中國熱帶珊瑚島優(yōu)勢(shì)木本植物抗風(fēng)桐和草海桐的水分適應(yīng)策略解析
2024-02-02 06:07:14唐瑋澤李沁張浩萍金益李強(qiáng)殷德意葉清劉慧
熱帶亞熱帶植物學(xué)報(bào) 2024年1期
關(guān)鍵詞:植物
唐瑋澤, 李沁, 張浩萍, 金益, 李強(qiáng), 殷德意, 葉清,劉慧*
中國熱帶珊瑚島優(yōu)勢(shì)木本植物抗風(fēng)桐和草海桐的水分適應(yīng)策略解析
唐瑋澤1,2,3, 李沁1,2,3, 張浩萍1,2,3, 金益1,3, 李強(qiáng)4, 殷德意1,3, 葉清1,3,劉慧1,3*
(1. 廣東省應(yīng)用植物學(xué)重點(diǎn)實(shí)驗(yàn)室,中國科學(xué)院退化生態(tài)系統(tǒng)植被恢復(fù)與管理重點(diǎn)實(shí)驗(yàn)室,中國科學(xué)院華南植物園,廣州 510650;2. 中國科學(xué)院大學(xué),北京 100049;3. 華南國家植物園,廣州 510650;4. 海南醫(yī)學(xué)院,熱帶醫(yī)學(xué)院,海口 571199)
植物水力性狀能夠反映植物對(duì)不同水分條件的適應(yīng)能力,研究熱帶珊瑚島特殊生境下優(yōu)勢(shì)植物的水力功能特征對(duì)深入理解熱帶珊瑚島植物的水分適應(yīng)策略,從而選擇熱帶珊瑚島植被構(gòu)建和恢復(fù)的適生物種具有重要意義。該研究以中國熱帶珊瑚島生境中2種優(yōu)勢(shì)適生木本植物:抗風(fēng)桐()和草海桐()為研究對(duì)象,比較了其葉片和枝條的水力性狀,并分析了其水分適應(yīng)策略。結(jié)果表明, 抗風(fēng)桐的葉片栓塞抗性、枝條邊材比導(dǎo)水率和葉片膨壓?jiǎn)适c(diǎn)顯著高于草海桐,而枝條栓塞抗性、葉片導(dǎo)水率、邊材密度和葉面積邊材面積比均顯著低于草海桐。抗風(fēng)桐的葉片具有比枝條更強(qiáng)的抗栓塞能力,對(duì)水分脅迫敏感,但同時(shí)選擇以高效的枝干水分運(yùn)輸來滿足葉片高蒸騰需求的充足供水。草海桐枝條與葉片則存在水力脆弱性分區(qū),在面臨水分脅迫時(shí)葉片充當(dāng)“安全閥”以保證枝干木質(zhì)部的水力安全。抗風(fēng)桐與草海桐能夠通過協(xié)調(diào)葉片與枝條水力性狀采取不同的水分適應(yīng)策略,從而更好地適應(yīng)熱帶珊瑚島的特殊生境。
植物水力性狀;栓塞;水力脆弱性分區(qū);水分運(yùn)輸;熱帶珊瑚島植物
熱帶珊瑚島是分布于熱帶海域的特殊陸地類型,主要由死亡后的珊瑚蟲殘?bào)w堆筑而成。因此, 珊瑚島的土壤主要由珊瑚殘屑經(jīng)長期生物作用下形成的磷質(zhì)石灰土和珊瑚砂沖積至海邊形成的濱海沙積土組成,粗沙粒多,缺乏粘粒,具有高鹽、強(qiáng)堿性及含水量低等特點(diǎn)[1–2]。中國的珊瑚島廣泛分布于南海地區(qū),在高溫和強(qiáng)光輻射等極端條件下,植物生長困難,難以形成穩(wěn)定、多樣化的熱帶珊瑚島植被,且極易發(fā)生退化。然而,植被是熱帶珊瑚島生態(tài)系統(tǒng)形成的基礎(chǔ),可供給、調(diào)節(jié)和支持各項(xiàng)生態(tài)系統(tǒng)服務(wù)功能[3]。構(gòu)建和恢復(fù)熱帶珊瑚島植被能夠?yàn)闊釒汉鲘u提供宜居環(huán)境,對(duì)促進(jìn)熱帶珊瑚島生態(tài)系統(tǒng)的可持續(xù)發(fā)展具有重要價(jià)值。研究珊瑚島適生植物的功能特征及其對(duì)環(huán)境的適應(yīng)機(jī)理,是熱帶珊瑚島植被構(gòu)建和生態(tài)恢復(fù)的前提和理論基礎(chǔ)。
植物水力性狀是指與植物水分吸收、轉(zhuǎn)運(yùn)和喪失過程有關(guān)的功能特征,反映植物對(duì)環(huán)境水分條件的適應(yīng)能力[4]。植物可在一定程度上調(diào)整水力性狀,減小環(huán)境水分波動(dòng)帶來的不利影響,維持自身的正常生長發(fā)育,從而逐步形成有效適應(yīng)當(dāng)?shù)丨h(huán)境水分條件的水分適應(yīng)策略[5],因此植物水力性狀可以作為評(píng)價(jià)植物對(duì)熱帶珊瑚島水分條件適應(yīng)性的重要依據(jù)。植物水力性狀可以分為2個(gè)方面:與木質(zhì)部抗栓塞能力有關(guān),代表水力安全的性狀;與木質(zhì)部水分運(yùn)輸效率有關(guān),代表水力效率的性狀[4]。水分在植物蒸騰拉力和水分子內(nèi)聚力的作用下,于木質(zhì)部中形成連續(xù)體保持水分的向上運(yùn)輸。在面臨植物干旱脅迫時(shí),木質(zhì)部張力會(huì)導(dǎo)致空氣由導(dǎo)管上的紋孔進(jìn)入木質(zhì)部管道,導(dǎo)致栓塞發(fā)生。栓塞將在木質(zhì)部管腔擴(kuò)散,阻礙水分運(yùn)輸,持續(xù)發(fā)展的栓塞阻礙植物的正常生理活動(dòng),最終導(dǎo)致植物死亡[6]。木質(zhì)部抵抗栓塞形成和擴(kuò)張的能力通常使用木質(zhì)部導(dǎo)水率損失50%時(shí)的水勢(shì)(P50)衡量,反映木質(zhì)部水分運(yùn)輸?shù)陌踩訹7]。葉片膨壓?jiǎn)适c(diǎn)(tlp)反映植物葉片在干旱脅迫下維持細(xì)胞膨壓的能力[8]。tlp越低的植物抵抗葉片脫水的能力越強(qiáng),從而可在水分有效性較低的條件下維持氣孔開放,保證光合作用的正常進(jìn)行[9]。在水力效率方面,枝條邊材比導(dǎo)水率(Ks)可以代表枝條尺度的木質(zhì)部水分運(yùn)輸效率,葉片導(dǎo)水率(Kleaf)反映葉片尺度的水分傳輸效率[4,10]。一般認(rèn)為,水分運(yùn)輸效率越高的植物,其光合速率越高,徑向生長也越快,可以歸為“資源獲取型策略”,但采取這種策略可能會(huì)降低植物的水力安全性[11]。
植物的水力脆弱性分區(qū)假說認(rèn)為,葉片在面對(duì)木質(zhì)部空化引發(fā)的栓塞時(shí),比枝條和主干表現(xiàn)得更為脆弱(即葉片P50值更高)[12],因此植物可利用葉片作為安全閥,將栓塞限制在碳投資較低的器官即葉片,從而避免枝條和莖干等碳投資較高的器官出現(xiàn)嚴(yán)重的水力功能障礙[13–14],這可能是植物為應(yīng)對(duì)干旱脅迫,維護(hù)木質(zhì)部水分運(yùn)輸安全所采取的重要策略。同時(shí),前人還報(bào)道了亞熱帶濕潤地區(qū)的一些物種缺乏水力脆弱性分區(qū),即枝條比附于其上的葉片更容易發(fā)生栓塞。這些物種的葉片在干旱脅迫下可能失去作為“安全閥”的功能,但可能用其他補(bǔ)償性的水力策略來維持水分平衡[14]。然而,熱帶地區(qū)代表性木本植物采取何種水力脆弱性分區(qū)的策略,尚缺乏研究。
植物的形態(tài)結(jié)構(gòu)性狀也與光合、水力等功能聯(lián)系緊密,可以很好的表征植物的生態(tài)適應(yīng)策略。如邊材密度(WD)具有廣泛的生理意義,植物的高邊材密度與更高的耐旱性和生存率有關(guān),因?yàn)檫@類植物往往具有更強(qiáng)的抗栓塞能力和機(jī)械抗性[15]。葉面積邊材面積比(Al/As)代表單位邊材橫截面積可供給的莖末端葉片面積,反映了莖干邊材木質(zhì)部水分運(yùn)輸能力與葉片光合潛能之間的權(quán)衡,與個(gè)體生物量密切相關(guān),是植被水分利用模型中的重要參數(shù)[16]。在干旱脅迫下,Al/As降低可以減少整株蒸騰耗水,從而提高剩余葉片的水分供給,維持植物的水分平衡[16]。
抗風(fēng)桐()和草海桐()分別為紫茉莉科(Nyctaginaceae)常綠喬木和草海桐科(Goodeniaceae)常綠亞灌木,是中國熱帶珊瑚島植被中最常見的優(yōu)勢(shì)喬木樹種和灌木樹種[17]。二者均具有較高的生長速率和無性繁殖能力(離體組織再生能力)[18–19],且耐鹽堿和干旱,在海岸固沙、調(diào)節(jié)熱帶珊瑚島氣候及熱帶珊瑚島植被恢復(fù)中發(fā)揮了重要作用[20–21]。同時(shí)2種植物因花型獨(dú)特, 造型優(yōu)美,可作園林觀賞樹種,其葉片可作藥用和飼料,也具有重要的生態(tài)和經(jīng)濟(jì)價(jià)值[22–23]。目前,對(duì)這2種植物的研究主要集中在形態(tài)生物學(xué)特性及藥用、經(jīng)濟(jì)學(xué)特性等方面[21],而對(duì)其水力功能特性的研究較少[22–23]。因此,本研究以海南省文昌市熱帶珊瑚島植被恢復(fù)實(shí)驗(yàn)基地培育的抗風(fēng)桐和草海桐為研究對(duì)象,通過測(cè)量其枝條和葉片的水力性狀, 結(jié)合形態(tài)解剖結(jié)構(gòu),分析其水力功能特性及水分適應(yīng)策略,為熱帶珊瑚島植被構(gòu)建與恢復(fù)的物種選擇提供參考。
1 材料和方法
1.1 研究地概況
本研究地點(diǎn)位于海南省文昌市熱帶珊瑚島植被恢復(fù)實(shí)驗(yàn)基地(110°45′ E,19°31′ N),為典型的熱帶海洋性季風(fēng)氣候,氣溫變化小,年均氣溫24 ℃,年均降水量1 800 mm,水熱條件豐富,但降水時(shí)間分布不均,每年6月至11月為濕季,12月至翌年5月為干季,具有明顯的季節(jié)性干旱現(xiàn)象。土壤基質(zhì)為濱海沉積物沙積土,與熱帶珊瑚島上的常綠喬木和常綠灌木的土壤基質(zhì)基本一致,濕季土壤含水量約為10.5%,干季土壤含水量約為8.2%,但因?yàn)闊釒汉鲘u降雨較多,濕季水分有效性依然較高[17]。實(shí)驗(yàn)基地的氣候條件也和熱帶珊瑚島基本一致,但植物生長和養(yǎng)護(hù)條件良好,環(huán)境均一,因此避免了熱帶珊瑚島生境異質(zhì)性導(dǎo)致植物個(gè)體間和物種間差異性來源難以確定的問題,保證了2種植物在同一環(huán)境條件下測(cè)量結(jié)果的可比性,能更準(zhǔn)確的反映植物自身的水力性狀差異。
1.2 試驗(yàn)材料和方法
基于前期對(duì)中國熱帶珊瑚島植物的調(diào)查及研究,本研究選取熱帶珊瑚島特殊生境(高溫、強(qiáng)光和強(qiáng)堿等)的2種優(yōu)勢(shì)適生木本植物:抗風(fēng)桐()和草海桐()作為研究對(duì)象。其中抗風(fēng)桐為多年生常綠喬木,樹皮灰白且皮孔明顯,卵形對(duì)生紙質(zhì)葉片;草海桐為多年生常綠亞灌木,匙形至倒卵形肉質(zhì)葉片,螺旋狀排列于分枝頂端[17]。二者均具有生長速度快、斷枝可再殖,耐強(qiáng)光、干旱和貧瘠等特性[16,21]。2022年7月,每種植物選取株高接近(抗風(fēng)桐為2.5 m,草海桐為1.5 m)且長勢(shì)健康的植株,采集冠層周圍無遮蔭、長度為1.0~1.4 m的帶葉枝條,并立即在水下修剪切口并套上黑色塑料袋(防止水分散失和外界空氣等進(jìn)入被切開的導(dǎo)管),立即帶回實(shí)驗(yàn)室。本實(shí)驗(yàn)中所有用水均為超純水,并用真空泵抽氣6 h以上以排除水中的氣體。
1.3 枝條邊材比導(dǎo)水率
枝條邊材比導(dǎo)水率采用低壓液流法測(cè)定[24]。將采回的樣品在實(shí)驗(yàn)室內(nèi)暗適應(yīng)30~60 min,使用注氣法測(cè)量最大導(dǎo)管長度[25]。將收集樣段的形態(tài)學(xué)頂端枝條用鋒利的刀片修剪平滑,并連接一條塑料軟管,在0.1 MPa的壓力下注入空氣。塑料軟管的末端浸入水中,自樣段末端起以2 cm長度在水中連續(xù)切割,直到樣段末端出現(xiàn)第1個(gè)氣泡,此時(shí)剩余的樣段長度為最大導(dǎo)管長度(Lmax, cm)。在水下將待測(cè)樣段切割成約30~ 50 cm (超過其最大導(dǎo)管長度, 抗風(fēng)桐約為24.5 cm,草海桐約為21.5 cm)并記錄(L, m)。將樣段兩端剝?nèi)?~ 2 cm韌皮部后用生料帶纏繞,連入蠕動(dòng)泵(KCP Pro, Kamoer, Shanghai, China)橡皮管并檢查氣密性。首先在0.15 MPa的壓力下沖洗莖段20 min,去除導(dǎo)管中的氣泡。測(cè)定使用的溶液為除氣后的10 mmol/L KCl溶液。利用50 cm高度的水柱產(chǎn)生的靜水壓力(P, 約0.005 MPa)驅(qū)動(dòng)液流通過樣段,樣段下端連接到移液管,并記錄移液管中凹液面通過一定數(shù)量連續(xù)刻度標(biāo)記所需要的時(shí)間,計(jì)算流速Jv(kg/s), 同時(shí)通過溫濕計(jì)測(cè)定樣段附近空氣的溫度(T,℃)和濕度(RH, %), 保持空氣溫度為22 ℃~28 ℃。計(jì)算樣段最大導(dǎo)水率[Kmax, (kg·m)/(s·MPa)]=Jv/(?P/?L)。
測(cè)定完成后,使用體視顯微鏡拍攝莖段基部的橫截面,使用ImageJ (National Institutes of Health, Bethesda, USA)測(cè)定木質(zhì)部面積(As, m2),計(jì)算枝條木質(zhì)部邊材比導(dǎo)水率[Ks, kg/(m·s·MPa)]=Kmax/As。
1.4 枝條脆弱性曲線
枝條脆弱性曲線采用自然干燥法測(cè)定[24]。用錫箔紙和自封袋將采回的枝條欲測(cè)莖段的上游和下游4~6片成熟葉片包裹后,將切口端插入水桶中, 同時(shí)在水下剪去約10 cm的一段后,用黑色塑料袋包裹復(fù)水2 h,以平衡枝條水勢(shì)。復(fù)水結(jié)束后,用壓力室(plant moisture stress, Corvallis, USA)分別測(cè)定被包裹的上游和下游端葉片水勢(shì),當(dāng)兩者的水勢(shì)之差小于0.2 MPa時(shí),認(rèn)為此時(shí)的葉片水勢(shì)等同于枝條的木質(zhì)部水勢(shì)(x, MPa)。將平衡后的枝條剪去葉片,并在切口端和末端在水槽中各切去一截,讓枝條在水中釋放木質(zhì)部張力30 min。將釋放張力后的枝條兩端切口修平后連入上述導(dǎo)水率測(cè)量裝置測(cè)量此時(shí)的實(shí)際導(dǎo)水率Ki,隨后在0.15 MPa的壓力下沖洗莖段20 min后測(cè)量其最大導(dǎo)水率Kmax。其余枝條在室內(nèi)(26 ℃)自然晾干,達(dá)到不同的水勢(shì)梯度,然后重復(fù)上述過程,測(cè)得不同水勢(shì)下的枝條Ki與Kmax并計(jì)算導(dǎo)水損失率(PLC, %):PLC=100(1–Ki/Kmax)。
將水勢(shì)與PLC的關(guān)系構(gòu)建曲線即為枝條脆弱性曲線,擬合方程為PLC=100/{1+exp[a(-b)]},其中a和b為常數(shù)。根據(jù)擬合曲線得到枝條導(dǎo)水率損失50%時(shí)的水勢(shì)P50 branch作為枝條栓塞脆弱性的指示指標(biāo)[24]。
1.5 葉片壓力-容積曲線
葉片壓力-容積曲線參照Tyree等[26]的方法。從復(fù)水后的枝條上剪下成熟健康葉片進(jìn)行測(cè)定,記錄葉片水勢(shì)(sat, MPa),確保其初始值均高于–0.05 MPa, 稱量此時(shí)的葉片質(zhì)量即為飽和鮮重(Wsat, mg)。此后將葉片置于天平上進(jìn)行自然干燥,在失去2~5 mg水分后重新記錄葉片質(zhì)量(Wi, mg)和測(cè)定此時(shí)的葉片水勢(shì)(i, MPa),重復(fù)以上過程,直至葉片萎蔫。然后于70 ℃烘箱干燥葉片48 h,測(cè)量其干質(zhì)量(Wd, mg)。相對(duì)含水量(RWC, %)=100(Wi-Wd)/(Wsat-Wd), 基于葉面積的葉片水容[Cleaf, mmol/(m2·MPa)]=RWC/i(Wd/Aleaf)(Wsat/Wd)/M, 式中,Aleaf為葉面積(m2),M為水的摩爾質(zhì)量(g/mol)。
以1/i為縱坐標(biāo),(1-RWC)為橫坐標(biāo)構(gòu)建的曲線即為葉片壓力-容積曲線,利用Schulte等[27]編制的程序確定葉片膨壓?jiǎn)适c(diǎn)時(shí)的水勢(shì)(tlp, MPa)。
1.6 葉片脆弱性曲線
葉片脆弱性曲線采用復(fù)水法測(cè)定[28]。將采回的帶葉枝條在自然狀態(tài)下失水不同時(shí)間,以形成不同的葉片水勢(shì)梯度,達(dá)到不同水勢(shì)梯度的枝條需進(jìn)行暗處理至少30 min使枝條整體水勢(shì)達(dá)到平衡。每個(gè)枝條選取至少1片葉片用壓力室測(cè)定初始水勢(shì)(0, MPa),然后在水下切下相鄰葉片,在暗處復(fù)水一定時(shí)間(, 30~300 s),復(fù)水完成后取出葉片,迅速擦干,然后測(cè)定復(fù)水后水勢(shì)(f, MPa)。結(jié)合葉片壓力-容積曲線得到的葉片水容Cleaf計(jì)算葉片導(dǎo)水率[Kleaf,mmol/(m2·s MPa)]=Cleafln(0/f)/t。
將所有測(cè)得的初始葉片水勢(shì)大于-0.3 MPa的葉片導(dǎo)水率取平均值作為葉片最大導(dǎo)水率(Kmax leaf), 根據(jù)不同葉片水勢(shì)下測(cè)定的葉片導(dǎo)水率與Kmax leaf計(jì)算導(dǎo)水損失率(PLC, %):PLC=100(1-Kleaf/Kmax leaf)。
將葉片水勢(shì)與PLC的關(guān)系構(gòu)建曲線即為葉片脆弱性曲線,擬合方程為PLC=100/{1+exp[a(-b)]},以葉片導(dǎo)水率損失50%時(shí)的水勢(shì)P50 leaf作為葉片栓塞脆弱性的指示指標(biāo)[27]。
1.7 邊材密度
從測(cè)定導(dǎo)水率后的枝條(直徑5~10 mm)截取一段長度約5 cm的無分支莖段,去除髓心和樹皮, 浸泡在水中過夜,充分飽和后利用排水法測(cè)定邊材體積(V, cm3),然后置于70 ℃烘箱中烘72 h后測(cè)定干重(DW, g),邊材密度(WD, g/cm3)即為DW與的比值。
1.8 葉面積邊材面積比
將取回的枝條形態(tài)學(xué)末端上所有的葉片取下,使用葉面積儀(Li-3000A, Li-Cor, Lincoln, USA)測(cè)定其總?cè)~面積(Al, m2)。使用體視顯微鏡拍攝莖段基部的橫截面,使用ImageJ測(cè)定木質(zhì)部面積(As, cm2)。葉面積邊材面積比(Al/As, m2/cm2)即為Al與As比值。
1.9 數(shù)據(jù)分析
所有分析均在R 4.1.3 (R Development Core Team, 2022)中進(jìn)行,所有圖片使用SigmaPlot 14.0 (Systat Software, San Jose, USA)繪制。使用R軟件中的“fitplc”包擬合脆弱性曲線。采用檢驗(yàn)(-test)比較抗風(fēng)桐和草海桐水力性狀的差異。
2 結(jié)果和分析
2.1 枝條與葉片脆弱性曲線
抗風(fēng)桐和草海桐的枝條和葉片脆弱性曲線均為“S型”曲線(圖1)。抗風(fēng)桐的葉片比枝條具有更低的木質(zhì)部導(dǎo)水率損失50%的水勢(shì)(即P50 leaf
2.2 枝條與葉片水力功能性狀
抗風(fēng)桐和草海桐的葉片膨壓?jiǎn)适c(diǎn)(tlp)、枝條邊材比導(dǎo)水率(Ks)和葉片導(dǎo)水率(Kleaf)均存在顯著差異(<0.001)(圖2)。抗風(fēng)桐和草海桐的tlp分別為–0.73和–1.44 MPa,草海桐比抗風(fēng)桐低49.3%;Ks分別為1.59和0.51 kg/(m·s·MPa),草海桐比抗風(fēng)桐低31.8%;草海桐具有比抗風(fēng)桐更高的Kleaf,分別為6.42和3.71 mmol/(m2·s·MPa),草海桐比抗風(fēng)桐高57.8%。
2.3 水力結(jié)構(gòu)性狀
對(duì)于邊材密度(WD)和葉面積邊材面積比(Al/ As),抗風(fēng)桐與草海桐之間均存在顯著差異(<0.05) (圖3)。與抗風(fēng)桐相比,草海桐具有更高的邊材密度,分別為0.28和0.38 g/cm3。另外,草海桐同樣擁有更高的葉面積邊材面積比,抗風(fēng)桐和草海桐的Al/As分別為0.33和0.51 m2/cm2。

圖1 兩種植物的枝條和葉片脆弱性曲線。PLC: 導(dǎo)水損失率; P50 branch: 枝條木質(zhì)部導(dǎo)水率損失50%的水勢(shì); P50 leaf: 葉片導(dǎo)水率損失50%的水勢(shì); ***:P<0.001。

圖2 兩種植物的水力功能性狀對(duì)比。ψtlp: 葉片膨壓?jiǎn)适c(diǎn); Ks: 枝條邊材比導(dǎo)水率; Kleaf: 葉片導(dǎo)水率; ***: P<0.001。
3 討論和結(jié)論
3.1 水力脆弱性分區(qū)策略
木質(zhì)部P50能夠作為判斷植物栓塞脆弱性的指標(biāo)[6]。本研究表明,抗風(fēng)桐葉片相較于枝條具有更低的P50,而草海桐枝條的P50較葉片更低,說明抗風(fēng)桐的葉片比草海桐更能抵抗栓塞,而枝條抗栓塞能力則稍弱。這兩物種的枝條P50比陸地生態(tài)系統(tǒng)木本植物的平均P50都高(熱帶季節(jié)性森林樹種枝條的P50均值約為–2.18 MPa[29]),表明在熱帶降雨較多的地區(qū),植物普遍不具有很強(qiáng)的耐旱能力,主要應(yīng)對(duì)季節(jié)性干旱或其他的脅迫環(huán)境條件。然而即使如此,2種植物也采取了截然不同的水分適應(yīng)策略,草海桐的枝條與葉片之間存在顯著的水力脆弱性分區(qū),表明其葉片能夠充當(dāng)“安全閥”來保護(hù)水力通路免受水力障礙的影響。因?yàn)椴莺M┤~片的提前栓塞能夠觸發(fā)氣孔關(guān)閉,降低蒸騰作用,從而降低其枝條和莖中木質(zhì)部的張力,有利于其適應(yīng)環(huán)境中高的飽和水汽壓差,這是其在惡劣的珊瑚島生存的重要策略[30]。抗風(fēng)桐則可能選擇以高效的木質(zhì)部運(yùn)水和儲(chǔ)水能力的策略應(yīng)對(duì)高蒸散環(huán)境可能造成的水分脅迫,并缺乏葉片的安全閥功能,這和前人[14]報(bào)道氣候濕潤地區(qū)的物種缺乏水力脆弱性分區(qū)的現(xiàn)象一致。作為補(bǔ)償,抗風(fēng)桐可能利用其他水力策略,如在土壤水分條件充足時(shí),利用較高的枝條導(dǎo)水效率使土壤到末端葉片的水勢(shì)梯度降低,從而避免枝條和葉片發(fā)生較大程度的栓塞[31]。此外,抗風(fēng)桐還可以利用多孔隙、低邊材密度的莖木質(zhì)部存儲(chǔ)水分,在面對(duì)環(huán)境干旱條件時(shí)能釋放足夠的水量來緩沖并維持植物水分平衡[32–33]。
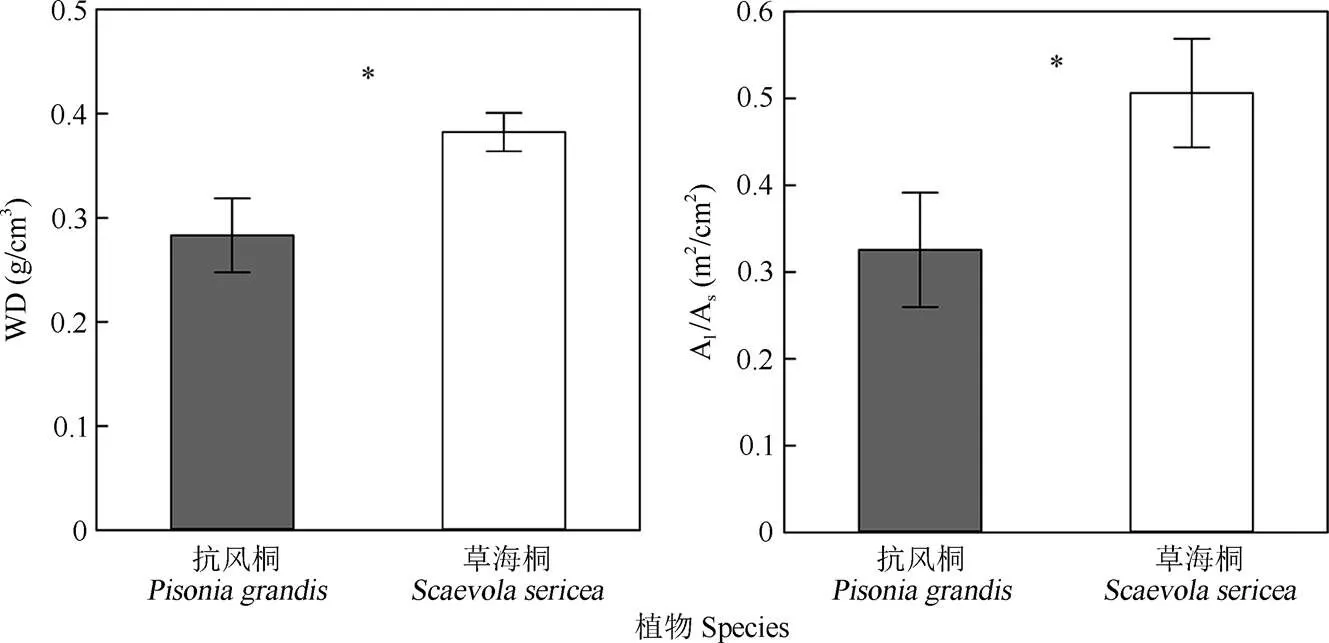
圖3 2種植物的水力結(jié)構(gòu)性狀對(duì)比。*: P<0.05; WD: 木材密度; Al/As: 葉面積邊材面積比。
3.2 水分運(yùn)輸策略
水分運(yùn)輸效率高的物種,可以保持葉片良好的水分狀況,從而保持氣孔開放,增加碳同化的時(shí)間,有助于植物的生長[33]。本研究表明,抗風(fēng)桐的葉片膨壓?jiǎn)适c(diǎn)(tlp)和枝條木質(zhì)部導(dǎo)水效率(Ks)顯著高于草海桐,而草海桐的葉片效率(Kleaf)則顯著高于抗風(fēng)桐,這表明草海桐作為灌木,選擇以較低的Ks減小枝葉的水分散失,使在根運(yùn)輸上的有限水分能夠在植株內(nèi)均勻分配,使植物體內(nèi)有充足的水分維持正常生長,同時(shí)以高的Kleaf來增強(qiáng)葉片水分運(yùn)輸能力以滿足熱帶珊瑚島高光高溫生境引起的高蒸騰要求,在此條件下保護(hù)葉片免受過熱造成的危害[34]。另一方面,草海桐tlp較低,Kleaf較高,表明其葉片可在更低的水勢(shì)下維持葉細(xì)胞膨壓,葉片關(guān)閉氣孔較晚,因此能維持更長時(shí)間的氣體交換[9,35]。雖然葉片中儲(chǔ)存的水分散失較快,但可以優(yōu)先栓塞部分葉片,降低蒸騰,以保護(hù)枝條等碳投資較高器官的水力安全,這也支持了水力脆弱性分區(qū)的觀點(diǎn)。
抗風(fēng)桐則選擇更高的枝條木質(zhì)部導(dǎo)水效率,這可能是由于抗風(fēng)桐作為喬木,根系可以在表層土壤干燥時(shí),從深層土壤獲得水分,從而可以將足量的水分通過枝條快速運(yùn)輸?shù)饺~片,以保持枝葉良好的水分狀況[35],但這需要進(jìn)一步的根系性狀對(duì)比研究證實(shí)。抗風(fēng)桐的tlp較高且葉片導(dǎo)水能力較低,這表明其葉片對(duì)干旱脅迫更加敏感,受到干旱脅迫后快速關(guān)閉氣孔,及時(shí)降低蒸騰速率來防止水分散失,同時(shí)葉片具有更高的抗栓塞能力,以此來應(yīng)對(duì)干旱[36]。
3.3 不同水分適應(yīng)策略的結(jié)構(gòu)基礎(chǔ)
邊材密度(WD)反映木質(zhì)部管道及其周邊包圍組織的性質(zhì),而葉面積邊材面積比(Al/As)則反映了單位枝干組織量可供水的葉面積大小[15–16]。本研究表明抗風(fēng)桐的WD和Al/As均顯著低于草海桐,但兩者的邊材密度均屬于熱帶地區(qū)木本植物中的較低水平(熱帶季節(jié)性森林樹種WD均值約為0.54 g/cm3[37])。前人[38]研究認(rèn)為,邊材密度與邊材水容通常有負(fù)的相關(guān)關(guān)系,這是因?yàn)榫哂懈咚莸闹参镏Ω蓳碛懈嗟谋”诮M織,能夠在遭遇水分脅迫時(shí)釋放薄壁細(xì)胞中的儲(chǔ)存水來暫時(shí)緩解木質(zhì)部張力,從而在一定程度上避免水力失敗。這兩種植物可能選擇以此方式來應(yīng)對(duì)一定程度的干旱脅迫,而不需要增加木質(zhì)部導(dǎo)管的抗栓塞能力,同時(shí)也可以獲得相對(duì)較高的生長速率以獲取競(jìng)爭(zhēng)優(yōu)勢(shì)[39]。此外,抗風(fēng)桐更低的Al/As表明其邊材向單位葉片面積的水分供應(yīng)能力更強(qiáng),因此其葉片可以獲得更大的蒸騰速率以提升單位面積的光合能力,但尚需更多光合作用相關(guān)數(shù)據(jù)證明這一推測(cè)。根據(jù)實(shí)地觀察,作為高大的喬木,抗風(fēng)桐更低的Al/As也是其在暴風(fēng)雨頻發(fā)的熱帶珊瑚島生存的一大優(yōu)勢(shì),由于冠層葉片非常稀疏,主要以交錯(cuò)的枝干形成屏障,有效避免了被強(qiáng)風(fēng)摧折。草海桐的單位枝干供水可以支撐更大的葉片面積, 這表明草海桐選擇增加碳分配給葉片以增加總光合面積獲取更強(qiáng)的競(jìng)爭(zhēng)力,而在面臨干旱脅迫時(shí),則允許損失部分葉片以保證枝干的水力安全性[14]。同時(shí)生活型為灌木的草海桐從地表到冠層均生長了緊密的葉片,選擇投資更多的碳到支撐組織,以換取更高的抵抗機(jī)械損傷的能力以及抗生物侵害能力, 從而應(yīng)對(duì)更多的葉面積可能帶來的潛在風(fēng)險(xiǎn)[40–41]。
綜上,中國熱帶珊瑚島植被典型適生木本植物抗風(fēng)桐和草海桐表現(xiàn)出不同的水分適應(yīng)策略。抗風(fēng)桐在熱帶地區(qū)高溫、高降雨、高光照的環(huán)境條件下,選擇以高效的枝干水分運(yùn)輸來滿足葉片高蒸騰需求的充足供水,同時(shí)葉片具有比枝條更強(qiáng)的抗栓塞能力,對(duì)水分脅迫更加敏感,能夠快速關(guān)閉氣孔, 降低蒸騰速率維持整體水分平衡。而草海桐在相同環(huán)境條件下,則選擇高效的葉片水分運(yùn)輸維持蒸騰, 降低葉溫以免受過熱傷害,葉片膨壓?jiǎn)适c(diǎn)較低,可在低水勢(shì)下維持葉片氣孔開放,同時(shí)葉片和枝條之間存在水力脆弱性分區(qū),葉片可充當(dāng)“安全閥”,在面臨水分脅迫時(shí)葉片首先栓塞,從而降低蒸騰, 以保證枝干的水力安全。這兩種植物均具有相對(duì)較低的木材密度,選擇以較高的生長速率獲得競(jìng)爭(zhēng)優(yōu)勢(shì),同時(shí)木質(zhì)部可在面臨干旱時(shí)釋放儲(chǔ)存水分緩解水分脅迫造成的影響。總之,通過采取不同的水分適應(yīng)策略,這兩種植物均能較好地適應(yīng)熱帶珊瑚島獨(dú)有的氣候條件,成為熱帶珊瑚島植被的優(yōu)勢(shì)樹種。本研究揭示了熱帶珊瑚島優(yōu)勢(shì)植物的兩種水分適應(yīng)策略,為熱帶珊瑚島植被修復(fù)工具種的篩選提供了一定的科學(xué)依據(jù)和理論參考。
[1] LI J, LIU N, REN H, et al. Ecological adaptability of seven plant species to tropical coral island habitat [J]. Ecol Environ Sci, 2016, 25 (5): 790–794. [李婕, 劉楠, 任海, 等. 7種植物對(duì)熱帶珊瑚島環(huán)境的生態(tài)適應(yīng)性 [J]. 生態(tài)環(huán)境學(xué)報(bào), 2016, 25(5): 790–794. doi: 10.16258/j. cnki.1674-5906.2016.05.009.]
[2] LIN Y X, LIU H, HE P C, et al. Physiological and biochemical responses of three species to environment stresses of tropical coral islands [J]. J Trop Subtrop Bot, 2017, 25(6): 562–568. [林憶雪, 劉慧, 賀鵬程, 等. 三種適生植物對(duì)熱帶珊瑚島脅迫生境的生理生化響應(yīng)[J]. 熱帶亞熱帶植物學(xué)報(bào), 2017, 25(6): 562–568. doi: 10.11926/ jtsb.3755.]
[3] YOU C J, HOU P X, DENG C F, et al. Investigation of tourism resources in Xisha Islands [J]. Resour Sci, 2015, 37(8): 1609–1620. [游長江, 侯佩旭, 鄧燦芳, 等. 西沙群島旅游資源調(diào)查與評(píng)價(jià) [J]. 資源科學(xué), 2015, 37(8): 1609–1620.]
[4] XU T Y, NIU X, WANG B. Recent advances in plant leaf hydraulic traits [J]. Terr Ecosyst Conserv, 2022, 2(2): 83–91. [許庭毓, 牛香, 王兵. 植物葉片水力性狀研究綜述[J]. 陸地生態(tài)系統(tǒng)與保護(hù)學(xué)報(bào), 2022, 2(2): 83–91. doi: 10.12356/j.2096-8884.2022-0005.]
[5] ZHU S D, CHEN Y J, CAO K F, et al. Interspecific variation in branch and leaf traits among threetree species from different successional tropical forests [J]. Funct Plant Biol, 2015, 42(4): 423– 432. doi: 10.1071/FP14201.
[6] CHOAT B, BRODRIBB T J, BRODERSEN C R, et al. Triggers of tree mortality under drought [J]. Nature, 2018, 558(7711): 531–539. doi: 10. 1038/s41586-018-0240-x.
[7] ZHOU H H, LI W H, AYUP Mubarek, et al. Xylem hydraulic conductivity and embolism properties of desert riparian forest plants and its response to drought stress [J]. Chin J Plant Ecol, 2012, 36(1): 19–29. [周洪華, 李衛(wèi)紅, 木巴熱克·阿尤普, 等. 荒漠河岸林植物木質(zhì)部導(dǎo)水與栓塞特征及其對(duì)干旱脅迫的響應(yīng)[J]. 植物生態(tài)學(xué)報(bào), 2012, 36(1): 19–29.doi: 10.3724/SP.J.1258.2012.00019.]
[8] BARTLETT M K, SCOFFONI C, SACK L. The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: A global meta-analysis [J]. Ecol Lett, 2012, 15(5): 393–405. doi: 10.1111/j.1461-0248.2012.01751.x.
[9] BLACKMAN C J, BRODRIBB T J, JORDAN G J. Leaf hydraulic vulnerability is related to conduit dimensions and drought resistance across a diverse range of woody angiosperms [J]. New Phytol, 2010, 188(4): 1113–1123. doi: 10.1111/j.1469-8137.2010.03439.x.
[10] GLEASON S M, WESTOBY M, JANSEN S, et al. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species [J]. New Phytol, 2016, 209(1): 123–136. doi: 10.1111/nph.13646.
[11] TYREE M T, EWERS F W. The hydraulic architecture of trees and other woody plants [J]. New Phytol, 1991, 119(3): 345–360. doi: 10. 1111/j.1469-8137.1991.tb00035.x.
[12] BUCCI S J, SCHOLZ F G, CAMPANELLO P I, et al. Hydraulic differences along the water transport system of South Americanspecies: Do leaves protect the stem functionality? [J]. Tree Physiol, 2012, 32(7): 880–893. doi: 10.1093/treephys/tps054.
[13] WHITEHEAD D, JARVIS P G. Coniferous forests and plantations [M]// KOZLOWSKI T T. Water Deficits and Plant Growth, VI. New York: Academic Press, 1981: 49–152.
[14] ZHU S D, LIU H, XU Q Y, et al. Are leaves more vulnerable to cavitation than branches? [J]. Funct Ecol, 2016, 30(11): 1740–1744. doi: 10.1111/1365-2435.12656.
[15] CHAVE J, COOMES D, JANSEN S, et al. Towards a worldwide wood economics spectrum [J]. Ecol Lett, 2009, 12(4): 351–366. doi: 10.1111/j. 1461-0248.2009.01285.x.
[16] MENCUCCINI M, MANZONI S, CHRISTOFFERSEN B. Modelling water fluxes in plants: From tissues to biosphere [J]. New Phytol, 2019, 222(3): 1207–1222. doi: 10.1111/nph.15681.
[17] REN H, JIAN S G, ZHANG Q M, et al. Plants and vegetation on South China Sea Islands [J]. Ecol Environ Sci, 2017, 26(10): 1639–1648. [任海, 簡(jiǎn)曙光, 張倩媚, 等. 中國南海諸島的植物和植被現(xiàn)狀[J]. 生態(tài)環(huán)境學(xué)報(bào), 2017, 26(10): 1639–1648. doi: 10.16258/j.cnki.1674- 5906.2017.10.001.]
[18] YU X C, LIANG H Z, CHEN S Y, et al. Multiple shoot proliferation and plant regeneration in[J]. Guihaia, 2021, 41(6): 890–896. [于昕塍, 梁韓枝, 陳雙艷, 等. 抗風(fēng)桐的叢生芽誘導(dǎo)與再生 [J]. 廣西植物, 2021, 41(6): 890–896. doi: 10.11931/guihaia.gxzw 202004015.]
[19] WANG J. Breeding and mixed planting of local tree speciesandin coastal shelterbelt on Hainan Island [D]. Haikou: Hainan Normal University, 2015. [王瑾. 海南島海岸鄉(xiāng)土樹種紅厚殼、草海桐的育苗和在海防林下混交種植的研究 [D]. 海口: 海南師范大學(xué), 2015.]
[20] XU B B. Study on drought and saline-alkaline resistance of[D]. Guangzhou: Zhongkai University of Agriculture and Engineering, 2019. [徐貝貝. 草海桐的抗干旱和鹽堿性研究 [D]. 廣州: 仲愷農(nóng)業(yè)工程學(xué)院, 2019.]
[21] XU B B, LIU N, REN H, et al. Stress resistance biological character- ristics ofin Paracel Islands [J]. Guihaia, 2018, 38(10): 1277–1285. [徐貝貝, 劉楠, 任海, 等. 西沙群島草海桐的抗逆生物學(xué)特性 [J]. 廣西植物, 2018, 38(10): 1277–1285. doi: 10.11931/ guihaia.gxzw201711012.]
[22] HAN S S, LIU S X, MO X G, et al. Soil water dynamics and water balance on a tropical coral island [J]. Hydrol Process, 2021, 35(12): e14415. doi: 10.1002/hyp.14415.
[23] WANG C, ZHANG H, LIU H, et al. Application of a trait-based species screening framework for vegetation restoration in a tropical coral island of China [J]. Funct Ecol, 2020, 34(6): 1193–1204. doi: 10.1111/1365-2435.13553.
[24] PAMMENTER N W, VAN DER WILLIGEN C. A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation [J]. Tree Physiol, 1998, 18(8–9): 589–593. doi: 10.1093/ treephys/18.8-9.589.
[25] EWERS F W, FISHER J B. Techniques for measuring vessel lengths and diameters in stems of woody plants [J]. Am J Bot, 1989, 76(5): 645–656. doi: 10.1002/j.1537-2197.1989.tb11360.x.
[26] TYREE M T, HAMMEL H T. The measurement of the turgor pressure and the water relations of plants by the pressure-bomb technique [J]. J Exp Bot, 1972, 23(1): 267–282. doi: 10.1093/jxb/23.1.267.
[27] SCHULTE P J, HINCKLEY T M. A comparison of pressure-volume curve data analysis techniques [J]. J Exp Bot, 1985, 36(10): 1590–1602. doi: 10.1093/jxb/36.10.1590.
[28] BRODRIBB T J, HOLBROOK N M. Changes in leaf hydraulic conductance during leaf shedding in seasonally dry tropical forest [J]. New Phytol, 2003, 158(2): 295–303. doi: 10.1046/j.1469-8137.2003. 00736.x.
[29] LIU H, GLEASON S M, HAO G Y, et al. Hydraulic traits are coordinated with maximum plant height at the global scale [J]. Sci Adv, 2019, 5(2): eaav1332. doi: 10.1126/sciadv.aav1332.
[30] TYREE M T, COCHARD H, CRUIZIAT P, et al. Drought-induced leaf shedding in walnut: Evidence for vulnerability segmentation [J]. Plant Cell Environ, 1993, 16(7): 879–882. doi: 10.1111/j.1365-3040.1993. tb00511.x.
[31] BUCCI S J, SCHOLZ F G, GOLDSTEIN G, et al. Soil water availa- bility and rooting depth as determinants of hydraulic architecture of Patagonian woody species [J]. Oecologia, 2009, 160(4): 631–641. doi: 10.1007/s00442-009-1331-z.
[32] MCCULLOH K A, JOHNSON D M, MEINZER F C, et al. The dynamic pipeline: Hydraulic capacitance and xylem hydraulic safety in four tall conifer species [J]. Plant Cell Environ, 2014, 37(5): 1171–1183.doi: 10.1111/pce.12225.
[33] GUYOT G, SCOFFONI C, SACK L. Combined impacts of irradiance and dehydration on leaf hydraulic conductance: Insights into vulnera- bility and stomatal control [J]. Plant Cell Environ, 2012, 35(5): 857– 871. doi: 10.1111/j.1365-3040.2011.02458.x.
[34] PINEDA-GARCIA F, PAZ H, MEINZER F C. Drought resistance in early and late secondary successional species from a tropical dry forest: The interplay between xylem resistance to embolism, sapwood water storage and leaf shedding [J]. Plant Cell Environ, 2013, 36(2): 405–418.doi: 10.1111/j.1365-3040.2012.02582.x.
[35] SCOFFONI C, MCKOWN A D, RAWLS M, et al. Dynamics of leaf hydraulic conductance with water status: Quantification and analysis of species differences under steady state [J]. J Exp Bot, 2012, 63(2): 643– 658. doi: 10.1093/jxb/err270.
[36] ARITSARA A N A, WANG S, LI B N, et al. Divergent leaf and fine root “pressure-volume relationships” across habitats with varying water availability [J]. Plant Physiol, 2022, 190(4): 2246–2259. doi: 10.1093/ plphys/kiac403.
[37] LI R H, ZHU S D, CHEN H Y H, et al. Are functional traits a good predictor of global change impacts on tree species abundance dynamics in a subtropical forest? [J]. Ecol Lett, 2015, 18(11): 1181–1189. doi: 10.1111/ele.12497.
[38] MEINZER F C, JOHNSON D M, LACHENBRUCH B, et al. Xylem hydraulic safety margins in woody plants: Coordination of stomatal control of xylem tension with hydraulic capacitance [J]. Funct Ecol, 2009, 23(5): 922–930. doi: 10.1111/j.1365-2435.2009.01577.x.
[39] ZANNE A E, WESTOBY M, FALSTER D S, et al. Angiosperm wood structure: Global patterns in vessel anatomy and their relation to wood density and potential conductivity [J]. Am J Bot, 2010, 97(2): 207–215. doi: 10.3732/ajb.0900178.
[40] CLARK D A, CLARK D B. Getting to the canopy: Tree height growth in a neotropical rain forest [J]. Ecology, 2001, 82(5): 1460–1472. doi: 10.1890/0012-9658(2001)082[1460:GTTCTH]2.0.CO;2.
[41] FALSTER D S, BR?NNSTR?M ?, DIECKMANN U, et al. Influence of four major plant traits on average height, leaf‐area cover, net primary productivity, and biomass density in single‐species forests: A theoretical investigation [J]. J Ecol, 2011, 99(1): 148–164. doi: 10. 1111/j.1365-2745.2010.01735.x.
Hydraulic Adaptation Strategies of Dominant Woody Plantsandon Tropical Coral Islands of China
TANG Weize1,2,3, LI Qin1,2,3, ZHANG Haoping1,2,3, JIN Yi1,3, LI Qiang4, YIN Deyi1,3, YE Qing1,3, LIU Hui1,3*
(1. Guangdong Provincial Key Laboratory of Applied Botany, Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650, China; 2. University of Chinese Academy of Sciences,Beijing 100049, China; 3.South China National Botanical Garden, Guangzhou 510650, China; 4. School of Tropical Medicine, Hainan Medical University, Haikou 571199, China)
Plant hydraulic traits can reflect the adaptability of plants to different water conditions. It is important to study the hydraulic traits of dominant plants in the special habitat of tropical coral islands for a deep understanding of the hydraulic adaptation strategies of tropical coral island plants, and for selecting suitable tree species for tropical coral island vegetation construction and restoration. Thus, the leaf and branch hydraulic traits of two representative dominant tree species in the habitat of tropical coral islands of China,,and, were measured, and their hydraulic adaptation strategies were compared and analyzed. The results showed that the leaf embolism resistance, leaf turgor point and branch specific xylem conductivity of.were significantly higher than those of., but the values of branch embolism resistance, leaf hydraulic conductance, sapwood density and leaf to stem area ratio were significantly lower than those of.. Furthermore, leaves of.were more resistant to embolism than branches, and were sensitive to water stress. Meanwhile, high water transport efficiency in branches of.provided sufficient water to ensure the high transpiration of the leaves. On the other hand, there was a significant hydraulic vulnerability segmentation between leaves and branches of.. The leaves of.could act as “safety valves” to protect branch hydraulic pathway from dysfunction.and.could adapt to the special habitats of tropical coral islands by coordinating the water transport efficiency and safety of leaves and branches and adopting different hydraulic strategies.
Hydraulic vulnerability segmentation; Plant hydraulic trait; Embolism; Tropical coral island plant; Water transport
10.11926/jtsb.4797
2023-04-19
2023-05-15
國家重點(diǎn)研發(fā)計(jì)劃項(xiàng)目(2021YFC3100401);國家自然科學(xué)基金項(xiàng)目(32371575);中國科學(xué)院華南植物園青年人才專項(xiàng)(QNXM-01);中國科學(xué)院重點(diǎn)部署項(xiàng)目(KGFZD-135-19-08)資助
This work was supported by the National Key R&D Program of China (Grant No. 2021YFC3100401), the National Natural Science Foundation of China (Grant No. 32371575), the Special Project for Youth Talent of South China Botanical Garden, Chinese Academy of Sciences (Grant No. QNXM-01), and the Key Research Program of Chinese Academy of Sciences (Grant No. KGFZD-135-19-08).
唐瑋澤(1998年生),男,碩士研究生,研究方向?yàn)橹参锷砩鷳B(tài)學(xué)。E-mail: tangweize@scbg.ac.cn
* 通訊作者 Corresponding author. E-mail: hui.liu@scbg.ac.cn
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
