基于最大熵模型的中國(guó)兜蘭屬植物潛在分布模擬
2024-02-02 06:18:22黃智聰舒江平嚴(yán)岳鴻陳建兵
熱帶亞熱帶植物學(xué)報(bào) 2024年1期
黃智聰, 舒江平,2, 嚴(yán)岳鴻,陳建兵*
基于最大熵模型的中國(guó)兜蘭屬植物潛在分布模擬
黃智聰1, 舒江平1,2, 嚴(yán)岳鴻1,陳建兵1*
(1. 深圳市蘭科植物保護(hù)研究中心,廣東 深圳 518114,2. 中國(guó)科學(xué)院華南植物園, 廣州 510650)
基于已知分布點(diǎn)和20個(gè)環(huán)境因子,該研究利用MaxEnt模型模擬在現(xiàn)在(1970—2000年)氣候條件和2種不同共享經(jīng)濟(jì)路徑情景下(SSP1-2.6、SSP5-8.5)未來(lái)(2081—2100年)兜蘭屬()植物的潛在分布格局,找出影響物種分布的環(huán)境因子。結(jié)果表明,兜蘭屬植物的最適宜分布區(qū)位于滇東南地區(qū)、貴州西南、廣西西部、廣東南部、海南北部。影響該屬植物分布的主要環(huán)境因子是年降水量、年溫度變化和最干旱季降水量。隨著全球變暖,適生區(qū)有向北和西北方向擴(kuò)張的趨勢(shì),逐漸往西北亞熱帶方向延伸。在SSP5-8.5的情景下,高適生區(qū)出現(xiàn)大幅度收縮。在未來(lái)氣候情景下,不同種群的分布區(qū)變化規(guī)律并不一致,其分布格局響應(yīng)氣候變化的趨勢(shì)也有所不同,因此該文針對(duì)分布區(qū)變化趨勢(shì)不同的物種提出了不同的保護(hù)策略。
兜蘭屬;最大熵模型;氣候因子;氣候變化
物種的分布區(qū)是物種重要的空間特征,與物種滅絕、生態(tài)入侵、生態(tài)位幅度密切相關(guān),對(duì)物種的起源、擴(kuò)散和演化研究具有重要意義[1]。在全球氣候變化的背景下,進(jìn)行物種潛在分布區(qū)的模擬和主導(dǎo)因子分析,可以為有效保護(hù)和可持續(xù)利用植物資源提供科學(xué)依據(jù)。氣候是影響物種自然地理分布最重要的因素之一,全球氣候變化對(duì)物種的分布格局和生物多樣性產(chǎn)生顯著影響[2–3]。氣候變化導(dǎo)致的物種棲息地喪失和破碎化已經(jīng)嚴(yán)重威脅到自然分布范圍狹小的瀕危物種,加劇其滅絕風(fēng)險(xiǎn)[4–5]。在過(guò)去的100年里,全球平均氣溫上升約0.85 ℃,且預(yù)測(cè)未來(lái)氣溫仍將持續(xù)快速上升,未來(lái)冬季變暖幅度將大于春季[6]。全球變暖導(dǎo)致植物物候發(fā)生顯著變化, 冬季升溫導(dǎo)致開(kāi)花-展葉時(shí)間間隔延長(zhǎng),但春季升溫導(dǎo)致開(kāi)花-展葉時(shí)間間隔縮短,這種季節(jié)性升溫幅度的差異可能導(dǎo)致開(kāi)花-展葉時(shí)間間隔向不同的方向改變,改變植物的營(yíng)養(yǎng)分配、繁殖以及種間關(guān)系,并影響生態(tài)系統(tǒng)的結(jié)構(gòu)和功能,進(jìn)而影響生態(tài)系統(tǒng)碳循環(huán)、水循環(huán)和能量平衡[7]。研究植物適宜生境對(duì)氣候變化的響應(yīng),對(duì)于展現(xiàn)物種形成、遷移的變化,提出合理有效的種質(zhì)資源保護(hù)對(duì)策,有著重要的理論與現(xiàn)實(shí)意義。物種分布模型[8]是利用物種的分布數(shù)據(jù),并結(jié)合其生境下的環(huán)境變量推測(cè)物種的基礎(chǔ)生態(tài)位, 再結(jié)合不同時(shí)期的環(huán)境數(shù)據(jù)模擬不同時(shí)期物種的潛在分布區(qū)。其中,由美國(guó)學(xué)者Phillips等[9]開(kāi)發(fā)的最大熵法(MaxEnt)生態(tài)位模型具有建模快、精度高等優(yōu)點(diǎn),即便較小的樣本量也能獲得具有較高精度和穩(wěn)定性的模型[10]。朱瑩瑩等[11]研究了我國(guó)特有珍稀植物水杉()種群分布對(duì)氣候變化的響應(yīng),結(jié)果表明,在不同氣候情境下我國(guó)水杉生境有較低的適宜程度。張佳琦等[12]研究了裸果木()分布格局的變化,認(rèn)為在未來(lái)氣候情景下其適生區(qū)明顯縮小。Hu等[13]分析了側(cè)柏()在未來(lái)氣候變化下的潛在分布,結(jié)果表明側(cè)柏的分布可能會(huì)隨著氣候的變暖而增加。不同物種的采樣分布不同和影響物種分布的環(huán)境因素不同, 其結(jié)果也有差異。然而蘭科植物的地理分布格局如何響應(yīng)未來(lái)氣候變化仍知之甚少。
兜蘭屬()是蘭科植物最具觀賞價(jià)值的類群之一,是熱帶及亞熱帶地區(qū)的林下多年生草本植物,大多數(shù)生于地面或積土的巖石表面或巖縫中,較少數(shù)著生于樹(shù)上,主要生存在排水、通風(fēng)好,有一定蔭蔽,又能透光而且濕潤(rùn)的環(huán)境下。全世界約有79種,主要分布于東南亞至喜馬拉雅山低地和中國(guó)西南部,只有少數(shù)物種到達(dá)新幾內(nèi)亞和所羅門群島[14]。我國(guó)兜蘭屬植物資源非常豐富,約占全世界的三分之一,但由于過(guò)度采集、走私出境以及生境破壞等原因,近20年來(lái)其數(shù)量急劇減少, 瀕臨滅絕[15–18]。目前,兜蘭屬所有野生種均被列入《國(guó)家重點(diǎn)保護(hù)野生植物(第二批)》名錄和《瀕危野生動(dòng)植物種國(guó)際貿(mào)易公約》(CITES)附錄Ⅰ中。然而, 兜蘭屬的生存狀況及其適宜分布區(qū)對(duì)未來(lái)氣候變化的響應(yīng)尚未得到深入研究。
在未來(lái)不同氣候情景下,兜蘭屬植物的地理分布格局將呈現(xiàn)怎樣的變化?影響分布格局的主導(dǎo)氣候因子是否隨物種的不同而變化?兜蘭屬的保護(hù)策略該如何制定?本研究采用MaxEnt模型與GIS技術(shù)相結(jié)合的方法,根據(jù)物種資源分布信息, 結(jié)合當(dāng)代和未來(lái)氣候環(huán)境數(shù)據(jù),研究其在我國(guó)的潛在適生分布區(qū)及其主導(dǎo)氣候因子,以期為兜蘭屬植物的野外調(diào)查、分類學(xué)與生物地理學(xué)研究提供理論依據(jù),同時(shí)為兜蘭屬植物的保護(hù)提供參考。
1 材料和方法
1.1 數(shù)據(jù)收集與處理
1.1.1 物種分布信息
依據(jù)《中國(guó)植物志》[19]、《中國(guó)兜蘭屬植物》[14]、文獻(xiàn)以及標(biāo)本記錄記載的分布和性狀描述,查閱兜蘭屬植物的歷史分布點(diǎn),記錄物種的海拔、生境類型等信息。
物種分布信息來(lái)源于3個(gè)方面:一是根據(jù)蘭科中心的野外調(diào)查數(shù)據(jù),分布點(diǎn)經(jīng)緯度信息由GPS實(shí)地定位獲得;二是通過(guò)查閱文獻(xiàn),找出其分布地點(diǎn),用百度拾取坐標(biāo)查找相應(yīng)的經(jīng)緯度坐標(biāo);三是通過(guò)中國(guó)數(shù)字植物標(biāo)本館(http://www.cvh.org.cn/)和全球生物多樣性信息網(wǎng)絡(luò)(https://www.gbif.org/)獲取標(biāo)本采集地經(jīng)緯度信息。
《中國(guó)物種名錄》(2022版)[20]記載中國(guó)兜蘭屬包含30種,但在數(shù)據(jù)收集過(guò)程中,根莖兜蘭()、紫斑兜蘭()、天倫兜蘭()和文山兜蘭()等4種由于分布信息單一或未收集到縣級(jí)以下的具體分布點(diǎn),因此未納入計(jì)算。本研究列入兜蘭屬模型計(jì)算的物種為26種,分別為卷萼兜蘭()、杏黃兜蘭()、小葉兜蘭()、巨瓣兜蘭()、紅旗兜蘭()、同色兜蘭()、德氏兜蘭()、長(zhǎng)瓣兜蘭()、白花兜蘭()、紅花兜蘭()、瑰麗兜蘭()、廣東兜蘭()、綠葉兜蘭()、巧花兜蘭()、亨利兜蘭()、帶葉兜蘭()、波瓣兜蘭()、麻栗坡兜蘭()、硬葉兜蘭()、飄帶兜蘭()、紫紋兜蘭()、白旗兜蘭()、虎斑兜蘭()、秀麗兜蘭()、紫毛兜蘭()和彩云兜蘭()。
為了保證數(shù)據(jù)的準(zhǔn)確性,對(duì)收集到的物種分布數(shù)據(jù)進(jìn)行過(guò)濾處理。去除人工試驗(yàn)樣地及人工溫室栽培的條目,去除重復(fù)的標(biāo)本條目, 以及經(jīng)緯度和小地點(diǎn)信息都沒(méi)有的條目,去掉分布信息與《中國(guó)植物志》記載明顯不符的條目以及物種鑒定有誤的分布數(shù)據(jù)。對(duì)所有數(shù)據(jù)按照MaxEnt模型要求進(jìn)行標(biāo)準(zhǔn)化處理,并且為了防止在進(jìn)行最大熵模型分析時(shí)出現(xiàn)過(guò)擬合現(xiàn)象,將兜蘭屬所有物種的分布數(shù)據(jù)進(jìn)行篩選,在2.5 km2以內(nèi)的物種分布點(diǎn)僅保留1個(gè),共計(jì)得到274個(gè)具有經(jīng)緯度信息的兜蘭屬物種分布點(diǎn)。另外,由于經(jīng)過(guò)篩選后部分物種的種群分布點(diǎn)過(guò)于稀少和集中,因此本次研究中僅對(duì)14物種進(jìn)行了最大熵模型分析。
1.1.2 氣候環(huán)境數(shù)據(jù)
IRF2是IFN信號(hào)通路的重要組成成分,在其信號(hào)通路中IRF2與其它調(diào)節(jié)因子共同調(diào)控細(xì)胞周期,在對(duì)腫瘤的調(diào)控中具有不可忽視的作用,通過(guò)對(duì)IRF2結(jié)構(gòu)和功能的研究,我們發(fā)現(xiàn),IRF2對(duì)免疫細(xì)胞的增值具有促進(jìn)作用,可以通過(guò)提高IRF2的表達(dá)水平來(lái)增強(qiáng)機(jī)體抵御外界病毒的能力,從而減少機(jī)體患病的可能,并在某些疾病發(fā)生之初就能有效的應(yīng)對(duì)。而且IRF2與IRF1具有競(jìng)爭(zhēng)性抑制作用,可以利用這一點(diǎn)對(duì)癌癥進(jìn)行抑制,但是由于其對(duì)癌癥的雙重作用,我們可以嘗試?yán)玫鞍椎臉?gòu)象改變對(duì)其進(jìn)行進(jìn)一步研究,以希望能盡早的征服癌癥,研發(fā)出對(duì)癌癥有效的藥物。
本研究選取當(dāng)代(1970—2000年)和未來(lái)(2081— 2100年)的20個(gè)環(huán)境數(shù)據(jù)(表1),均來(lái)源于世界氣候數(shù)據(jù)庫(kù)WorldClim (http://worldclim.org), 空間分辨率為2.5 min。當(dāng)代氣候數(shù)據(jù)是根據(jù)1970—2000年全球不同氣象站點(diǎn)記錄的氣候數(shù)據(jù)經(jīng)插值法生成。未來(lái)(2081—2100年)氣候數(shù)據(jù)選擇CMIP6計(jì)劃中精度為2.5 min的BCC-CSM2-MR模式下的兩組極端的共享經(jīng)濟(jì)路徑:SSP1-2.6和SSP5-8.5兩組數(shù)據(jù)。SSP1-2.6描述了1個(gè)全球二氧化碳排放量在2050年左右被削減到凈零的世界,到本世紀(jì)末溫度穩(wěn)定比世紀(jì)初高1.8 ℃;SSP5-8.5描述目前的二氧化碳排放水平到2050年大約翻一番,到2100年,二氧化碳排放水平將降低至與世紀(jì)前葉接近, 全球平均氣溫將比世紀(jì)初高4.4 ℃[21]。數(shù)據(jù)經(jīng)ArcGIS 10.6軟件剪裁為中國(guó)范圍并轉(zhuǎn)換為ASCII格式。
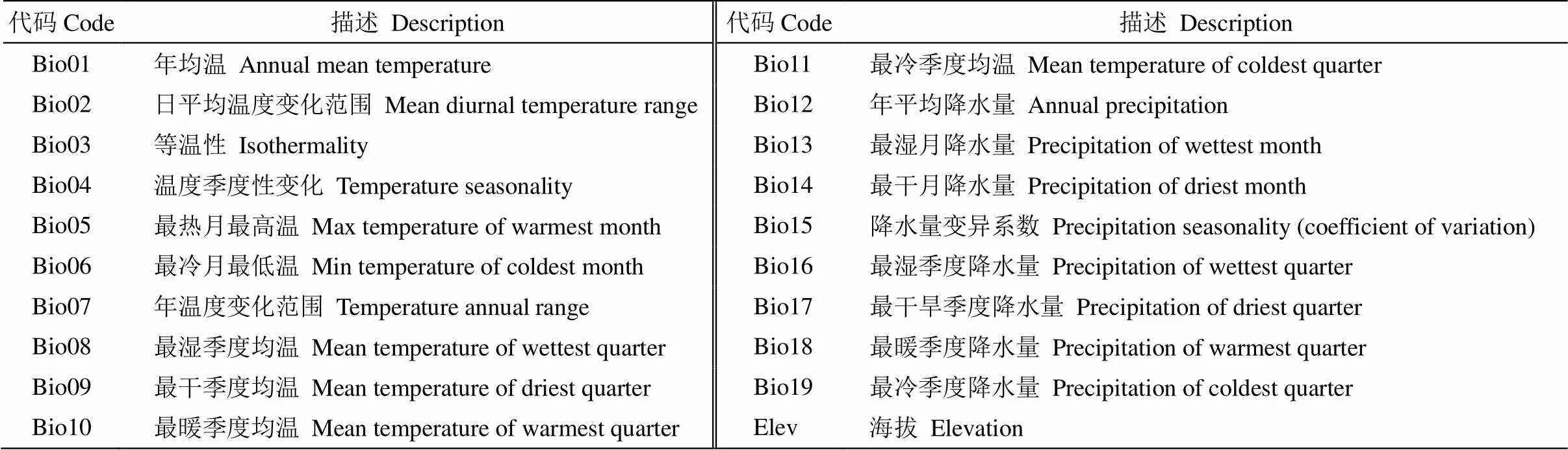
表1 環(huán)境變量及其描述
1.2 方法
1.2.1 建立模型
采用MaxEnt Version 3.4.4 (http://biodiversity-infor matics.amnh.org/open_source/maxent)模擬兜蘭屬植物在不同氣候情境下的潛在分布格局,設(shè)置訓(xùn)練集為75%,測(cè)試集為25%進(jìn)行模擬分析。將物種分布數(shù)據(jù)與氣候環(huán)境數(shù)據(jù)一起導(dǎo)入MaxEnt中,選擇刀切法,繪制響應(yīng)曲線并制作預(yù)測(cè)圖,所使用的底圖為自然資源部監(jiān)制的GS(2019)1823號(hào)中國(guó)地圖。
1.2.2 模型精度檢驗(yàn)
采用受試者工作特征曲線(receiver operator characteristic curves, ROC)對(duì)模型的精度進(jìn)行評(píng)價(jià)。ROC曲線與橫坐標(biāo)所圍成區(qū)域面積(AUC)的大小具有不受臨界值影響的特性,可用于評(píng)價(jià)預(yù)測(cè)模型的準(zhǔn)確性[22–24]。
利用刀切法測(cè)試可以評(píng)估環(huán)境變量對(duì)模型的影響程度,從而判斷不同環(huán)境變量對(duì)生態(tài)布局的重要性,其環(huán)境因子貢獻(xiàn)率是評(píng)估環(huán)境變量對(duì)模型影響程度的重要指標(biāo),多數(shù)研究者將累積貢獻(xiàn)率超過(guò)某一特定值之前的氣候因子作為影響物種分布的主導(dǎo)因子,該特定值通常根據(jù)各自研究物種的特性和結(jié)果主觀選擇。本文中將累計(jì)貢獻(xiàn)率超過(guò)70%時(shí)參與累積的因子作為主導(dǎo)因子,其中貢獻(xiàn)率大于10%的為最關(guān)鍵因子[25]。通過(guò)利用單一的變量而不考慮其他變量的模型生成的單變量響應(yīng)曲線可估算物種的適宜生存環(huán)境的范圍[26]。
1.2.3 適生區(qū)等級(jí)的劃分
MaxEnt結(jié)果輸出文件為ASCII格式,將其加載到ArcGIS 10.6中,使用“ArcToolbox”中的“格式轉(zhuǎn)換工具”轉(zhuǎn)換為柵格數(shù)據(jù),利用“重分類工具”將兜蘭屬植物的生境適宜性按照“自然間斷點(diǎn)分級(jí)法”劃分成4類,即非適生區(qū)、低適生區(qū)、中適生區(qū)和高適生區(qū),并計(jì)算各適生區(qū)的面積及其占比。
2 結(jié)果和分析
2.1 模型準(zhǔn)確性檢驗(yàn)
運(yùn)用MaxEnt模型基于20個(gè)環(huán)境變量模擬并重復(fù)10次運(yùn)算,兜蘭屬植物的潛在分布區(qū)AUC值為0.868~0.998 (圖1)。
2.2 影響兜蘭屬植物分布的主導(dǎo)氣候因子
最大熵模型模擬結(jié)果表明,對(duì)兜蘭屬地理分布影響較大的氣候因子有最干旱季降水量、年降水量、年溫度變化范圍、最冷季降水量。為了減少氣候因子之間的自相關(guān),利用ArcGis軟件提取兜蘭屬物種274個(gè)分布點(diǎn)的氣候變量,利用SPSS26軟件進(jìn)行相關(guān)性分析,保留相關(guān)系數(shù)<|0.8|的因子,對(duì)于相關(guān)系數(shù)≥|0.8|的,保留貢獻(xiàn)率更高的1個(gè),最后篩選出7個(gè)氣候因子(年平均溫度、日溫度平均變化范圍、等溫線、年溫度變化范圍、年降水量、降水量變異系數(shù)、最干旱季的降水量),用篩選過(guò)的環(huán)境因子進(jìn)行分析預(yù)測(cè),結(jié)果顯示當(dāng)前氣候情景下對(duì)兜蘭屬植物全屬地理分布影響排名前三的氣候因子仍為年降水量34%、年溫度變化范圍24.3%、最干旱季度降水量22.2%,其貢獻(xiàn)率綜合達(dá)到80.5%。刀切法檢驗(yàn)結(jié)果表明(圖2),變量單獨(dú)使用時(shí),年溫度變化范圍在受試變量中測(cè)試增益最突出;省略變量時(shí),減少增益最多的環(huán)境變量是降水量變異系數(shù)。
通過(guò)兜蘭屬因子的單變量響應(yīng)曲線(圖3),若以物種存在概率0.5為最適閥值,年溫度的變化為10 ℃~23 ℃時(shí),物種的存在概率基本保持在0.5~0.7的區(qū)域,說(shuō)明該溫差范圍是適合物種生存的舒適區(qū);當(dāng)年溫差在20 ℃~22 ℃時(shí),物種的存在概率達(dá)到峰值0.7,說(shuō)明這個(gè)溫差對(duì)物種而言是一種最適宜的生境溫度;年溫差超過(guò)22 ℃時(shí),物種的存在概率就逐漸降低。最干旱降水量在0~80 mm時(shí)存在概率呈上升趨勢(shì),超過(guò)80 mm之后存在概率逐漸降低,在50~120 mm區(qū)間時(shí)物種存在概率處于0.5~0.6區(qū)域,說(shuō)明該降水量范圍是適合兜蘭屬物種生存的舒適區(qū),并且最干旱季降水量在80 mm時(shí)存在概率達(dá)到峰值0.66,說(shuō)明該降水量是一種最適宜的生境,當(dāng)最干旱降水量超過(guò)120 mm時(shí),物種存在概率開(kāi)始降低。年降水量在1 200~2 400 mm區(qū)域時(shí)存在概率均處于0.5~0.7區(qū)域,說(shuō)明該降水量區(qū)間是適合物種生存的舒適區(qū);年降水量超過(guò)2 000 mm后存在概率逐漸下降。
14種植物的最大熵模型模擬結(jié)果還表明, 大部分兜蘭屬物種的分布同時(shí)受到多個(gè)環(huán)境因子影響(圖4), 其中最干旱季降水量、最暖季降水量、年降水量和年溫度變化范圍對(duì)大多數(shù)兜蘭屬物種的分布有顯著影響。少數(shù)物種分布的主導(dǎo)環(huán)境因子(累積貢獻(xiàn)率大于70%)僅為單、雙氣候因子,卷萼兜蘭主要受年溫度變化范圍影響、秀麗兜蘭主要受等溫性影響,杏黃兜蘭、格力兜蘭主要受等溫性和溫度季節(jié)性變化影響,巧花兜蘭主要受年溫度變化范圍和最冷月最低溫度的影響,麻栗坡兜蘭主要受年溫度變化范圍和最干燥月降水量的影響。
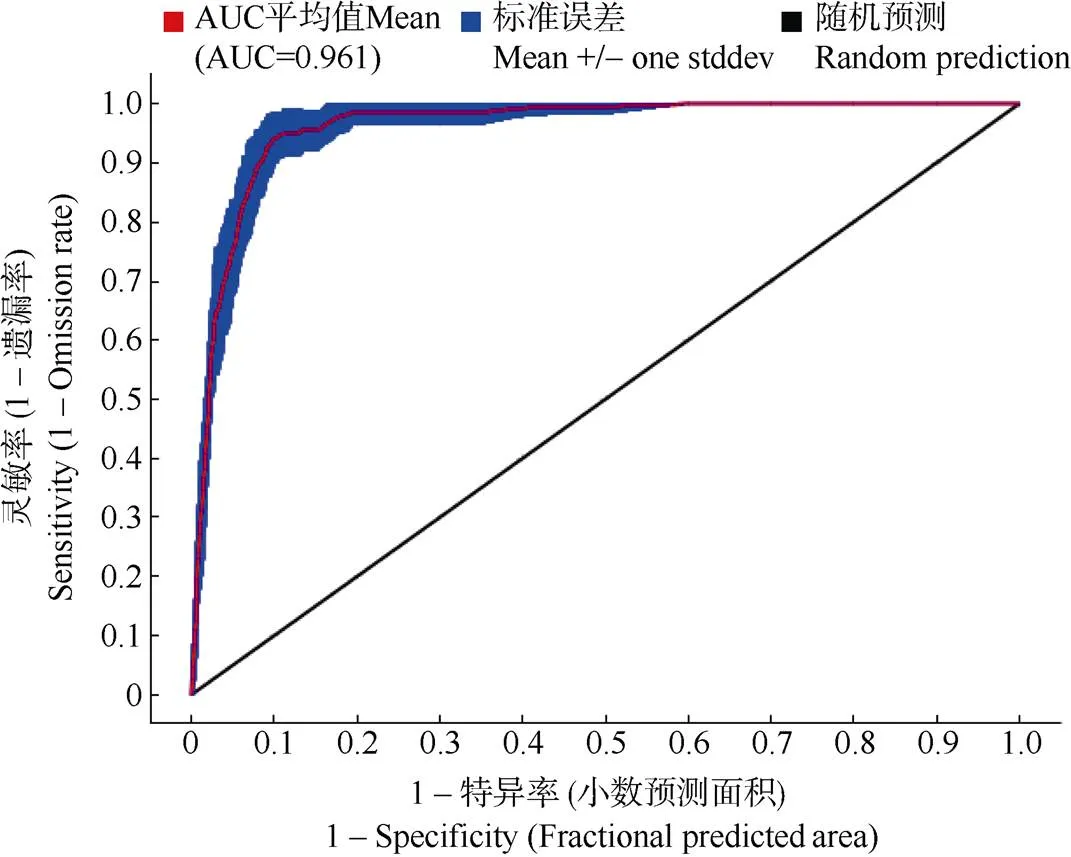
圖1 兜蘭屬當(dāng)前氣候條件下分布預(yù)測(cè)的ROC曲線
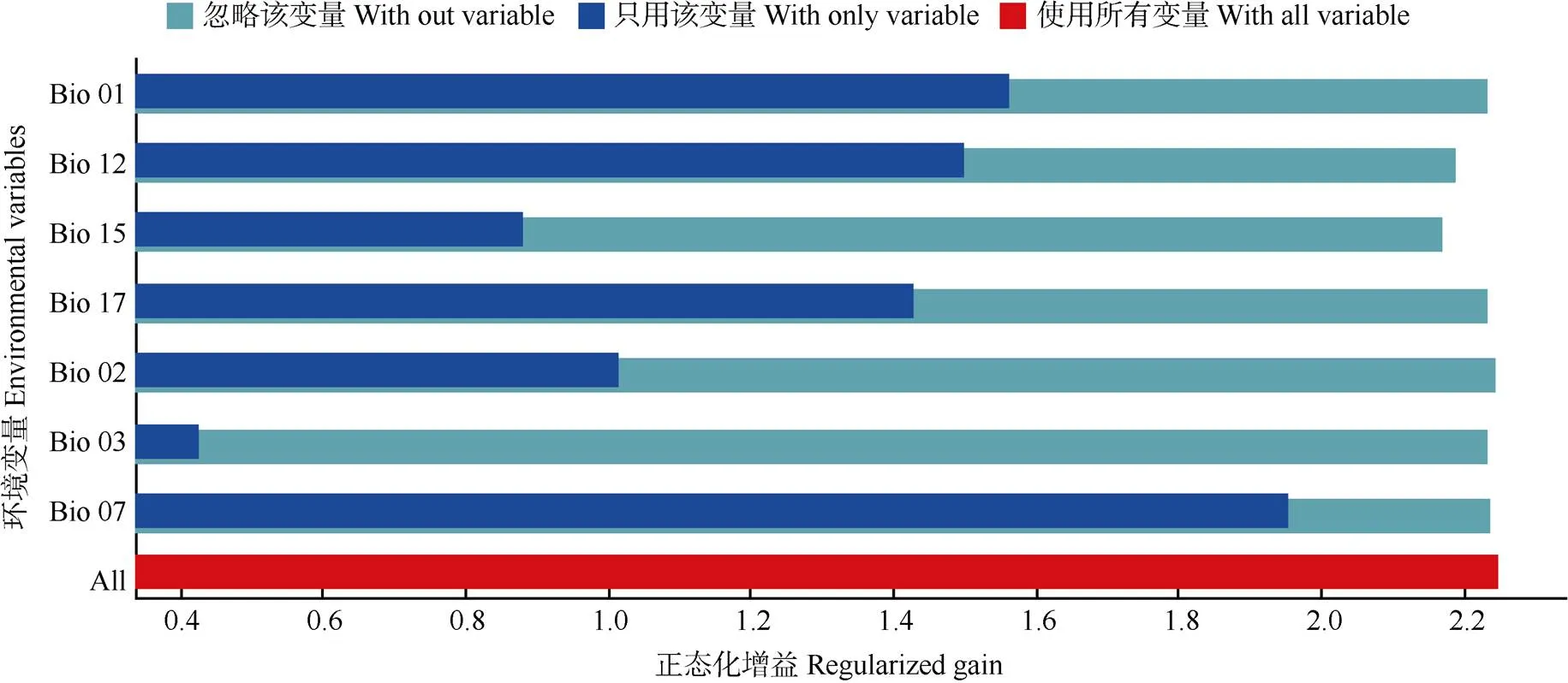
圖2 刀切法檢驗(yàn)的環(huán)境變量對(duì)兜蘭屬植物分布的貢獻(xiàn)率。環(huán)境變量代碼見(jiàn)表1。下同
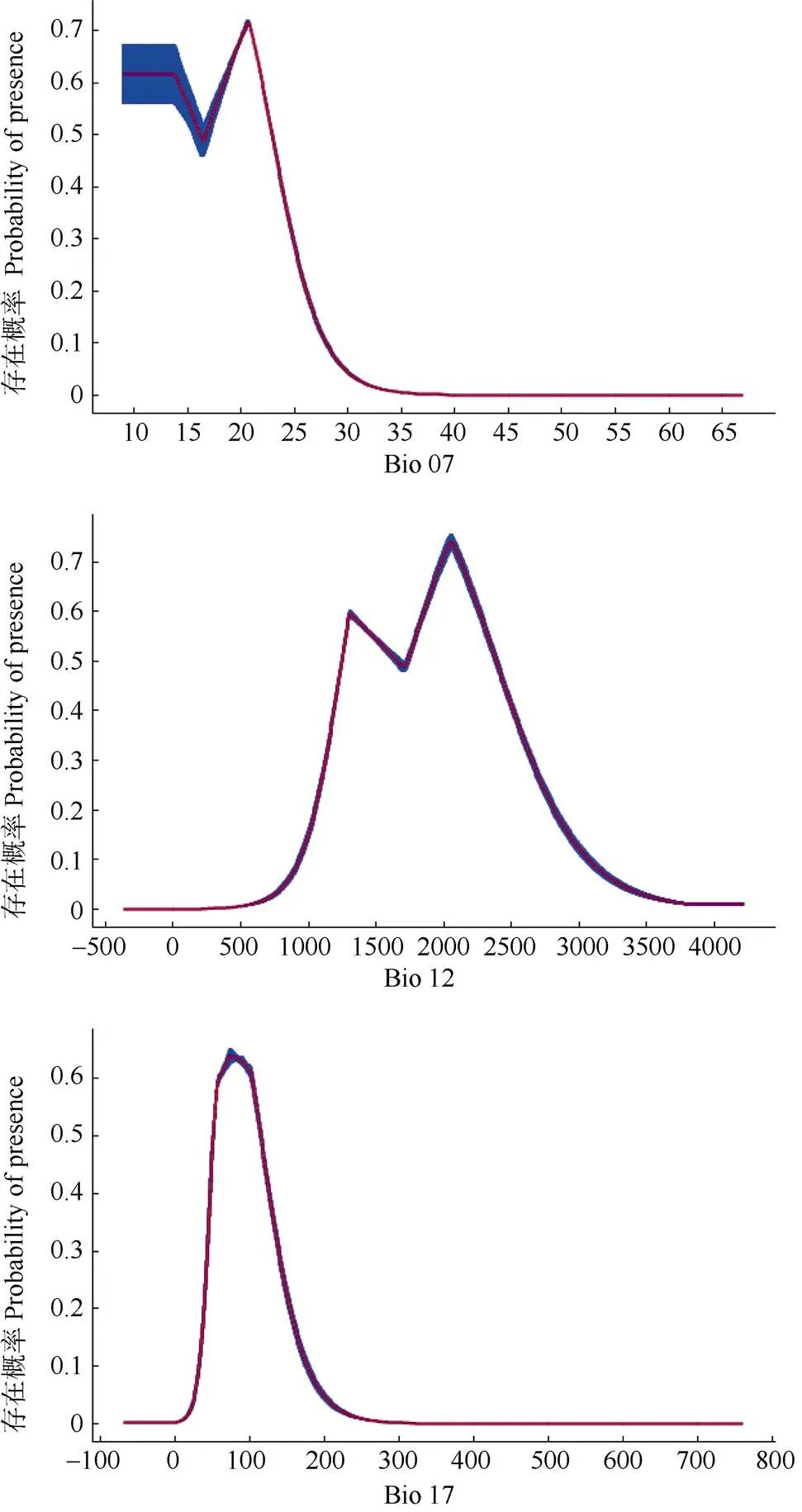
圖3 影響兜蘭屬潛在分布的主導(dǎo)因子響應(yīng)曲線。: 平均值;: 標(biāo)準(zhǔn)誤差。
2.3 當(dāng)代及未來(lái)情景下兜蘭屬植物的潛在地理分布格局
選取當(dāng)代氣候情景和2081—2100年2條共享社會(huì)經(jīng)濟(jì)途徑(SSP1-2.6和SSP5-8.5),利用MaxEnt模型對(duì)兜蘭屬植物和屬下14個(gè)種的潛在地理分布格局進(jìn)行模擬,得到不同情景下的適生區(qū)面積(圖5、6)。
當(dāng)代氣候條件下,兜蘭屬植物的高適生區(qū)面積為27.40×104km2,中適生區(qū)面積為32.44×104km2。在SSP1-2.6情景下,兜蘭屬植物高、中適生區(qū)面積和占比均有擴(kuò)大,面積增大幅度為4.3%和31.9%,達(dá)到28.58×104和42.80×104km2。在SSP5-8.5情景下,高適生區(qū)大幅縮減51%,中適生區(qū)擴(kuò)張16.7%,總適生區(qū)的范圍比當(dāng)代擴(kuò)張了15.8%。
到21世紀(jì)末,在SSP1-2.6情景下,分布在較北地區(qū)的巨瓣兜蘭、小葉兜蘭、同色兜蘭、帶葉兜蘭、長(zhǎng)瓣兜蘭、硬葉兜蘭、秀麗兜蘭、紫毛兜蘭和華南地區(qū)的紫紋兜蘭、卷萼兜蘭的適生區(qū)擴(kuò)張,而杏黃兜蘭、格力兜蘭的適生區(qū)收縮。在SSP5-8.5情景下,杏黃兜蘭、帶葉兜蘭、巨瓣兜蘭、同色兜蘭、長(zhǎng)瓣兜蘭、格力兜蘭、紫紋兜蘭、紫毛兜蘭的適生區(qū)擴(kuò)張,而卷萼兜蘭、小葉兜蘭、硬葉兜蘭、秀麗兜蘭的適生區(qū)收縮;在2種氣候情景下,巧花兜蘭和麻栗坡兜蘭的適生區(qū)都有不同程度的收縮,在SSP5-8.5情景下收縮幅度更大。
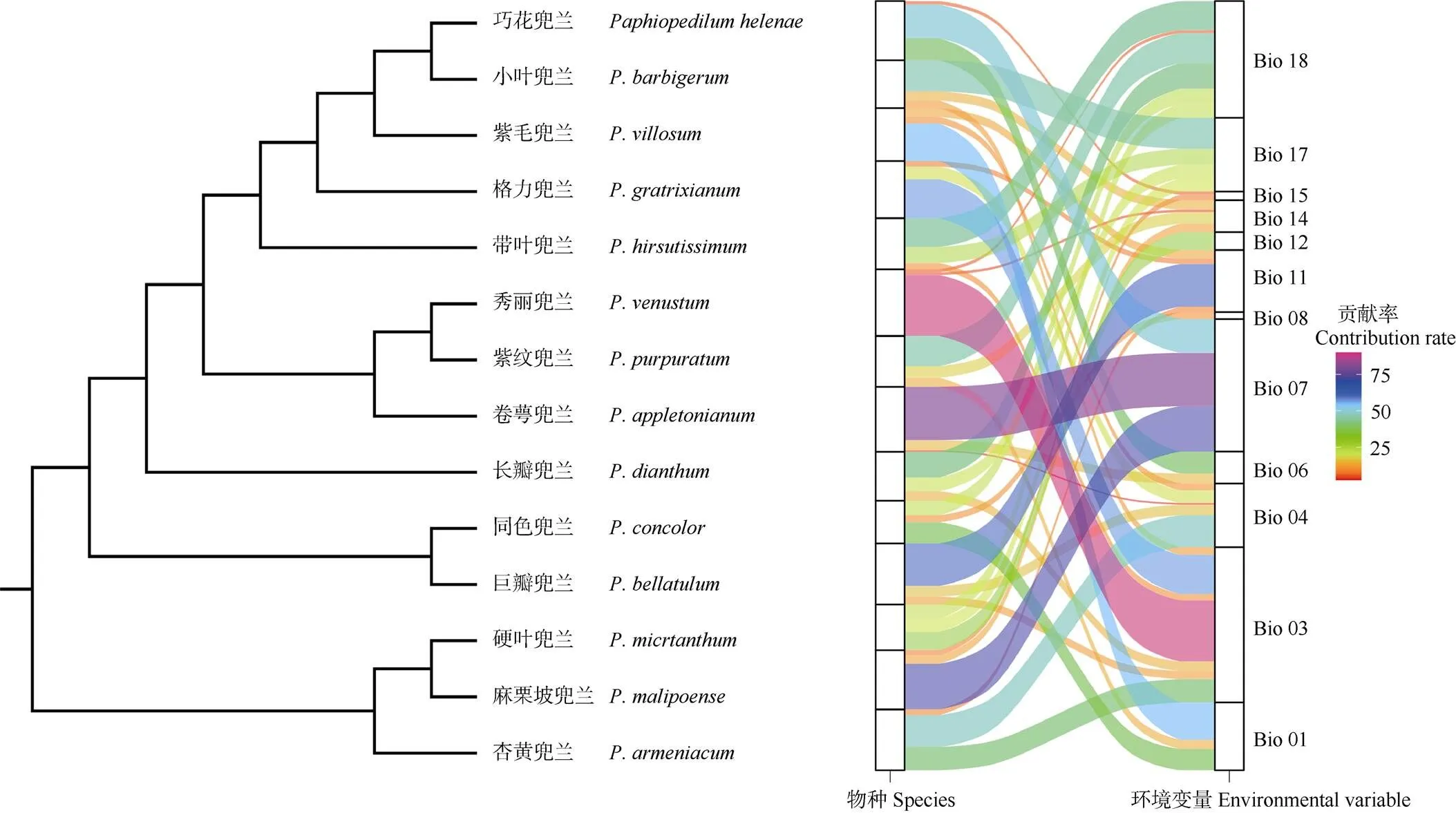
圖4 不同植物類群與環(huán)境變量的關(guān)系
計(jì)算各物種主導(dǎo)氣候因子的變異范圍,發(fā)現(xiàn)在同時(shí)受到多個(gè)環(huán)境因子影響的物種中,紫紋兜蘭適宜生境的氣候因子變異范圍較大,氣候適應(yīng)性較強(qiáng),而長(zhǎng)瓣兜蘭適宜生境的氣候因子變異范圍較為狹窄,對(duì)氣候的要求較苛刻。
當(dāng)代氣候條件下,紫紋兜蘭潛在分布區(qū)與現(xiàn)存記載的分布區(qū)域較為吻合,分布于福建、廣東、廣西南部,以及云南東南部和香港。紫紋兜蘭在中國(guó)的適生總面積為21.04×104km2,高度適生區(qū)面積和中度適生區(qū)面積分別為2.68×104和6.16×104km2。SSP1-2.6情景下,2081—2100年紫紋兜蘭的高、中度適生區(qū)面積都呈增加趨勢(shì),增加幅度為15.4%和19.5%;在SSP5-8.5情景下,2081—2100年紫紋兜蘭高、中適生區(qū)范圍略有增加,幅度為5%和7%。
當(dāng)代氣候條件下,長(zhǎng)瓣兜蘭的高、中適生區(qū)主要分布于云南東南部、廣西西部、貴州南部,零星分布于四川南部、重慶西南部和廣東西南部,與現(xiàn)有的野外種群記載較為吻合。長(zhǎng)瓣兜蘭在中國(guó)的適生總面積為25.87×104km2,高度適生區(qū)面積和中度適生區(qū)面積分別為3.91×104和5.40×104km2。在SSP1-2.6情景下(2081—2100年),高度適生區(qū)和中度適生區(qū)面積分別擴(kuò)大23%和6%;而在SSP5-8.5情景下(2081—2100年),高度適生區(qū)面積縮小43%,中度適生區(qū)面積擴(kuò)大18.5%。
3 結(jié)論和討論
3.1 兜蘭屬植物的分布格局
本研究利用最大熵模型模擬了兜蘭屬14種植物在1970—2000年和2種氣候情景下2081—2100年的分布區(qū),根據(jù)模型的ROC曲線和AUC值, 本次模擬的結(jié)果是理想、可靠的。影響兜蘭屬的主要環(huán)境因子是年降水量、年溫度變化范圍和最干旱季度降水量,降水是影響兜蘭屬物種分布的最主要因素。
前人[27]研究表明,中國(guó)的南亞熱帶地區(qū)是兜蘭屬的生態(tài)多樣性中心。本研究結(jié)果也表明在當(dāng)代氣候條件下,兜蘭屬的最適宜分布區(qū)位于滇東南地區(qū)、貴州西南、南部地區(qū)、廣西西部、廣東南部和海南北部。結(jié)合中心野外實(shí)地調(diào)查與標(biāo)本、文獻(xiàn)的數(shù)據(jù),在全國(guó)范圍內(nèi)本預(yù)測(cè)結(jié)果與兜蘭屬的實(shí)際野外分布區(qū)基本相吻合,且高適生區(qū)也符合兜蘭屬的起源和演化中心。
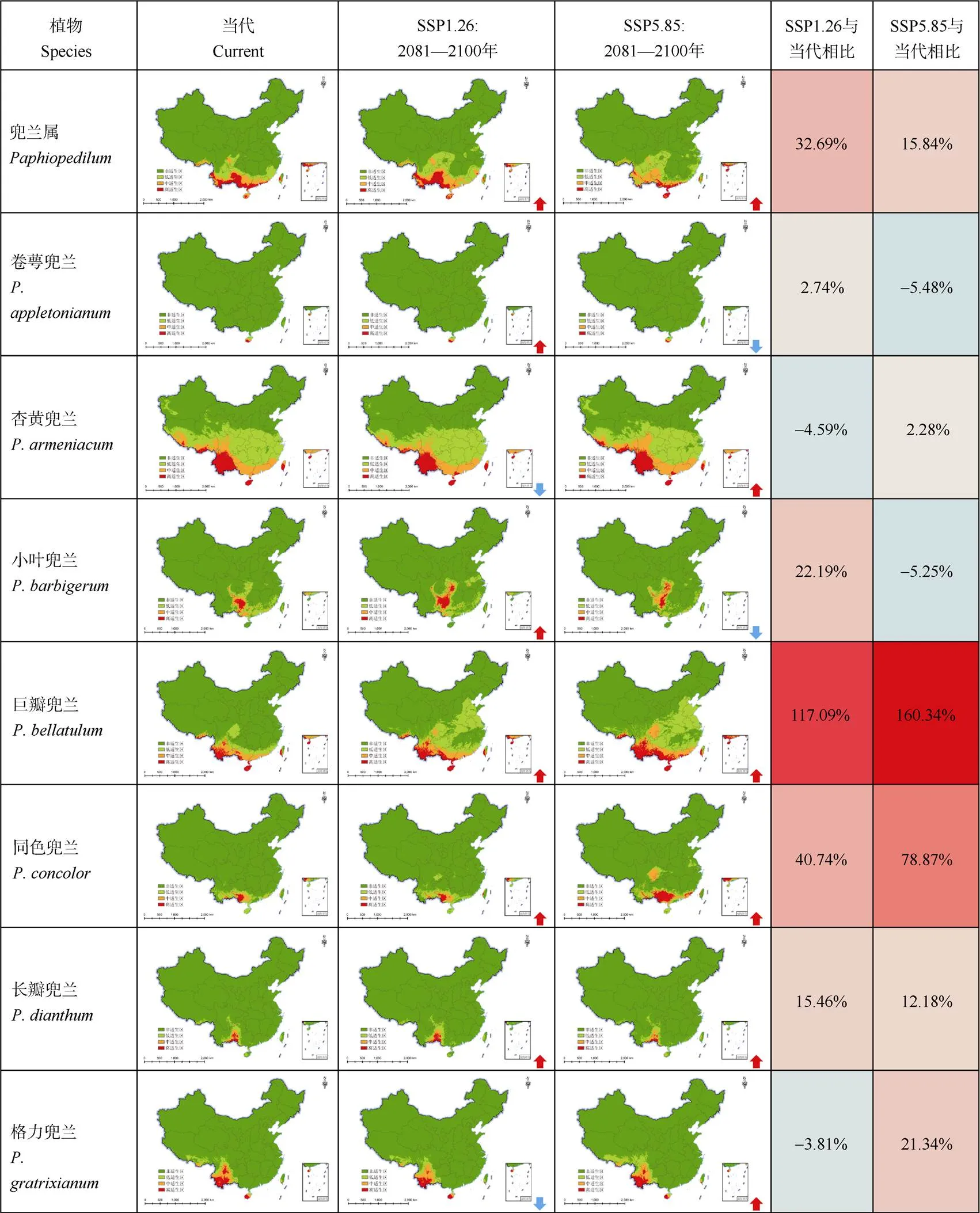
圖5 不同氣候變化情景下兜蘭屬的潛在分布格局及其變化趨勢(shì)。: 擴(kuò)張趨勢(shì);:收縮趨勢(shì);: –6%~160%。下同
我國(guó)亞熱帶地區(qū)主要受東亞季風(fēng)的影響,其中西部地區(qū)主要受印度洋西南季風(fēng)的影響, 而東部地區(qū)主要受太平洋季風(fēng)的影響, 季風(fēng)帶來(lái)的大量降水造就了我國(guó)亞熱帶豐富的生物多樣性。隨著全球氣溫升高,中國(guó)區(qū)域極端降水發(fā)生頻率增強(qiáng),且我國(guó)西南地區(qū)降水強(qiáng)度增加更為顯著[28]。與當(dāng)代氣候相比,在2種不同的未來(lái)氣候情景下兜蘭屬植物的適生區(qū)均呈擴(kuò)張趨勢(shì),雖然整體適生區(qū)的破碎化區(qū)域鏈接成帶狀,但是若僅針對(duì)高適生區(qū)而言破碎化愈加嚴(yán)重。隨著全球變暖,兜蘭屬植物的生境適生區(qū)有向北和西北方向擴(kuò)張的趨勢(shì),逐漸往西北亞熱帶方向延伸,這可能與氣候變暖條件下亞熱帶北界向北推移有很大的關(guān)聯(lián)[29]。在SSP5-8.5的情景下, 整體適生區(qū)范圍雖是呈擴(kuò)張趨勢(shì),但高適生區(qū)出現(xiàn)了大幅度的收縮,這表明隨著全球溫度的上升和降水的增加,部分不適生區(qū)將變成可適生區(qū), 然而溫度的劇烈上升和降水量的大幅度增加,也將導(dǎo)致物種的部分原生境不再適合其生存。
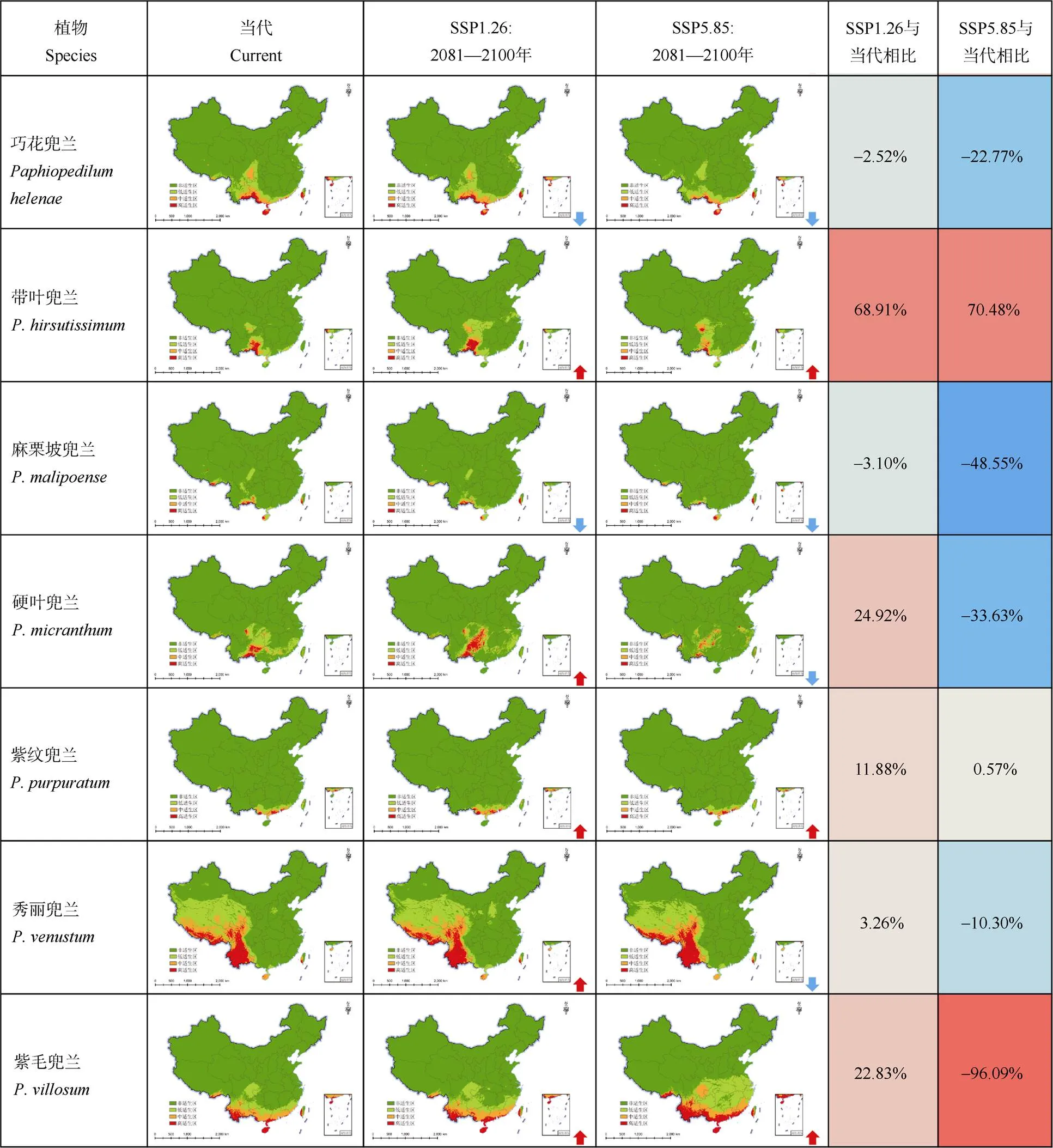
圖6 不同氣候變化情景下兜蘭屬的潛在分布格局及其變化趨勢(shì)
3.2 兜蘭屬植物的保護(hù)策略
雖然兜蘭屬物種有地生、石上附生和樹(shù)上附生之分,但是生于地面的兜蘭其生境要求與附生兜蘭也大致相近[14],因此在對(duì)兜蘭屬物種進(jìn)行保護(hù)時(shí), 首先要保證其附生的環(huán)境。在未來(lái)氣候情景下,不同種群的分布區(qū)變化規(guī)律并不一致,其分布格局響應(yīng)氣候變化的趨勢(shì)也有所不同,因此我們建議對(duì)不同變化趨勢(shì)的物種采用不同的保護(hù)策略。但首先要做的應(yīng)該是在物種的高適生區(qū)加強(qiáng)力度開(kāi)展資源調(diào)查,尋找可能存在的新種群的信息,為后續(xù)開(kāi)展工作提供更準(zhǔn)確的種群數(shù)據(jù)。例如紫紋兜蘭等對(duì)氣候的適應(yīng)性較強(qiáng)的,未來(lái)能適應(yīng)溫度、降水變化幅度較大的環(huán)境的物種,表明其具有遷地保護(hù)和就地保護(hù)的生物學(xué)基礎(chǔ)。在原產(chǎn)地因人為因素造成種群大幅縮減的當(dāng)前,應(yīng)盡快將一些植株遷往異地予以保存,并開(kāi)展人工繁育,在其未來(lái)新增或原有的高適生區(qū)域,開(kāi)展物種的遷地、就地保護(hù)或者野外回歸工作,穩(wěn)定種群數(shù)量和擴(kuò)大種群規(guī)模。又比如長(zhǎng)瓣兜蘭這類雖對(duì)氣候要求較為苛刻,但是在未來(lái)情境下分布區(qū)仍然呈擴(kuò)張趨勢(shì)的,首要是開(kāi)展種質(zhì)資源的收集工作,再者,從保護(hù)其原有生境入手,減少人為活動(dòng)造成的損害,確保原生境為物種提供維持其生存的環(huán)境,開(kāi)展就地保護(hù)工作和加強(qiáng)動(dòng)態(tài)監(jiān)測(cè)的力度。在未來(lái)氣候變化而新增的高適生區(qū),應(yīng)在衡量該區(qū)域物種多樣性和生態(tài)位競(jìng)爭(zhēng)的前提下, 嘗試人為主動(dòng)地、有管理地開(kāi)展物種輔助遷移[30], 從而使其種群未來(lái)的存續(xù)性更高。
而對(duì)于未來(lái)情景下縮減明顯的物種,例如巧花兜蘭,其高適生區(qū)穩(wěn)定存在的地區(qū)可能會(huì)成為將來(lái)應(yīng)對(duì)氣候變化的避難所。在過(guò)去氣候變化的過(guò)程中,避難所為生物體提供了在不利條件下維持其生存和生物多樣性的環(huán)境, 在應(yīng)對(duì)未來(lái)全球氣候變暖可能會(huì)導(dǎo)致的極端高溫等不利事件下, 避難所同樣可以為生物提供生存保障, 其保護(hù)值得優(yōu)先考慮[31],應(yīng)特別重視對(duì)這些區(qū)域原生境的保護(hù),確保在不對(duì)避難所的生境造成人為損傷的前提下開(kāi)展小幅度的人為干預(yù),提高物種的生存率。加強(qiáng)和做好對(duì)種群的長(zhǎng)期監(jiān)測(cè)工作,做好種質(zhì)資源的保存工作,防止物種滅絕,在合適的條件下開(kāi)展物種的回歸工作。
針對(duì)以上研究結(jié)果,為了更好的開(kāi)展兜蘭屬物種的保護(hù)工作,應(yīng)根據(jù)模型結(jié)果開(kāi)展實(shí)地調(diào)查,摸清遺漏區(qū)的資源現(xiàn)狀,收集更多物種的種群信息。對(duì)于兜蘭屬植物分布密集地區(qū)(云南東南部、廣西西部),應(yīng)當(dāng)擴(kuò)大當(dāng)?shù)貒?guó)家級(jí)自然保護(hù)區(qū)的面積,減少保護(hù)空缺區(qū)域,或?qū)⑹〖?jí)、縣級(jí)自然保護(hù)區(qū)升級(jí)為國(guó)家級(jí)保護(hù)區(qū)。從現(xiàn)有保護(hù)區(qū)的整體分布來(lái)看, 我國(guó)南方的保護(hù)區(qū)總體上呈現(xiàn)片斷化不連續(xù)格局, 并未建立較為連續(xù)的自然保護(hù)區(qū), 今后應(yīng)在多樣性高的區(qū)域優(yōu)先進(jìn)行自然保護(hù)區(qū)網(wǎng)絡(luò)建設(shè), 提高相鄰自然保護(hù)區(qū)的連通性、減少人為經(jīng)濟(jì)活動(dòng)對(duì)其生存構(gòu)成的威脅[32],維持生境的穩(wěn)定和連續(xù)。對(duì)物種開(kāi)展野外回歸、瀕危小種群復(fù)壯、生境優(yōu)化等保護(hù)工作后還需開(kāi)展長(zhǎng)期的管理和野外監(jiān)測(cè), 以確保兜蘭屬種群的自然繁衍、適應(yīng)性進(jìn)化和遺傳多樣性。
[1] ZHANG W J, CHEN J K. Advances in study of the distribution area of species [J]. Biodiv Sci, 2003, 11(5): 364–369. [張文駒, 陳家寬. 物種分布區(qū)研究進(jìn)展[J]. 生物多樣性, 2003, 11(5): 364–369. doi: 10. 3321/j.issn:1005-0094.2003.05.002.]
[2] VELOZ S D, WILLIAMS J W, BLOIS J L, et al. No-analog climates and shifting realized niches during the late quaternary: Implications for 21st-century predictions by species distribution models [J]. Glob Change Biol, 2012, 18(5): 1698–1713. doi: 10.1111/j.1365-2486.2011. 02635.x.
[3] BELLARD C, BERTELSMEIER C, LEADLEY P, et al. Impacts of climate change on the future of biodiversity [J]. Ecol Lett, 2012, 15(4): 365–377. doi: 10.1111/j.1461-0248.2011.01736.x.
[4] THOMAS C D, CAMERON A, GREEN R E, et al. Extinction risk from climate change [J]. Nature, 2004, 427(6970): 145–148. doi: 10. 1038/nature02121.
[5] ROOT T L, PRICE J T, HALL K R, et al. Fingerprints of global warming on wild animals and plants [J]. Nature, 2003, 421(6918): 57– 60. doi: 10.1038/nature01333.
[6] IPCC. Climate Change 2013: The Physical Science Basis [M]. Cam- bridge: Cambridge University Press, 2013: 1054–1062.
[7] WANG S X, WU Z F, GONG Y F, et alClimate warming shifts the time interval between flowering and leaf unfolding depending on the warming period [J]. Sci China Life Sci, 2022, 65(11): 2316–2324. doi: 10.1007/s11427-022-2094-6.
[8] ELITH J, LEATHWICK J R. Species distribution models: Ecological explanation and prediction across space and time [J]. Ann Rev Ecol Evol Syst, 2009, 40: 677–697. doi: 10.1146/annurev.ecolsys.110308. 120159.
[9] PHILLIPS S J, ANDERSON R P, SCHAPIRE R E. Maximum entropy modeling of species geographic distributions [J]. Ecol Model, 2006, 190(3/4): 231–259. doi: 10.1016/j.ecolmodel.2005.03.026.
[10] XU Z L, PENG H H, PENG S Z. The development and evaluation of species distribution models [J]. Acta Ecol Sin, 2015, 35(2): 557–567. [許仲林, 彭煥華, 彭守璋. 物種分布模型的發(fā)展及評(píng)價(jià)方法[J]. 生態(tài)學(xué)報(bào), 2015, 35(2): 557–567. doi: 10.5846/stxb201304030600.]
[11] ZHU Y Y, XU X T. Effects of climate change on the distribution of wild population of, an endangered and endemic species in China [J]. Chin J Ecol, 2019, 38(6): 1629–1636. [朱瑩瑩, 徐曉婷. 氣候變化對(duì)我國(guó)特有瀕危物種水杉野生種群分布的影響[J]. 生態(tài)學(xué)雜志, 2019, 38(6): 1629–1636. doi: 10.13292/j.1000- 4890.201906.018.]
[12] ZHANG J Q, XU Z P, WAN T, et al. Study on potential distribution areas ofin China under future climatic conditions [J]. J Plant Resour Environ, 2019, 28(3): 51–57. [張佳琦, 徐振朋, 宛濤, 等. 未來(lái)氣候條件下裸果木在中國(guó)的潛在分布區(qū)研究[J]. 植物資源與環(huán)境學(xué)報(bào), 2019, 28(3): 51–57. doi: 10.3969/j.issn. 1674-7895.2019.03.07.]
[13] HU X G, JIN Y Q, WANG X R, et al. Predicting impacts of future climate change on the distribution of the widespread conifer[J]. PLoS One, 2015, 10(7): e0132326. doi: 10.1371/ journal.pone.0132326.
[14] LIU Z J, CHEN X Q, CHEN L J, et al. The Genusin China [M]. Beijing: Science Press, 2009: 8–10. [劉仲健, 陳心啟, 陳利君, 等. 中國(guó)兜蘭屬植物 [M]. 北京: 科學(xué)出版社, 2009: 8–10.]
[15] LUO Y B, JIA J S, WANG C L. A general review of the conservation status of Chinese orchids [J]. Biodiv Sci, 2003, 11(1): 70–77. [羅毅波, 賈建生, 王春玲. 中國(guó)蘭科植物保育的現(xiàn)狀和展望[J]. 生物多樣性, 2003, 11(1): 70-77. doi: 10.3321/j.issn:1005-0094.2003.01.010.]
[16] LUO Y B, JIA J S, WANG C L. Conservation strategy and potential advantages of the Chinese[J]. Biodiv Sci, 2003, 11(6): 491–498. [羅毅波, 賈建生, 王春玲. 初論中國(guó)兜蘭屬植物的保護(hù)策略及其潛在資源優(yōu)勢(shì)[J]. 生物多樣性, 2003, 11(6): 491-498. doi: 10. 3321/j.issn:1005-0094.2003.06.009.]
[17] WANG Z, CONG L, LIU Y. A review ofresearch [J]. Sci Silv Sin, 2006, 42(7): 113–119. [王貞, 叢磊, 劉燕. 兜蘭屬植物研究現(xiàn)狀[J]. 林業(yè)科學(xué), 2006, 42(7): 113–119. doi: 10.3321/j.issn: 1001-7488.2006.07.020.]
[18] Chen S C. Orchids and their conservation in China [C]// Proceedings of the 5thAsia Pacific Orch Conference & Show. 1996: 49–52.
[19] Delectis Florae Reipublicae Popularis Sinicae, Agendae Academiae Sinicae Editta. Florae Reipublicae Popularis Sinicae, Tomus 17 [M]. Beijing: Science Press, 2006: 52–72. [中國(guó)科學(xué)院中國(guó)植物志編輯委員會(huì). 中國(guó)植物志, 第17卷 [M]. 北京: 科學(xué)出版社, 2006: 52– 72.]
[20] Biodiversity Committee of Chinese Academy of Sciences. Catalogue of Life China: 2022 [OL]. (2022-05-26) http://www.sp2000.org.cn/.
[21] ZHOU T J, CHEN Z M, CHEN X L, et al. Interpreting IPCC AR6: Future global climate based on projection under scenarios and on near- term information [J]. Climate Change Res, 2021, 17(6): 652-663. [周天軍, 陳梓明, 陳曉龍, 等. IPCC AR6報(bào)告解讀: 未來(lái)的全球氣候——基于情景的預(yù)估和近期信息[J].氣候變化研究進(jìn)展, 2021, 17(6): 652–663. doi: 10.12006/j.issn.1673-1719.2021.239.]
[22] GUO J, LIU X P, ZHANG Q, et al. Prediction for the potential distri- bution area ofat global scale based on Maxent model [J]. Chin J Appl Ecol, 2017, 28(3): 992–1000. [郭杰, 劉小平, 張琴, 等. 基于Maxent模型的黨參全球潛在分布區(qū)預(yù)測(cè)[J]. 應(yīng)用生態(tài)學(xué)報(bào), 2017, 28(3): 992–1000. doi: 10.13287/j.1001-9332.201703.026.]
[23] DENG F, LI X B, WANG H, et al. The suitability of geographic distribution and the dominant factors of alfalfa based on MaxEnt model in Xilin Gol [J]. Pratac Sci, 2014, 31(10): 1840–1847. [鄧飛, 李曉兵, 王宏, 等. 基于Maxent模型評(píng)價(jià)紫花苜蓿在錫林郭勒盟的分布適宜性及主導(dǎo)因子[J]. 草業(yè)科學(xué), 2014, 31(10): 1840–1847. doi: 10. 11829/j.issn.1001-0629.2013-0686.]
[24] ZHOU Y, YI Y J, YANG Y F, et al. Predicting geographical distributions of homonoia riparia lour by using maximum entropy [J]. Water Resour Hydropower Technol, 2019, 50(5): 73–81. [周揚(yáng), 易雨君, 楊雨風(fēng), 等. 基于最大熵模型預(yù)測(cè)水楊柳的潛在生境分布[J]. 水利水電技術(shù),2019, 50(5): 73–81. doi: 10.13928/j.cnki.wrahe.2019. 05.009.]
[25] ZHANG X Q. Geographical distribution and climatic suitability of typical eco-economical tree species in the dryland of northwest China [D]. Beijing: University of Chinese Academy of Sciences, 2018. [張曉芹. 西北旱區(qū)典型生態(tài)經(jīng)濟(jì)樹(shù)種地理分布與氣候適宜性研究[D]. 北京: 中國(guó)科學(xué)院大學(xué), 2018.]
[26] Merow C, Smith M J, Silander J A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter [J]. Ecography, 2013, 36(10): 1058–1069. doi: 10.1111/j.1600-0587.2013.07872.x.
[27] WANG Y Q. The geography of Chinese species of[J]. Guihaia, 2000, 20(4): 289–294. [王英強(qiáng). 中國(guó)兜蘭屬植物生態(tài)地理分布[J]. 廣西植物, 2000, 20(4): 289–294. doi: 10.3969/j.issn.1000- 3142.2000.04.001.]
[28] CHEN X C, XU Y, YAO Y. Changes in climate extremes over China in a 2 ℃, 3 ℃, and 4 ℃ warmer world [J]. Chin J Atmos Sci, 2015, 39 (6): 1123–1135. [陳曉晨, 徐影, 姚遙. 不同升溫閾值下中國(guó)地區(qū)極端氣候事件變化預(yù)估[J]. 大氣科學(xué), 2015, 39(6): 1123–1135. doi: 10.3878/j.issn.1006-9895.1502.14224.]
[29] MIAO Q L, DING Y Y, WANG Y. Impact of climate warming on the northern boundary of subtropical zone of China [J]. Geogr Res, 2009, 28(3): 634–642. [繆啟龍, 丁園圓, 王勇. 氣候變暖對(duì)中國(guó)亞熱帶北界位置的影響[J]. 地理研究, 2009, 28(3): 634–642. doi: 10.3321/ j.issn:1000-0585.2009.03.008.]
[30] Ste-Marie C, Nelson E A, Dabros A, et al. Assisted migration: Introduction to a multifaceted concept [J]. For Chron, 2011, 87(6): 724–730. doi: 10.5558/tfc2011-089.
[31] NOSS R F. Beyond kyoto: Forest management in a time of rapid climate change [J]. Conserv Biol, 2001, 15(3): 578–590. doi: 10.1046/j. 1523-1739.2001.015003578.x.
[32] Tarabon S, Dutoit T, Isselin-Nondedeu F. Pooling biodi- versity offsets to improve habitat connectivity and species conservation [J]. J Environ Manag, 2021, 277: 111425. doi: 10.1016/j.jenvman.2020. 111425.
Simulation of Potential Distribution ofin China Based on MaxEnt Model
HUANG Zhicong1, SHU Jiangping1,2, YAN Yuehong1, CHEN Jianbing1*
(1. Orchid Conservation and Research Centre of Shenzhen, Shenzhen 518114, Guangdong, China; 2. South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China)
Based on the known distribution and 20 environmental factors, the potential distribution pattern of genuswas simulated by using MaxEnt model under two different sharing economy scenarios (SSP1-2.6, SSP5-8.5) in the present (1970—2000) and the future (2081—2100). The contribution rate of climate factors was analyzed to find out the dominant climate factors affecting species distribution and their variation range. The results showed that the most suitable distribution areas of the genus were located in southeast Yunnan, southwest Guizhou, western Guangxi, southern Guangdong and northern Hainan. The main environmental factors affecting the distribution of the genus were annual precipitation, annual temperature range and precipitation in the driest season. With global warming, the suitable area had a tendency to expand northward and northwestward, gradually extending to the northwest subtropical direction. Under the SSP5-8.5 scenario, the high suitability area showed a large shrinkage. In the future climate scenarios, the distribution patterns of different populations are not consistent, and their distribution patterns respond to climate change trends. Therefore, different conservation strategies are proposed for species with different distribution trends.
; Maximum entropy model; Dominant climate factor; Climate change
10.11926/jtsb.4703
2022-07-15
2022-09-21
中央林業(yè)改革發(fā)展基金項(xiàng)目(粵財(cái)資環(huán)[2019]5號(hào))資助
This work was supported by the National Forestry Reform and Development Fund (Grant No. [2019]5).
黃智聰(1987年生),男,碩士,助理工程師,研究方向?yàn)樘m科植物保護(hù)。E-mail: 342972251@qq.com
* 通訊作者 Corresponding author. E-mail: cjb@cnocc.cn
猜你喜歡
課堂內(nèi)外·初中版(科學(xué)少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2025年2期)2025-02-28 00:00:00
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03

- 熱帶亞熱帶植物學(xué)報(bào)的其它文章
- 基于黃花菜產(chǎn)量與品質(zhì)特征的優(yōu)異種質(zhì)篩選及評(píng)價(jià)
- 不同生長(zhǎng)光強(qiáng)下8種紅樹(shù)植物幼苗葉片的光響應(yīng)特征
- 基于SSR標(biāo)記的紫花風(fēng)鈴木群體遺傳多樣性分析
- 不同培育方式對(duì)橡膠樹(shù)實(shí)生苗生理指標(biāo)、養(yǎng)分及長(zhǎng)勢(shì)的影響
- 低溫促進(jìn)毛竹葉片類黃酮合成及相關(guān)基因的表達(dá)模式分析
- 小桐子植物特異性酪蛋白激酶PS-CK1-5基因的克隆及原核表達(dá)分析