夾竹桃天蛾胚胎細(xì)胞的原代培養(yǎng)
2024-01-10 08:30:30李江懷占智高王金昌鄭國華關(guān)麗梅況文東陳俊暉
江西科學(xué) 2023年6期
李江懷,占智高,王金昌,鄭國華,關(guān)麗梅,況文東,陳俊暉,楊 健,靳 亮
(1.江西省科學(xué)院微生物研究所,330096,南昌;2.江西省科學(xué)院生物資源研究所,330096,南昌)
0 引言
從Grace首次建立昆蟲細(xì)胞系以來[1],昆蟲細(xì)胞培養(yǎng)得到了快速的發(fā)展。昆蟲細(xì)胞系成為了生理學(xué)、發(fā)育生物學(xué)和遺傳學(xué)等學(xué)科的重要研究工具[2]。昆蟲細(xì)胞與桿狀病毒表達(dá)系統(tǒng)是目前重要的外源蛋白表達(dá)工具,對(duì)特定病毒敏感的細(xì)胞系是探索病毒侵染機(jī)制的重要研究材料。昆蟲細(xì)胞原代培養(yǎng)是一項(xiàng)經(jīng)典的細(xì)胞生物技術(shù), 從2009年到2018年的十年間,昆蟲細(xì)胞系從500多株增長到了1 000多株,這些細(xì)胞系分別來自于100多種昆蟲[3],這其中有多種成功被用于商業(yè)的昆蟲細(xì)胞系,例如Sf-9、Sf-21和BTI-TN-5B1-4(High Five)。但目前仍然有大量的昆蟲并未建立細(xì)胞系,且缺少可供研究的細(xì)胞株。
夾竹桃天蛾(Oleander Hawkmoth,Daphnisnerii(Linnaeus)),又名粉綠白腰天蛾、鷹紋天蛾、夾竹桃白腰天蛾,屬鱗翅目Lepidoptera,天蛾科Sphingidae,是夾竹桃重要害蟲[4]。該蟲低溫適應(yīng)能力強(qiáng),成蟲有較強(qiáng)的遷越能力,在全球范圍內(nèi)均有分布,其幼蟲能夠?qū)A竹桃等植物造成大面積的侵害,影響夾竹桃生長,甚至造成植株死亡[5]。江西省科學(xué)院微生物研究所2015年分離到包含質(zhì)型多角體病毒DnCPV-NC[6]在內(nèi)的多株新病毒株[7-8],同時(shí)發(fā)現(xiàn)DnCPV-NC具有廣泛的殺蟲譜,具有開發(fā)為新型天蛾科害蟲生物殺蟲劑的潛力[9]。但目前缺乏有效感染的細(xì)胞系,因此,本文意在建立一株對(duì)上述昆蟲病毒敏感的細(xì)胞系,為后續(xù)開展病毒入侵宿主感染機(jī)制提供有效工具。
1 材料與方法
1.1 實(shí)驗(yàn)材料與試劑
1.1.1 實(shí)驗(yàn)用蟲和細(xì)胞系 夾竹桃天蛾蟲卵采自江西省南昌市艾溪湖濕地公園夾竹桃林;Sf-9,Tnh5,C6/36細(xì)胞系由中科院武漢病毒研究所惠贈(zèng),本實(shí)驗(yàn)室保存。
1.1.2 主要試劑 Grace’s insect cell medium(美國Gibco公司),RPMI 1640(美國Gibco公司),TNM-FH insect cell medium(Procell公司),DMEM(Sigma公司),Schneider’s Drosophila Medium(Gibco公司),胎牛血清(美國Gibco公司),三抗(100 μg/mL 硫酸鏈霉素, 100 U/mL 青霉素,2.5 ug/mL兩性霉素B,上海生工)。
1.1.3 設(shè)備 倒置相差顯微鏡(上海儀圓光學(xué)YYE-700),恒溫培養(yǎng)箱(上海新苗GZX-150BS-III),生物安全柜(蘇州蘇凈BHC-1300IIA/B2)。
1.2 實(shí)驗(yàn)方法
1.2.1 原代細(xì)胞分離 將新鮮采集的夾竹桃天蛾蟲卵浸泡于70%乙醇(含1%次氯酸鈉)進(jìn)行表面消毒,然后用雙蒸水清洗3遍,用消毒刀片削破卵殼,將胚胎組織釋放到無血清培養(yǎng)基當(dāng)中。用眼科剪小心地切成小塊,輕輕吹打,用紗布濾去其中大塊的組織。將細(xì)胞懸液低速離心后,除去細(xì)胞碎片,并加入2 mL含15%FBS及1%三抗的培養(yǎng)基,重懸沉淀,轉(zhuǎn)移至細(xì)胞培養(yǎng)板中。
1.2.2 原代細(xì)胞和細(xì)胞系的培養(yǎng) 28 ℃靜置培養(yǎng),根據(jù)培養(yǎng)物的狀態(tài),每5—7 d天對(duì)原代細(xì)胞進(jìn)行換液,每次更換一半培養(yǎng)基。Tnh5和Sf-9細(xì)胞在含10 % FBS的Grace’s培養(yǎng)基中,28 ℃恒溫培養(yǎng),隔3—4 d天傳代一次。C6/36細(xì)胞用添加15 % FBS的RPMI1640培養(yǎng)基,置于5%的CO2培養(yǎng)箱,28 ℃恒溫培養(yǎng)。
1.2.3 細(xì)胞形態(tài)觀察 采用Grace’s培養(yǎng)基?倎28 ℃培養(yǎng)在倒置相差顯微鏡下,觀察細(xì)胞形態(tài)。
2 結(jié)果與分析
2.1 結(jié)果
2.1.1 夾竹桃天蛾原代胚胎細(xì)胞的分離 采集到蟲卵后,將其中色澤鮮艷和卵殼飽滿的挑選出來作為同一批次進(jìn)行原代培養(yǎng)(圖1(a))。對(duì)新獲取的原代培養(yǎng)物進(jìn)行鏡檢,可觀察到不同的細(xì)胞形態(tài),多以圓形油滴狀的胚胎干細(xì)胞 (圖1(b))為主,細(xì)胞大小不一,還能觀察到較小的細(xì)胞團(tuán)和未成形的組織塊(細(xì)胞團(tuán))。剛轉(zhuǎn)移到培養(yǎng)基中的胚胎細(xì)胞以離散狀態(tài)均勻漂浮在培養(yǎng)液中,其中部分干細(xì)胞和細(xì)胞團(tuán)會(huì)在最初的24 h內(nèi)逐漸貼壁。整個(gè)原代培養(yǎng)初期,培養(yǎng)液中都懸浮著一定量的離散或成團(tuán)的細(xì)胞,這些細(xì)胞幾乎不增殖,在一個(gè)月內(nèi)隨著培養(yǎng)液的更換而被清除出培養(yǎng)板。

圖1 (a)從夾竹桃樹葉上采集到的夾竹桃天蛾蟲卵 (b)夾竹桃天蛾胚胎細(xì)胞形態(tài)(100×)(標(biāo)尺 = 50 μm)
2.1.2 原代培養(yǎng)一周后細(xì)胞形態(tài)觀察結(jié)果 選擇最適合的培養(yǎng)基是原代培養(yǎng)的關(guān)鍵之一,昆蟲細(xì)胞系的成功建立高度依賴于培養(yǎng)基和起源組織,尤其是適合細(xì)胞類型的培養(yǎng)基[9]。在Grace’s培養(yǎng)基中原代細(xì)胞呈長梭型和圓形(圖2(a)),和TNM-FH培養(yǎng)基中均有不少胚胎細(xì)胞以細(xì)胞團(tuán)的形式貼壁(圖2(b)),而Schneider培養(yǎng)基中的細(xì)胞貼壁少只能觀察到少數(shù)細(xì)胞團(tuán)(圖2(c)),DMEM和RPMI1640培養(yǎng)基中則難以貼壁(圖2(d),(e))。

圖2 培養(yǎng)7 d的夾竹桃天蛾胚胎原代細(xì)胞(100x) aGrace’s bTNM-FH cSchneider dDEME eRPMI 1640(標(biāo)尺 = 50 μm)
2.1.3 夾竹桃天蛾原代培中胚胎細(xì)胞的分化和形態(tài)觀察結(jié)果 整個(gè)原代培養(yǎng)過程,胚胎細(xì)胞在形態(tài)分化上展現(xiàn)了高度的多樣性,多種不同形態(tài)的細(xì)胞依次出現(xiàn)在培養(yǎng)物當(dāng)中,最早出現(xiàn)的是長梭形細(xì)胞和圓形細(xì)胞(圖3(a)),通常在細(xì)胞貼壁一周后陸續(xù)出現(xiàn)在培養(yǎng)物中。紡錘形細(xì)胞(圖3(b))只存在較短的時(shí)間,成纖維狀細(xì)胞(圖3(c))通常在原代培養(yǎng)的第一個(gè)月有較快的生長速度,然后就陷入長期的靜止?fàn)顟B(tài)。少量的豆莢形細(xì)胞曾短暫存在過(圖3(d)),這些細(xì)胞的個(gè)體相對(duì)其他類型要大出數(shù)倍。原代培養(yǎng)初期,可觀察到部分肌肉細(xì)胞在有節(jié)律地跳動(dòng)(圖3(e)),頻率約為每分鐘20~30次,但這些跳動(dòng)的細(xì)胞通常在2—3個(gè)月的培養(yǎng)后逐漸消失在培養(yǎng)物中。此外,還有少量的扁平的圓形細(xì)胞(圖3(f))和相對(duì)較小的顆粒型細(xì)胞(圖3(g))也被觀察到。而成團(tuán)的細(xì)胞周圍通常會(huì)有些離散出來的多邊形細(xì)胞(圖3(h)),這類細(xì)胞大小中等,可明顯看到細(xì)胞核,細(xì)胞的偽足呈不規(guī)則多邊形。夾竹桃天蛾原代培養(yǎng)細(xì)胞與本實(shí)驗(yàn)室培養(yǎng)的多種昆蟲細(xì)胞系,包括Tnh5、sf-9和C6/36等細(xì)胞系(圖3(i),(j),(k)),在形態(tài)與生長速度上存在顯著差異。
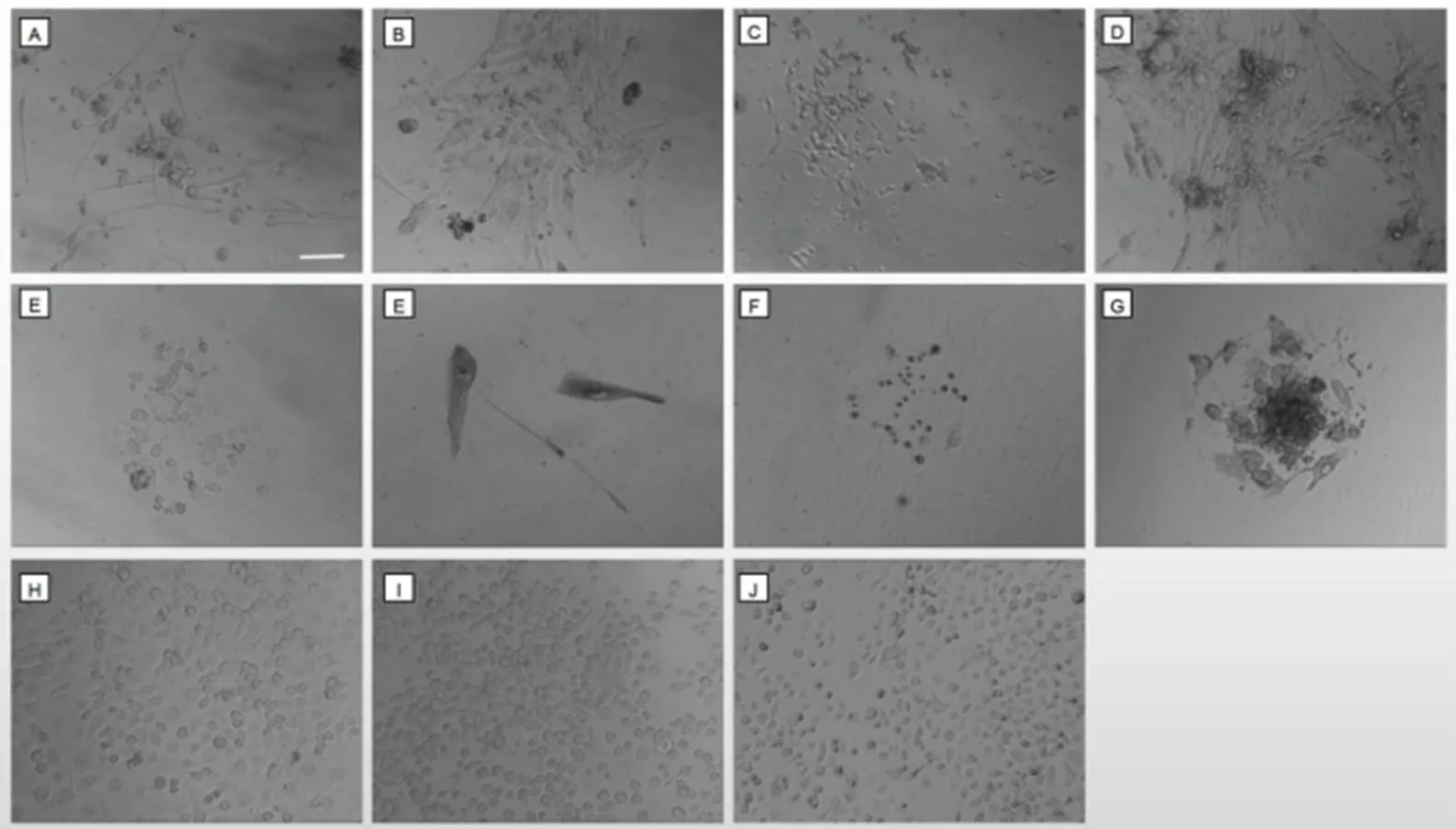
(a)類脂肪細(xì)胞;(b)肌肉細(xì)胞;(c)長梭形細(xì)胞;(d)成纖維狀細(xì)胞;(e)圓形細(xì)胞;(f)豆莢形細(xì)胞;(g)顆粒狀細(xì)胞;(h)多邊形細(xì)胞;(i)Tnh5細(xì)胞系;(j)Sf-9細(xì)胞系;(k)C6/36細(xì)胞系(標(biāo)尺 = 50 μm)圖3 夾竹桃天蛾原代細(xì)胞培養(yǎng)初期的細(xì)胞形態(tài)以及與其它昆蟲細(xì)胞系對(duì)比(100×)
2.1.4 長期培養(yǎng)后的細(xì)胞狀態(tài)觀察結(jié)果 夾竹桃天蛾原代細(xì)胞在經(jīng)過長達(dá)一年的原代培養(yǎng)過程,處于相對(duì)靜止的狀態(tài),與外界交換緩慢,培養(yǎng)每7 d更換一次培養(yǎng)液。經(jīng)過長期的換液后,原代細(xì)胞主要呈現(xiàn)肌肉型(圖4(a))與多邊形(圖4(b)細(xì)胞為主的混合形態(tài)。

圖4 夾竹桃天蛾原代細(xì)胞培養(yǎng)12Mo(100×)(標(biāo)尺 = 50 μm)
3 討論與結(jié)論
污染和培養(yǎng)基結(jié)晶是原代培養(yǎng)失敗的重要原因[10]。以往的研究者對(duì)昆蟲原代細(xì)胞培養(yǎng)的各個(gè)技術(shù)環(huán)節(jié)都做了大量的研究和實(shí)驗(yàn),培養(yǎng)基的選擇、血清濃度、滲透壓、pH等培養(yǎng)條件對(duì)細(xì)胞貼壁、遷移、增殖的影響[11]。鱗翅目昆蟲胚胎細(xì)胞系的建立已經(jīng)比較成熟,基于Grace’s 昆蟲培養(yǎng)基和TNM-FH培養(yǎng)基,棉鈴蟲[12]、小菜蛾[13]、玉米螟[14]等多種昆蟲的細(xì)胞系得以建立。本研究利用這2種培養(yǎng)基,成功對(duì)夾竹桃天蛾卵來源的胚胎細(xì)胞進(jìn)行了持續(xù)培養(yǎng),其中Grace’s培養(yǎng)基中的培養(yǎng)物多為離散分布的細(xì)胞,平鋪生長,呈現(xiàn)出豐富多樣的細(xì)胞形態(tài)(圖3(a)~(f)),其中,纖維狀細(xì)胞、梭形細(xì)胞、肌肉細(xì)胞分別約占原代細(xì)胞總數(shù)的30%、30%和20%,這些細(xì)胞在增殖過程中具備一定的遷移能力[15]。TNM-FH中則以聚集的細(xì)胞團(tuán)為主,新生細(xì)胞以細(xì)胞團(tuán)為中心呈放射狀生長,約占到細(xì)胞總量的60%,對(duì)于細(xì)胞形態(tài)多樣性的維持弱于Grace’s培養(yǎng)基。由于不確定哪種形態(tài)的細(xì)胞對(duì)昆蟲病毒是易感細(xì)胞,為保持原代細(xì)胞中盡可能多的細(xì)胞種類,因而選擇Grace’s培養(yǎng)基作為夾竹桃天蛾胚胎細(xì)胞的最佳培養(yǎng)基。實(shí)驗(yàn)中分離到夾竹桃天蛾胚胎細(xì)胞中由大量離散的干細(xì)胞組成,因此未進(jìn)行胰酶消化和機(jī)械分散。在對(duì)蟲卵解剖后,釋放到培養(yǎng)液中的即是胚胎干細(xì)胞和發(fā)育中的組織塊。
胚胎、卵巢等的組織是昆蟲細(xì)胞原代培養(yǎng)最常用到的材料,具有易于獲取和分化潛力強(qiáng)等優(yōu)點(diǎn)。目前已經(jīng)建立的1 200株昆蟲細(xì)胞系而言,其中一半來源于胚胎組織,這可能源于胚胎干細(xì)胞巨大分化潛能[16]。而中腸、神經(jīng)系統(tǒng)等組織則難度較大,一方面是因?yàn)樵囵B(yǎng)過程中的污染難以控制,另一方面可能是分化成熟的組織難以適應(yīng)體外的培養(yǎng)環(huán)境。國內(nèi)目前僅有一株來源于棉鈴蟲中腸的細(xì)胞系建立成功[17]。雖然昆蟲生物學(xué)的研究人員在細(xì)胞原代培養(yǎng)上取得了很多成果,但并不是每一株細(xì)胞系都能成為合適的實(shí)驗(yàn)材料。細(xì)胞在原代培養(yǎng)過程中的遺傳物質(zhì)可能會(huì)發(fā)生不確定性的改變,使得原代細(xì)胞特性產(chǎn)生與昆蟲體內(nèi)的細(xì)胞差異,這些差異對(duì)于后續(xù)的研究也存在著諸多不可預(yù)測(cè)的影響。本研究中選擇夾竹桃天蛾的蟲卵作為原代培養(yǎng)的材料,因其胚胎由堅(jiān)硬厚實(shí)的卵殼保護(hù),最大程度降低了原代培養(yǎng)過程中消毒操作的難度和污染的幾率,且成功地在Grace’s培養(yǎng)基中維持了原代細(xì)胞存活超過12個(gè)月。由于目前夾竹桃天蛾的人工養(yǎng)殖難度較高,原代培養(yǎng)獲取的材料主要來自于野生環(huán)境,選擇蟲卵作為原代培養(yǎng)的材料也可盡可能降低被昆蟲病毒感染的幾率,從而在源頭上降低了原代細(xì)胞被昆蟲病毒感染的風(fēng)險(xiǎn),在漫長培養(yǎng)過程中持續(xù)的觀察也證實(shí)了這一點(diǎn)。
哺乳動(dòng)物細(xì)胞可通過對(duì)腫瘤細(xì)胞進(jìn)行原代培養(yǎng)的方式以快速得到相應(yīng)的細(xì)胞系[18],或采用慢病毒定向整合基因的方式對(duì)原代細(xì)胞實(shí)現(xiàn)永生化[19]。昆蟲細(xì)胞取材等原因及基礎(chǔ)研究不足,難以快速建立需要的細(xì)胞系。Li等[20]通過MNNG處理原代細(xì)胞實(shí)現(xiàn)了化學(xué)誘導(dǎo)昆蟲細(xì)胞轉(zhuǎn)化并建立了細(xì)胞系,說明化學(xué)誘導(dǎo)的方法對(duì)昆蟲細(xì)胞的轉(zhuǎn)化是可行的。張欣等[21]通過重組桿狀病毒在培養(yǎng)體系中引入端粒酶逆轉(zhuǎn)錄酶獲得了可連續(xù)傳代的美洲大蠊的細(xì)胞系,并顯著縮短了原代培養(yǎng)所需的時(shí)間。張寰等[22]通過物理或化學(xué)低氧的方式來處理原代細(xì)胞可誘導(dǎo)其加快轉(zhuǎn)化的進(jìn)程。這些新開發(fā)的手段均為極具潛力的快捷建系方法,并有待于廣大研究者的應(yīng)用和檢驗(yàn)。唐凱等[23]通過轉(zhuǎn)錄組測(cè)序發(fā)現(xiàn)原代細(xì)胞在離體培養(yǎng)的過程中,cdk4,cycd,skp2和mad1等多個(gè)基因與細(xì)胞的轉(zhuǎn)化相關(guān),在分子生物學(xué)層面為昆蟲細(xì)胞建系提供了創(chuàng)新的理論基礎(chǔ)。本研究探索了對(duì)夾竹桃天蛾的胚胎細(xì)胞原代培養(yǎng)的條件,通過機(jī)械破碎卵殼分離的原代細(xì)胞在多種培養(yǎng)基中順利貼壁,在Grace’s培養(yǎng)基中維持了原代細(xì)胞12個(gè)月,觀察到原代細(xì)胞分化出的多種形態(tài),細(xì)胞緩慢增殖。原代細(xì)胞的鑒定包括DAF-PCR、染色體核型分析及對(duì)桿狀病毒和DnCPV的敏感性實(shí)驗(yàn)需在原代細(xì)胞開始傳代后展開。以胚胎組織為材料建立的細(xì)胞系,細(xì)胞形態(tài)具備較高的多樣性,若能通過單細(xì)胞克隆分離出混合在夾竹桃天蛾原代細(xì)胞中肌肉細(xì)胞、脂肪細(xì)胞等細(xì)胞株,將為昆蟲細(xì)胞離子通道和脂類代謝調(diào)節(jié)機(jī)制的研究奠定基礎(chǔ)。
建立新的昆蟲細(xì)胞系和發(fā)現(xiàn)新的昆蟲病毒一直在昆蟲病理研究當(dāng)中發(fā)揮著相輔相成的作用,二者缺一不可。對(duì)于昆蟲病毒方面的研究人員來說,發(fā)現(xiàn)一株新的昆蟲病毒,就至少需要有一株相應(yīng)的敏感細(xì)胞系來實(shí)現(xiàn)細(xì)胞水平的機(jī)理研究,這也是下一步開展的工作內(nèi)容。

- 江西科學(xué)的其它文章
- 人工智能社會(huì)的戰(zhàn)略問題與應(yīng)對(duì)措施
- 假儉草不同種植方式在鄱陽湖區(qū)堤防的適宜性實(shí)證研究
- 《插花藝術(shù)》課程教學(xué)改革探索與實(shí)踐
——以江西財(cái)經(jīng)大學(xué)為例 - 基于AHP-熵權(quán)法的省級(jí)科技專項(xiàng)資金投入規(guī)模和資金分配綜合評(píng)價(jià)研究
——對(duì)江西省科學(xué)院2020—2022年數(shù)據(jù)的考察 - 基于資源特色的安義古村保護(hù)策略研究
- 基于規(guī)則引擎的氣象臺(tái)站元數(shù)據(jù)質(zhì)量控制方法