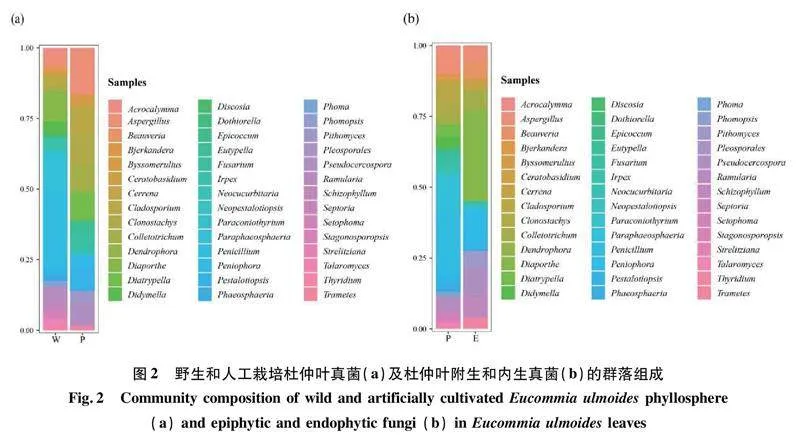
野生杜仲葉樣本共分離獲得附生真菌(WP)2門4綱11目18科25屬35種119株。其中,子囊菌門Ascomycota是主要的優勢類群。綱水平上,座囊菌綱Dothideomycetes、散囊菌綱Eurotiomycetes是優勢綱,分離頻率分別為50.42%和37.82%。其次是傘菌綱Agaricomycetes(8.40%)和糞殼菌綱Sordariomycetes(3.36%)。屬水平上,青霉屬Penicillium(31.93%)是優勢屬。其次是近暗球腔菌屬Paraphaeosphaeria(17.65%)、長隔孢屬Ramularia(6.72%)、亞隔孢殼屬Didymella(5.88%)、枝孢菌屬Cladosporium(5.04%)、耙齒菌屬Irpex(5.04%)、棘殼孢屬Setophoma(3.36%)、Acrocalymma(2.52%)、球腔菌屬Phaeosphaeria(2.52%)、曲霉屬Aspergillus(2.52%)和籃狀菌屬Talaromyces(2.52%)等。其余屬的分離頻率都在1%左右(圖1)。
野生杜仲葉樣本共分離獲得內生真菌(WE)2門3綱8目9科10屬15種35株其中,子囊菌門為真菌優勢類群。綱水平上,糞殼菌綱(68.57%)和傘菌綱(20.00%)為優勢綱。其次是散囊菌綱(11.43%)。屬水平上,其優勢屬是間座殼屬Diaporthe(45.71%),其次是裂褶菌屬Schizophyllum(8.57%)、炭疽菌屬Colletotrichum(8.57%)、白僵菌屬Beauveria(8.57%)、曲霉屬(8.57%)、隔孢伏革屬Peniophora(5.71%)、擬盤多毛孢屬Pestalotiopsis(5.71%)、下皮黑孔菌屬Cerrena(2.86%)、栓菌屬Trametes(2.86%)和青霉屬(2.86%)(圖1)。
人工栽培杜仲葉樣本共分離獲得附生真菌(PP)2門4綱7目9科11屬15種56株。其中,子囊菌門為主要優勢菌群。綱水平上,糞殼菌綱(60.71%)和散囊菌綱(21.43%)是優勢綱。其次是座囊菌綱(14.29%)和傘菌綱(3.57%)。屬水平上,其優勢屬是曲霉屬,分離頻率為21.43%,其次是螺旋聚孢霉屬Clonostachys(14.29%)、假蕉孢殼屬Diatrypella(12.50%)、枝孢菌屬(12.50%)、擬盤多毛孢屬(10.71%)、炭疽菌屬(10.71%)、鐮刀菌屬Fusarium(7.14%)、絮干朽菌屬Byssomerulius(3.57%)、彎孢聚殼屬Eutypella(3.57%)、新擬盤多毛孢屬Neopestalotiopsis(1.79%)和附球霉屬Epicoccum(1.79%)(圖1)。
人工栽培杜仲葉樣本共分離獲得內生真菌(PE)2門2綱7目8科8屬10種16株。其中,子囊菌門和擔子菌門Basidiomycota為優勢菌群。綱水平上,傘菌綱和座囊菌綱,分離頻率均為50.00%,是優勢綱。屬水平上,假尾孢菌屬Pseudocercospora(31.25%)為優勢屬。其次是隔孢伏革菌屬(18.75%)、皮司霉菌屬Pithomyces(18.75%)、角擔菌屬Ceratobasidium(6.25%)、裂褶菌屬(6.25%)、栓菌屬(6.25%)和耙齒菌屬(6.25%)等(圖1)。
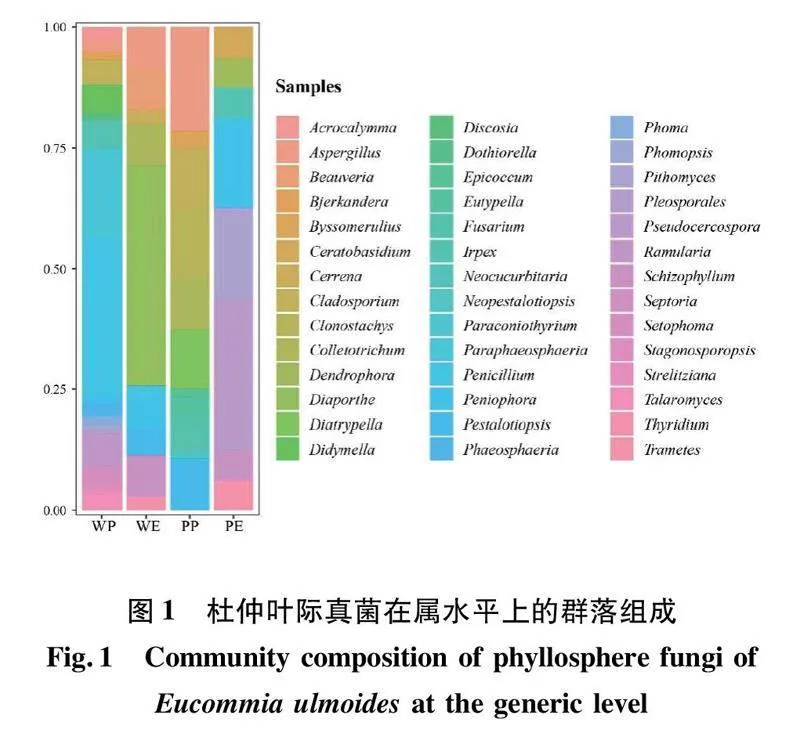
野生杜仲葉(W)共分離出真菌154株,隸屬于2門4綱14目24科31屬。在屬水平,如圖2(a)所示,主要包括青霉屬、近暗球腔菌屬、間座殼屬、長隔孢屬和亞隔孢殼屬等19個優勢屬,其中,青霉屬(25.32%)、近暗球腔菌屬(13.64%)和間座殼屬(10.39%)占有絕對優勢,為主要優勢屬。
人工栽培杜仲葉(P)共分離出真菌72株,隸屬于2門4綱12目16科19屬。在屬水平,如圖2(a)所示,主要包括曲霉屬(16.67%)和螺旋聚孢霉屬(11.11%)等19個優勢屬。
野生和人工栽培杜仲葉共分離到葉附生真菌(P)175株,隸屬于2門4綱13目22科33屬。在屬水平,如圖2(b)所示,主要包括青霉屬(21.71%)、近暗球腔菌屬(12.00%)和曲霉屬(8.57%)等21個優勢屬。
野生和人工栽培杜仲葉共分離到內生真菌(E)51株,隸屬于2門4綱12目14科15屬。在屬水平,如圖2(b)所示,主要包括間座殼屬(31.37%)、隔孢伏革菌屬(9.80%)和假尾孢菌屬(9.80%)等15個優勢屬,其中,間座殼屬(26.07%)為主要優勢屬。
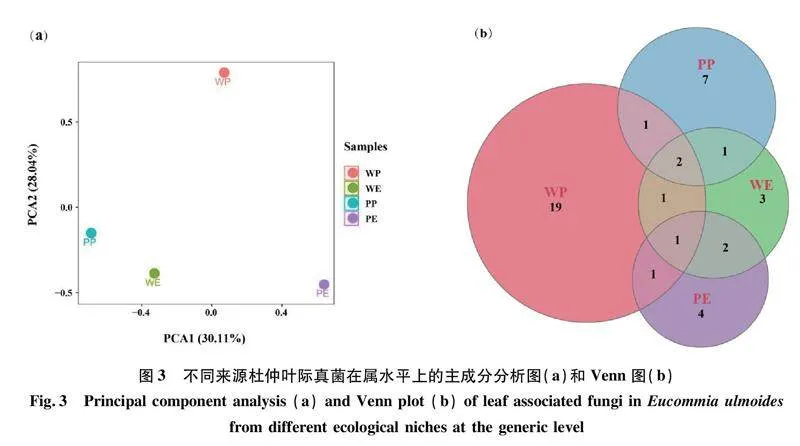
由主成分分析圖3(a)可知,第一主成分(PCA1)的方差貢獻率為30.11%,是主要特征,第二主成分(PCA2)的方差貢獻率為28.04%,是次要特征,累計方差貢獻率為58.15%。WP、WE、PP和PE這4個樣本在兩個主成分軸上呈現為4組,說明4樣本之間真菌群落具有顯著差異。
Venn圖顯示如圖3(b),野生和人工栽培杜仲葉際及內生可培養真菌群落不具有共有屬,互相之間差異明顯。不同生境不同區位杜仲葉樣本都存在一定數量的獨有真菌類群。例如,野生杜仲葉樣本中,內生(WE)真菌群落有3個獨有類群,依次為白僵菌屬、下皮黑孔菌屬和間座殼屬。人工栽培杜仲葉內生(PE)真菌群落具4個獨有類群,依次為角擔菌屬、Dendrophora、皮司霉菌屬和假尾孢菌屬。人工栽培杜仲葉附生(PP)真菌群落具7個獨有類群,依次為絮干朽菌屬、螺旋聚孢霉屬、假蕉孢殼屬、附球霉屬、彎孢聚殼屬、鐮刀菌屬和新擬盤多毛孢屬。野生杜仲葉附生(WP)真菌群落獨有類群最為豐富,多達19個屬,分別為:Acrocalymma、黑管菌屬Bjerkandera、亞隔孢殼屬、雙毛殼孢屬Discosia、小穴殼菌屬Dothiorella、新葫蘆科菌屬Neocucurbitaria、擬盾殼霉屬Paraconiothyrium、近暗球腔菌屬、球腔菌屬、莖點霉屬Phoma、擬莖點霉屬Phomopsis、Pleosporales、長隔孢屬、Septoria、棘殼孢屬、殼多孢菌屬Stagonosporopsis、橫斷孢屬Strelitziana、籃狀菌屬和Thyridium。
2.2野生和人工栽培杜仲葉真菌群落多樣性
微生物群落的多樣性指數是用來量化和比較不同樣本中微生物種類數量和豐度的統計量。Shannon-wiener 指數(香農-維納指數)是群落多樣性指數,它綜合考量群落中物種的豐富度(物種數量)和均勻度(每個物種的相對豐度)。它的數值越大,表示群落生物多樣性越高[24]。Magalef豐富度指數不考慮相對豐度,僅關注群落的豐富度,因此指數值越高,表示觀察到的物種越多,群落的物種豐富度也就越高[25]。
如表1所示,Shannon-wiener和 Margalef 指數的順序均為由高到低,表現為野生附生(WP)gt;人工附生(PP)≥野生內生(WE)gt;人工內生(PE)。這一結果表明,野生杜仲葉附生(WP)真菌群落最為豐富,其次是人工栽培杜仲葉附生(PP)和野生杜仲葉內生(WE)真菌群落,多樣性最低的是人工栽培杜仲葉內生(PE)真菌群落。結果還表明,野生(W)較人工栽培(P)杜仲葉真菌群落更為豐富;杜仲葉附生(P)較內生(E)真菌群落更為豐富。
2.3杜仲葉際真菌群落功能分析
使用FUNGuilds平臺對杜仲葉際真菌類群的營養型和生態功能類群進行分析(圖4)。其中,20個物種被成功注釋,營養型包括病原型(Pathotroph)(24.42%)、病原-腐生型(Pathotroph-Saprotroph)(24.42%)、共生型(Symbiotroph)(12.79%)、病原-共生型(Pathotroph-Symbiotroph)(38.37%)4大類。除未定義功能類群(Undefined functional group),所有真菌類群涉及植物病原菌群(Plant Pathogen)(11.63%)、動物病原菌群(Animal Pathogen)(12.79%)、內生菌群(Endophyte)(12.79%)以及多種混合功能類群如動物病原菌群-未定義腐生菌群(Animal Pathogen-Undefined Saprotroph)(3.49%)、內生菌群-附生菌群-植物病菌群(Endophyte-Epiphyte-Plant Pathogen)(24.42%)、動物病原菌群-內生菌群-木腐菌群(Animal Pathogen-Endophyte-Wood Saprotroph)(3.49%)、動物病原菌群-植物病原菌群-木腐菌群(Animal Pathogen-Plant Pathogen-Wood Saprotroph)(3.49%)、內生菌群-地衣寄生菌群-植物病原菌群(Endophyte-Lichen Parasite-Plant Pathogen)(1.16%)、植物病原菌群-木腐菌群(Plant Pathogen-Wood Saprotroph)(13.95%)、內生菌群-植物病原菌群(Endophyte-Plant Pathogen)(12.79%)10類。55個物種在FUNGuilds數據庫中未能注釋到對應功能。整體來看,野生杜仲葉際真菌功能群主要集中于內生菌群-附生菌群-植物病原菌群、內生菌群-植物病原菌群、動物病原菌群,人工栽培杜仲葉際真菌功能群則主要為植物病原菌群-木腐菌群和動物病原菌群,且在野生杜仲葉上的內生真菌數量顯著高于人工栽培杜仲。
2.4產IAA菌株篩選及能力評估
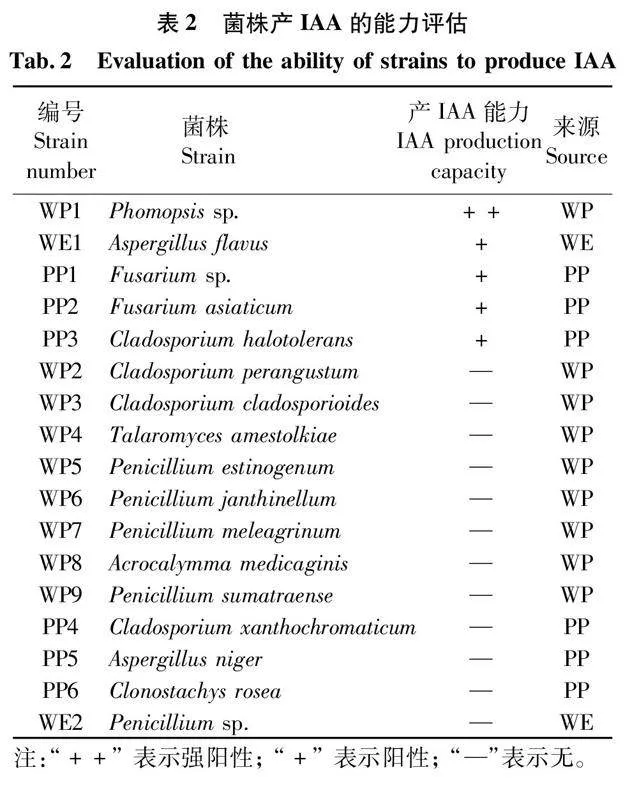
從野生及人工栽培杜仲葉中共分離75種真菌,如表2所示,只有擬莖點霉屬一未定種Phomopsis sp.,黃曲霉Aspergillus flavus,鐮刀菌屬一未定種Fusarium sp.,亞洲鐮刀菌Fusarium asiaticum和深海真菌耐鹽枝孢Cladosporium halotolerans能夠產IAA,其中,有3種屬于人工栽培杜仲葉附生真菌,1種屬于野生杜仲葉附生真菌,1種屬于野生杜仲葉內生真菌。由表可知,擬莖點霉屬一未定種Phomopsis sp.具有顯著陽性顏色反應,產IAA能力最強,高于其他4個菌株。
3討論
本研究從群落組成、多樣性及生態功能等方面對比分析了野生和人工栽培杜仲葉際真菌群落。采用傳統培養法、勻漿平板稀釋分離法對野生和人工栽培條件下的杜仲葉際真菌群落進行培養、分離和純化。
結合形態學和分子生物學方法,對分離獲得的真菌菌株進行鑒定,確定其在各分類單元上的分類地位。結果表明,從野生和人工栽培杜仲葉共分離真菌226株,分屬于42屬75種。在野生杜仲葉附生真菌(WP)優勢屬為青霉屬(32.48%)。符合以往研究結果即:青霉屬真菌是自然界中最大的真菌種屬,也是自然界中最常見的真菌。青霉屬真菌中報道了許多具有抗菌、抗腫瘤、抗病毒等活性的次級代謝產物,是藥物先導化合物的重要來源。其中,青霉素作為抗菌藥物在醫學領域有十分廣泛的應用。此外,環孢菌素作為一種免疫抑制劑,常用于器官移植后的抗排異反應治療;他汀類藥物則可用于降低降膽固醇藥水平[26-27]。
在野生杜仲葉附生真菌(WP)中發現特有屬:擬莖點霉屬。其有性態為子囊菌門的間座殼屬,這類真菌不僅會導致多種植物的葉斑病,還會引起茄子果實的褐紋病,是引起植株產量降低和植株落敗的關鍵因素。林宇等人觀察到一些非洲楝的葉片出現較嚴重的葉斑病,通過顯微鏡觀察,他們將葉斑病的發生歸因于擬莖點霉屬真菌的感染,并將這種葉斑病命名為非洲楝擬莖點霉葉斑病[28]。中國真菌志:擬莖點霉屬中有楝擬莖點霉Phomopsis abdita引起的楝樹枝枯病的記載[29]。SUN等證明了間座殼屬和擬莖點霉屬病原菌復合體對大豆具有致病性[30]。野生杜仲葉內生真菌(WE)優勢屬為間座殼屬,分離頻率為45.71%,屬于植物病原菌。
人工栽培杜仲葉附生真菌(PP)優勢屬為曲霉屬,分離頻率為21.43%。曲霉屬真菌在自然界中廣泛存在,并可應用于多個領域。曲霉屬真菌的某些類群能產生對昆蟲有毒的化合物。據COLE等報道,一株雜色曲霉Aspergillus versicolor可產生雜色酰亞胺,這種化合物對某些昆蟲的成蟲有毒殺和熏蒸作用[31]。因此,這類真菌可在農林害蟲的生物防治策略中成為潛在的工具。另外,在人工栽培杜仲葉附生真菌(PP)中的一特有屬鐮刀菌屬,能分泌多種毒素,可導致作物減產和品質下降,同時也可能對人和動物的健康構成威脅[32]。人工栽培杜仲葉內生真菌(PE)優勢屬為假尾孢菌屬,分離頻率為31.25%。尾孢類真菌是植物病原菌,也是其他植物病原菌知名的重寄生菌,一些種已用于有害雜草的生物防治,是一類具有重要經濟意義的真菌[33]。假尾孢屬是尾孢類真菌中世界性植物病原真菌的大屬,在熱帶和亞熱帶環境中造成谷類、果蔬、林木、藥材等植物的葉斑、疫病、果斑和果腐,致使葉片枯死,過早脫落或造成落花、落果[34]。
結合Shannon-wiener和Margalef這2個微生物群落多樣性指標的分析結果來看,在屬水平上,野生杜仲葉附生(WP)真菌群落多樣性最高,這可能是因為野外生態環境復雜多變,植物微生物與周圍環境中的微生物存在交換,從而導致其真菌群落更為豐富;其次是人工栽培杜仲葉附生(PP)真菌群落。這一結果的出現,可能是由于野生杜仲葉面積遠大于人工栽培杜仲,葉面積越大,允許棲息的微生物種類和數量越多。張曼研究發現,紫蘇等8種藥用植物的葉面積與部分附生真菌的物種多度顯著正相關[35]。目前很多研究是針對植物內生真菌,未來或可對植物附生真菌群落投入更多關注。葉附生真菌較內生真菌多樣性更高,這與AGLER等的研究結果“與內生微生物群相比,葉附生微生物群似乎更加多樣和豐富”一致[36]。此外,野生和人工栽培杜仲葉真菌群落共有屬比例低,具有顯著的空間異質性,這可能是由于真菌對不同環境的選擇偏好形成了不同的群落模式[37]。
在群落功能預測中,在杜仲葉上存在一定種類和數量的腐生型真菌,這些真菌被認為具有降解木質素、纖維素和半纖維素的能力,其可能參與凋落物的分解,釋放大量礦物質養分歸還到土壤,供綠色植物吸收利用[38]。另外,部分真菌還被注釋為內生真菌,這些類群有較強的適應能力,可促進宿主植物生長、土壤養分吸收與利用、改善土壤環境微生物群落結構。然而,也注釋到多種動植物病原菌,如枝孢樣枝孢霉Cladosporium cladosporioides會導致柑橘果實腐敗[39]。因此,揭示杜仲葉際真菌群落組成和功能是了解植株健康以及后續人為調整微生物菌群增強植物健康、提高作物產量和質量的先決條件。
本實驗篩選得到擬莖點霉屬一未定種Phomopsis sp.、黃曲霉Aspergillus flavus等5個具有產IAA活性的菌株,其中Phomopsis sp.的能力最為顯著。SHAO[12]等報道吲哚-3-乙酸和異戊烯基腺嘌呤這兩種激素是影響杜仲葉際微生物群落的關鍵植物激素,在調控其群落組成、多樣性和功能上發揮重要作用。因此,在杜仲的種植過程中,適當加含產吲哚-3-乙酸微生物的菌肥,通過植物-微生物相互作用實現植物生長正向調控,可能更有利于杜仲產量及品質的提升。
綜上所述,在野生和人工栽培條件下的杜仲葉際真菌群落差異顯著,這對優化農業實踐和提升藥材杜仲的質量有重要意義。掌握其中的變化趨勢以及主要影響因素,能夠幫助人們通過調控微生物群落來提高杜仲的藥用價值,也能為其野生資源保護和人工培育提供理論依據。除此之外,利用篩選到的產生長素菌株制成復合型菌肥,在杜仲生長過程中進行人工干預,或可促進杜仲更好更快生長,進而促進杜仲相關產業發展。藥用植物杜仲葉際真菌多樣性豐富,有大量尚待開發利用的微生物資源,有望成為解決杜仲資源與市場需求矛盾的關鍵替代物。參考文獻:
[1]向智, 李煥杰, 諶鑫陽, 等. 藥食同源植物杜仲化學成分、藥理作用及產品開發的研究概況[J]. 中國實驗方劑學雜志, 2024, 30(2): 190-202.
[2] 楊娟, 董醇波, 陳萬浩, 等. 不同地區杜仲樹皮內生真菌群落組成及生態功能結構的差異分析[J]. 中國中藥雜志, 2019, 44(6): 1126-1134.
[3] DONG C B, YAO T, ZHANG Z Y, et al. Structure and function of bacterial microbiota in Eucommia ulmoides bark[J]. Current Microbiology, 2020, 77(11): 3623-3632.
[4] 張青青, 董醇波, 邵秋雨, 等. 杜仲種子內生微生物群落組成及生態功能分析[J]. 林業科學研究, 2023, 36(2): 50-60.
[5] 楊娟. 杜仲相關真菌群落組成及其對活性成分的影響[D]. 貴陽: 貴州大學, 2019.
[6] 孫微微. 杜仲內生真菌的分離鑒定及活性菌株在小麥中的定殖研究[D]. 合肥: 安徽農業大學, 2016.
[7] 王維. 莖點菌屬SP-16F產松脂醇二葡萄糖苷的液體培養條件研究[D]. 楊凌: 西北農林科技大學, 2008.
[8] 徐愛玲, 蔣敏, 白萍萍, 等. 青島城市公園3種植物葉表面顆粒物形態特征及葉際細菌群落結構[J]. 地球環境學報, 2023, 14(4): 458-471.
[9] 魏亞情. 橡膠樹葉際微生物多樣性空間分布格局及其機制[D]. 海口: 海南大學, 2023.
[10]何聃, 任麗娟, 邢鵬, 等. 沉水植物附著細菌群落結構及其多樣性研究進展[J]. 生命科學, 2014, 26(2): 161-168.
[11]柳曉東, 朱海珍, 姜民志, 等. 北京龍形水系三種沉水植物根際及葉際微生物群落特征[J]. 生物工程學報, 2021, 37(10): 3663-3674.
[12]SHAO Q Y, RAN Q S, LI X, et al. Deciphering the effect of phytohormones on the phyllosphere microbiota of eucommia ulmoides[J]. Microbiological Research, 2024, 278: 127513.1-127513.10.
[13]潘明凱. 柑橘葉際腸桿菌功能活性及其耐藥性研究[D]. 昆明: 昆明理工大學, 2022.
[14]梁淑雅, 崔陳陳, 胡思文, 等. 苦草附生細菌的分離鑒定及高效功能菌株篩選[J]. 中國環境科學, 2024, 44(5): 2630-2641.
[15]劉元, 劉淼, 阮元, 等. 懷山藥內生真菌產3-吲哚乙酸能力檢測[J]. 中成藥, 2020, 42(9): 2517-2519.
[16]黃雪珍, 趙龍飛. 藥用植物內生菌對宿主植物促生作用機制研究進展[J]. 微生物學通報, 2023, 50(4): 1653-1665.
[17]張麗娜. 山茶葉部微生物區系與灰斑病發生關系的研究[D]. 雅安: 四川農業大學, 2010.
[18]蘇博.杜仲內生細菌在玉米中的定殖特征及強化抗病機制的研究[D]. 合肥: 安徽農業大學, 2017.
[19]楊娟, 董醇波, 陳萬浩, 梁建東, 等. 不同地區杜仲樹皮內生真菌群落組成及生態功能結構的差異分析[J]. 中國中藥雜志, 2019, 44(6): 1126-1134.
[20]譚玉琴, 張紅芳, 樊有賦, 等. 單葉蔓荊根際土壤真菌多樣性[J]. 菌物研究, 2014, 12(4): 214-217.
[21]沈湛云, 朱波, 張泉龍, 等. 不同產地玄參內生真菌種群結構的比較分析[J]. 中草藥, 2019, 50(4): 957-962.
[22]LANDEWEERT R, LEEFLANG P, KUYPER T W, et al. Molecular identification of ectomycorrhizal mycelium in soil horizons[J]. Applied and Environmental Microbiology, 2003, 69(1): 327-333.
[23]張芝元, 鄭歡, 韓燕峰,等. 不同寵物狗體表一些部位角蛋白降解真菌物種多樣性[J]. 菌物學報, 2017, 36(5): 563-572.
[24]康林玉, 王靜, 劉周斌, 等. 土壤類型對辣椒根系和果實顯微結構影響的研究[J]. 中國農學通報, 2017, 33(28): 73-80.
[25]康林玉. 不同類型土壤下辣椒的生長發育及辣椒種植對微生物的影響[D]. 長沙: 湖南大學, 2018.
[26]孫園園, 于豪冰, 翟祥寧, 等. 青霉屬真菌次級代謝基因挖掘的研究進展[J]. 中國海洋藥物, 2024, 43(1): 74-85.
[27]SEVDAMETOVA E, ZAINOL N. Morphological, physiological,biochemical and molecular characterizationof statin-producing Penicillium microfungi isolated from little-explored tropical ecosystems[J]. Curr Res Microb Sci, 2021, 2: 100044.1-100044.8.
[28]林宇, 李增平, 吳如慧, 等. 非洲楝擬莖點霉葉斑病病原菌鑒定及其生物學特性的測定[J]. 熱帶生物學報, 2019, 10(1): 34-40.
[29]戚佩坤, 姜子德, 向梅梅, 等. 中國真菌志:第34卷 擬莖點霉屬[M]. 北京: 科學出版社,2007.
[30]SUN S, KIM Y M, CHAISAN T, et al. 在韓國引起大豆種腐病的擬莖點霉屬Phomopsis/間座殼屬Diaporthe[J]. 大豆科技, 2014(5): 5.
[31]COLE M, ROLINSON G N. Microbial metabolites with insecticidal properties[J]. Appl Microbiol, 1972, 24(4): 660-662.
[32]張衛娜, 賈諫, 陸曉宇, 等. 鐮刀菌屬真菌毒素的研究進展[J]. 廣東農業科學, 2013, 40(15): 130-133.
[33]郭英蘭. 尾孢類真菌研究進展[J]. 菌物研究, 2020, 18(4): 282-293.
[34]郭英蘭. 中國羽葉楸上的羽葉楸假尾孢新種[J]. 菌物學報, 2017, 36(3): 275-277.
[35]張曼. 安國藥用植物葉際附生微生物多樣性研究[D]. 保定: 河北大學, 2023.
[36]AGLER T M, RUHE J, KROLL S, et al. Microbial hub taxa link host and abiotic factors to plant microbiome variation[J]. PLoS Biology, 2016, 14(1): 1002352.1-1002352.31.
[37]朱攀, 薛文凱, 李小燕, 等. 青藏高原墊狀點地梅葉際及內生可培養微生物多樣性[J]. 微生物學通報, 2023, 50(1): 13-34.
[38]賈梅花, 王哲, 劉金福, 等. 格氏栲天然林凋落物對馬尾松種子萌發的影響[J]. 林業科學研究, 2021, 34(1): 128-136.
[39]解淑慧, 邵興鋒, 王可, 等. 柑橘采后腐爛主要致病菌的分離鑒定及丁香精油對其抑制作用研究[J]. 果樹學報, 2013, 30(1): 134-139, 181.
(責任編輯:于慧梅)
Community Composition of Culturable Fungi in Eucommia Ulmoides
Phyllosphere and Screening of IAA-Producing Strains
QI Yinghua, ZHAO Yufeng, SONG Keyun, WANG Yanling, RAN Qingsong,
SHAO Qiuyu, DONG Chunbo, HAN Yanfeng*
(Institute of Fungus Resources, Guizhou University, Guiyang 550025, China)Abstract: As a unique medicinal plant in China, Eucommia ulmoides is widely distributed in Guizhou province. The phyllosphere microorganisms are of great significance for its growth. In order to reveal the community composition, diversity, and function of phyllosphere fungi in artificially cultivated and wild Eucommia ulmoides, and to screen strains with IAA-production activity, based on the traditional cultivation methods, the composition and diversity of endophytic fungal communities in Eucommia ulmoides phyllosphere was analyzed, and their function groups were annotated based on the FUNGuild database. As a result, a total of 226 fungal strains were isolated from E. ulmoides phyllosphere belonged to 2 phyla and 42 genera. Among them, 154 strains were from wild leaves, mainly including 19 dominant genera such as Penicillium (25.32%), and 72 strains were from artificially cultivated leaves, mainly including 19 dominant genera such as Aspergillus (16.67%). The functional annotation results showed that, except for undefined functional groups, fungal groups mainly involve Animal Pathogen (12.79%), Plant Pathogen Wood Saprotroph (13.95%) and Endophyte Epiphyte Plant Pathogen (24.42%). The fungal species composition of cultivated and wild Eucommia ulmoides was quite different. At the same time, this study also screened several bacterial strains producing IAA. This research will provide valuable microbial resources for the subsequent artificial regulation of E.ulmoides phyllosphere microbiota to enhance plant health, promote plant growth, and develop functional strains.
Key words: plant microbiota; phyllosphere fungi; diversity; ecological functions; indole-3-acetic acid
