槐提取物和維生素C 組合物對紫外照射誘導的HaCaT 細胞光損傷的保護作用
2024-02-18 13:21:20鄭嘉妮余丹陽張艷美
食品工業科技 2024年1期
孫 璇,李 萍,孫 靜,鄭嘉妮,余丹陽,張艷美,陳 朋,
(1.仙樂健康科技股份有限公司,廣東汕頭 515000;2.汕頭大學醫學院藥理教研室,廣東汕頭 515041)
隨著生活水平的提升,人們對皮膚健康和管理的關注度與日俱增。紫外線輻射作為主要的外源性老化因素對皮膚的傷害長期存在,與人體皮膚的生理病理狀態關系最為密切。維生素C(Vitamin C,VC),又稱為抗壞血酸,是人體健康必需的微量營養素,天然VC主要來源于柑橘類水果和蔬菜[1],有提高免疫力、預防細菌和病毒感染等作用。作為人體內最重要的水溶性維生素之一,VC具有抗氧化、減輕炎性反應、光保護、抗衰老和抗色素沉著的作用,在皮膚健康方向也起著重要作用[2]。槐(Sophora japonicaL.)是一種高大的多年生喬木,因其具有觀賞、藥用和食用價值而受到關注[3]。花/花芽具有外觀美觀、氣味芳香、口感獨特、保健功能優良等優點,在改善氧化應激,調節黑色素沉淀,修復紫外線輻射損傷等方向具有顯著作用[4-6]。近年來的研究表明,槐米中提取的多糖具有保護HaCaT 細胞免受UVB 照射誘導的損傷[5]。槐花中的主要成分蘆丁被認為可以降低順鉑導致的HMCs 細胞的氧化損傷[7]。因此,VC和槐提取物有抗氧化功效,具有對皮膚的光保護作用。
此外,有研究表明蘆丁和VC聯用對超氧陰離子自由基的清除或對脂質過氧化的抑制呈濃度依賴性,且蘆丁與VC組合可以產生超加性效應[8]。蘆丁和VC組合物在紫外輻照的人類皮膚成纖維細胞中也表現出保護作用,二者聯用對促炎信號蛋白的過度表達和DNA 重組/表達有抑制作用[9]。Zhang 等[10]的研究表明槐花和VC組合物顯著提高運動訓練小鼠血清中SOD 活性,且其抗氧化能力較單一物料處理組具有更好的效果。可見,槐提取物與維生素C 組合物有一定的協同抗氧化作用,可能對紫外線照射造成的皮膚光氧化損傷有效。
本實驗室之前的實驗研究表明,槐提取物和VC組合物可以促進紫外照射后皮膚成纖維細胞彈性蛋白的生成,且槐提取物和VC組合物的質量比為1:1 時有協同增效的作用。為進一步探究槐提取物和VC特定比例組合物對人體皮膚細胞的影響,本研究以槐提取物和VC為實驗材料,以HaCaT 細胞為研究對象,通過檢測槐提取物、VC及其組合物對UV 導致的光損傷的抗氧化保護與抗炎的初步探究,為槐提取物和維生素C 組合物的開發和利用提供實驗支撐。
1 材料與方法
1.1 材料與儀器
槐提取物 河南中大恒源生物科技股份有限公司(10%蘆丁,水提、包埋、干燥等工藝制備);維生素C(合成)帝斯曼江山制藥(江蘇)有限公司;胎牛血清(FBS)、0.25%胰蛋白酶/EDTA、青霉素/鏈霉素、DMEM 培養基 Gibco 公司;磷酸鹽緩沖液(PBS)普諾賽生命科技有限公司;6-羧基-2',7'-二氯二氫熒光素二乙酸酯(H2DCF-DA)Thermo Fisher Scientific 公司;人白介素-6(IL-6)Elisa 試劑盒 RD systems 公司;MitoSOX? Red Invitrogen 公司;鼠源Anti-Thymine Dimer 抗體 Abcam 公司;Alexa Fluor 488 標記山羊抗小鼠IgG(H+L)、抗熒光淬滅封片液(含DAPI)Beyotime Biotechnology 公司;即用型正常山羊血清 武漢博士德生物工程有限公司;永生化人角質形成細胞(HaCaT)普諾賽生命科技有限公司。
Axio Vert.A1 倒置熒光顯微鏡、LSM 880 激光共聚焦顯微鏡 ZEISS 公司;Spark?多模式微孔板酶標儀 TECAN 公司;CL-3000L 紫外交聯儀Analytik Jena 公司;Forma 3111 二氧化碳培養箱Thermo Fisher Scientific 公司。
1.2 實驗方法
1.2.1 槐提取物及維生素C 組合物對UV 誘導的HaCaT 細胞ROS 的影響
1.2.1.1 細胞內總ROS 水平的測定(酶標儀檢測)取對數生長期生長到80%的HaCaT 細胞,以1.5×104個/孔接種于黑壁透明板底的96 孔板。5% CO2的37 ℃培養箱中培養24 h。細胞隨機分為對照組、模型組(UVA 組)及槐提取物處理組(槐提取物組)、維生素C 處理組(VC組)、槐提取物和VC組合物處理組(槐提取物+VC組,質量比為1:1)。棄去上清液,10 mmol/L PBS 洗滌1 次,模型組及不同實驗組加入適量PBS 覆蓋細胞,模型組和各實驗組置于10 J/cm2UVA(波長為365 nm)照射[11],UVA 照射結束后30 min,未接受輻照的對照組和UVA 照射的模型組加入100 μL DMEM 基礎培養基繼續培養,實驗組加入100 μL DMEM 基礎培養基配制的125 μg/mL 槐提取物溶液、125 μg/mL VC溶液、總濃度為125 μg/mL 槐提取物和VC組合物溶液,對照組加入DMEM 基礎培養基。實驗組的處理濃度按照實驗室之前的研究得出。所有組別繼續培養30 min。棄去上清,加入50 μL 含5 μmol/L 的H2DCFDA 的DMEM 基礎培養基,37 ℃避光孵育30 min。多模式微孔板酶標儀(激發光波長Ex/發射光波長Em=488/530)測定各孔熒光值,并按照下式計算細胞內ROS 抑制率[12]:
式中:T-受試樣品熒光強度;C-模型組熒光強度;C0-對照組熒光強度。
1.2.1.2 細胞內總ROS 水平的測定(熒光顯微鏡觀察)取對數期HaCaT 細胞,以6×105個/孔接種于35 mm 培養皿,后續操作與上述類似,H2DCF-DA 探針處理后,PBS 洗滌2 次,熒光顯微鏡下拍照[13]。
1.2.2 線粒體ROS(Mitochondrial DNA,mtROS)水平的測定 取對數生長期生長到80%的HaCaT 細胞,以1.5×104個/孔接種于共聚焦培養皿中,培養24 h。細胞隨機分為對照組、模型組及實驗組。模型組和實驗組經10 J/cm2UVA 照射。模型組加入1 mL 基礎培養基,實驗組加入1 mL DMEM 基礎培養基配制的125 μg/mL 槐提取物溶液、125 μg/mL VC溶液、總濃度為125 μg/mL 槐提取物和VC組合物溶液(質量比為槐提取物:VC=1:1)培養箱中孵育30 min。吸去培養基,PBS 緩沖溶液洗3 次;加入500 μL 含MitoSOX? Red 終濃度為10 μmol/L 的DMEM 基礎培養基,37 ℃孵育30 min。30 min 后,吸去上清,用37 ℃預熱PBS 漂洗3 次,加入1 mL PBS。激光共聚焦顯微鏡觀察并拍照[14],Image-J 軟件分析各組的平均熒光強度(Mean fluorescence intensity,MFI),并計算mtROS 抑制率,計算公式如實驗方法1.2.1。
1.2.3 環丁烷嘧啶二聚體(Cyclobutane pyrimidine dimers,CPDs)含量的測定 24 孔板加入細胞爬片,HaCaT 細胞以1.5×105個/孔的密度種于孔板,培養24 h。細胞隨機分為對照組、模型組及實驗組。模型組和實驗組經30 mJ/cm2UVB(波長為302 nm)照射[15-16]。實驗組加入DMEM 基礎培養基配制的125 μg/mL槐提取物溶液、125 μg/mL VC溶 液、125 μg/mL 槐提取物和VC組合物溶液培養箱中孵育30 min。PBS 洗滌3 次。每孔加入1 mL 預冷4%多聚甲醛,室溫固定20 min;吸棄多聚甲醛,PBS 漂洗3 次,5 min/次。加入1 mL 0.1% PBST,室溫放置15 min;PBS 漂洗3 次,5 min/次。每孔加入1 mL 2 mol/L 鹽酸(現配現用),室溫放置10 min;吸棄鹽酸,PBS 漂洗3 次,5 min/次。加入50 μL 山羊血清封閉,覆蓋蓋玻片,室溫封閉1 h。吸棄封閉液,加入5%牛血清蛋白(BSA)稀釋的鼠源CPD 抗體,覆蓋蓋玻片,置于抗體孵育濕盒中,4 ℃過夜。PBS 漂洗3 次,5 min/次。加入5% BSA 稀釋的熒光二抗(Alexa Fluor488 標記的山羊抗鼠IgG),室溫避光孵育1 h;吸棄二抗,PBS 漂洗3 次,5 min/次。用含DAPI 的抗熒光淬滅劑封片。觀察并拍照,Image-J 軟件分析各組的MFI。
1.2.4 細胞因子IL-6 分泌水平的測定 HaCaT 細胞按照5×106濃度接種于10 cm 細胞培養皿中。10 J/cm2UVA 照射結束后DMEM 基礎培養基配制的125 μg/mL 槐提取物溶液、125 μg/mL VC溶 液、125 μg/mL 槐提取物和VC組合物溶液,置于培養箱中繼續培養6 h[17]。按照IL-6 Elisa 檢測試劑盒檢測IL-6 的含量。
1.3 數據處理
實驗數據至少3 次重復,實驗結果用Graphpad 8.0.0 統計軟件分析并繪圖,結果以mean±SD 表示,統計方法為單因素方差分析(One-way,ANOVA)。*P<0.05、**P<0.01、#P<0.05、##P<0.01。
2 結果與分析
2.1 槐提取物和維生素C 組合物對HaCaT 細胞ROS 的影響
具有細胞膜滲透性的ROS 熒光檢測探針H2DCFDA 進入細胞后,被水解生成DCFH 而無法穿過細胞膜,從而被裝載到細胞內,ROS 可將DCFH 氧化成在激發時發出綠色熒光產物DCF,因此DCF 的熒光強度可反映細胞內ROS 水平[18]。如圖1a 所示,對照組幾乎無熒光,UVA 處理后綠色熒光強度明顯增強,說明細胞內ROS 含量明顯增加,UVA 誘導的氧化應激導致細胞內ROS 含量增加。與UVA 組相比,槐提取物、VC及組合物處理組熒光明顯減弱,其中槐提取物和VC組合物組熒光最弱。結果表明,UVA 照射增加細胞內ROS 的含量,槐提取物、VC及組合物均可降低細胞中ROS 含量,且組合物的ROS 清除效果最好。如圖1b 所示,UVA 處理后,槐提取物的ROS 抑制率為38.23%±9.23%;VC的ROS抑制率為27.20%±6.87%;組合物的ROS 抑制率為64.13%±4.08%,槐提取物、VC及其組合物顯著降低UVA 導致的ROS 生成(對照組vs.槐提取物,P<0.01;對照組vs.VC,P<0.01;對照組vs.組合物,P<0.01)。結果表明,UVA 照射后,組合物組ROS 清除率極顯著高于單一組分處理組(組合物vs.槐提取物,P<0.01;組合物vs.VC,P<0.01)。熒光顯微法與酶標儀檢測均顯示槐提取物和VC組合物對于ROS 的清除效果最好。多酚(包括蘆丁)和VC可顯著降低抗氧化蛋白(如硫氧還蛋白和戊二醛)在細胞中的表達,硫氧還蛋白還原酶是一種在氧化應激下恢復細胞中還原硫氧還蛋白水平的酶[9]。因此,槐提取物和VC可支持抗氧化系統。槐花/槐米中含有豐富的蘆丁,蘆丁作為一類黃酮類物質,通過增加抗氧化(如SOD、TrxR 和Prx1/2)和炎癥反應(如IL-17F、PAK2和YWHAZ)相關蛋白總表達的增加發揮保護作用[19]。此外,蘆丁通過恢復照射后增加的p53、細胞色素c、細胞周期和凋亡調節蛋白2 的生理水平,部分防止紫外線誘導的細胞凋亡[19]。VC可以抑制紫外輻照誘導的表皮角質形成細胞中SOD、CAT、GST 和GSH-Px活性的降低,從而維持細胞內抗氧化防御系統[20-21]。一項關于蘆丁和VC聯用對紫外誘導氧化應激的HaCaT 細胞保護作用的研究表明蘆丁與VC組合物導致UVB 照射的HaCaT 細胞中超氧陰離子的生成減少約60%,優于蘆丁和VC的單獨作用[22]。本研究表明,槐提取物和VC對UVA 紫外輻射導致的細胞氧化損傷也具有保護作用,二者單獨使用可減少細胞ROS 及mtROS 的生成,對紫外輻照導致的氧化損傷起到保護作用。此外,本研究將槐提取物與VC進行特定比例配比,探究出一種對于對紫外線輻射導致的細胞氧化損傷具有更強保護作用的復合物。
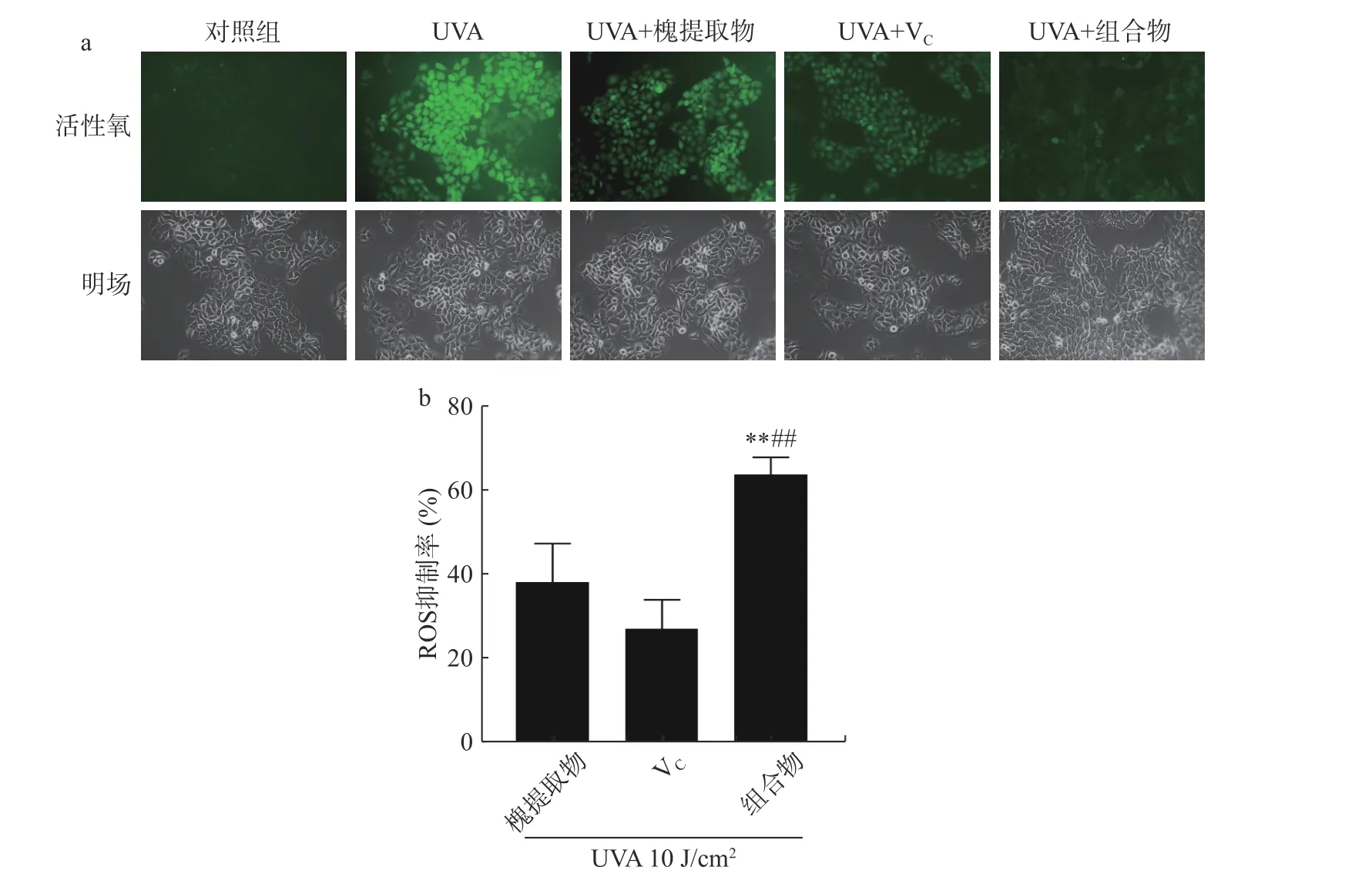
圖1 槐提取物、維生素C 及其組合物對UVA 照射HaCaT 細胞ROS 含量的影響(n=4)Fig.1 Effects of Sophora japonica L.,vitamin C and combination on ROS generation in UVA irradiated HaCaT cells (n=4)
2.2 槐提取物和維生素C 組合物對HaCaT 細胞mtROS 的影響
如圖2 所示,紅色熒光表示線粒體內超氧陰離子含量,對照組幾乎無熒光,UVA 組的熒光強度顯著增強,槐提取物、VC及組合物處理后熒光強度顯著減弱,其中槐提取物和VC組合物處理組的熒光強度弱于單一物料處理組。UVA 照射后,HaCaT 細胞mtROS 含量增加,分別給予槐提取物、VC及其組合物處理后mtROS 抑制率分別為50.29%±4.92%、38.81%±8.66%、84.74%±3.68%,均可一定程度上減少線粒體ROS 的生成,其中槐提取物和VC組合物處理組效果最佳(槐提取物+VCvs.槐提取物,P<0.01;槐提取物+VCvs.VC,P<0.01)。線粒體膜是細胞內ROS 的重要來源[23]。研究表明,VC對線粒體具有多種有益作用,包括直接清除ROS 以及基于分散在基質和膜中的其他線粒體抗氧化劑的再循環的關鍵作用[23]。VC對線粒體的復雜調節可能有助于多功能的抗氧化反應,從而為維生素提供核心作用,以充分控制與線粒體ROS 產生增加相關的線粒體功能障礙[24]。本研究也表明,槐提取物、VC及其組合物可抑制mtROS 的生成,且組合物具有一定的加成效果。
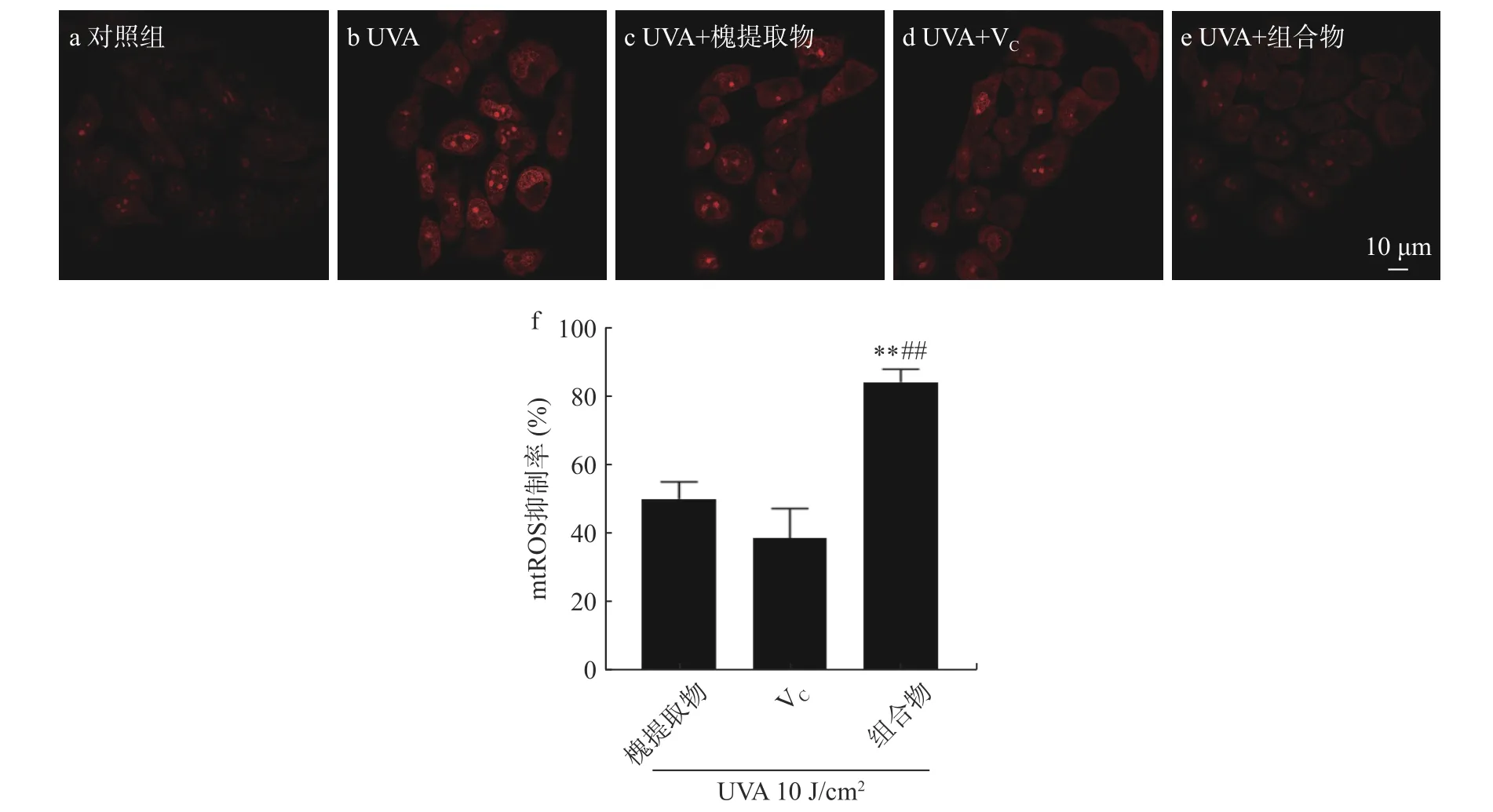
圖2 槐提取物、維生素C 及其組合物對UVA 照射HaCaT 細胞線粒體ROS 含量的影響(n=3)Fig.2 Effects of Sophora japonica L.,vitamin C and combination on mitochondrial ROS generation in UVA-irradiated HaCaT cells (n=3)
2.3 槐提取物和維生素C 組合物對HaCaT 細胞內CPDs 水平的影響
如圖3a 所示,UVB 照射后細胞中熒光強度明顯增加,說明UVB 導致細胞中的CPDs 水平上升。與UVB 照射組相比,槐提取物、VC及其組合物處理后CPDs 的水平顯著降低。對照組、模型組(UVB照射組)、槐提取物組、VC組及組合組的CPDs 的熒光強度分別為:6.95±0.80、23.22±1.23、13.10±1.21、15.75±0.44、9.17±0.47。與對照組比較,UVB 組細胞中CPDs 含量顯著升高(P<0.01),說明UVB 損傷模型造模成功;與模型組比較槐提取物組、VC組和組合組CPDs 含量均極顯著下降(P<0.01);其中,槐提取物和維生素C 組合組效果顯著優于單一組分處理組(P<0.05)。以上結果表明,槐提取物、VC及其組合組均可降低UVB 照射導致的細胞中高水平的CPDs,其中組合物對DNA 損傷的保護作用最強。
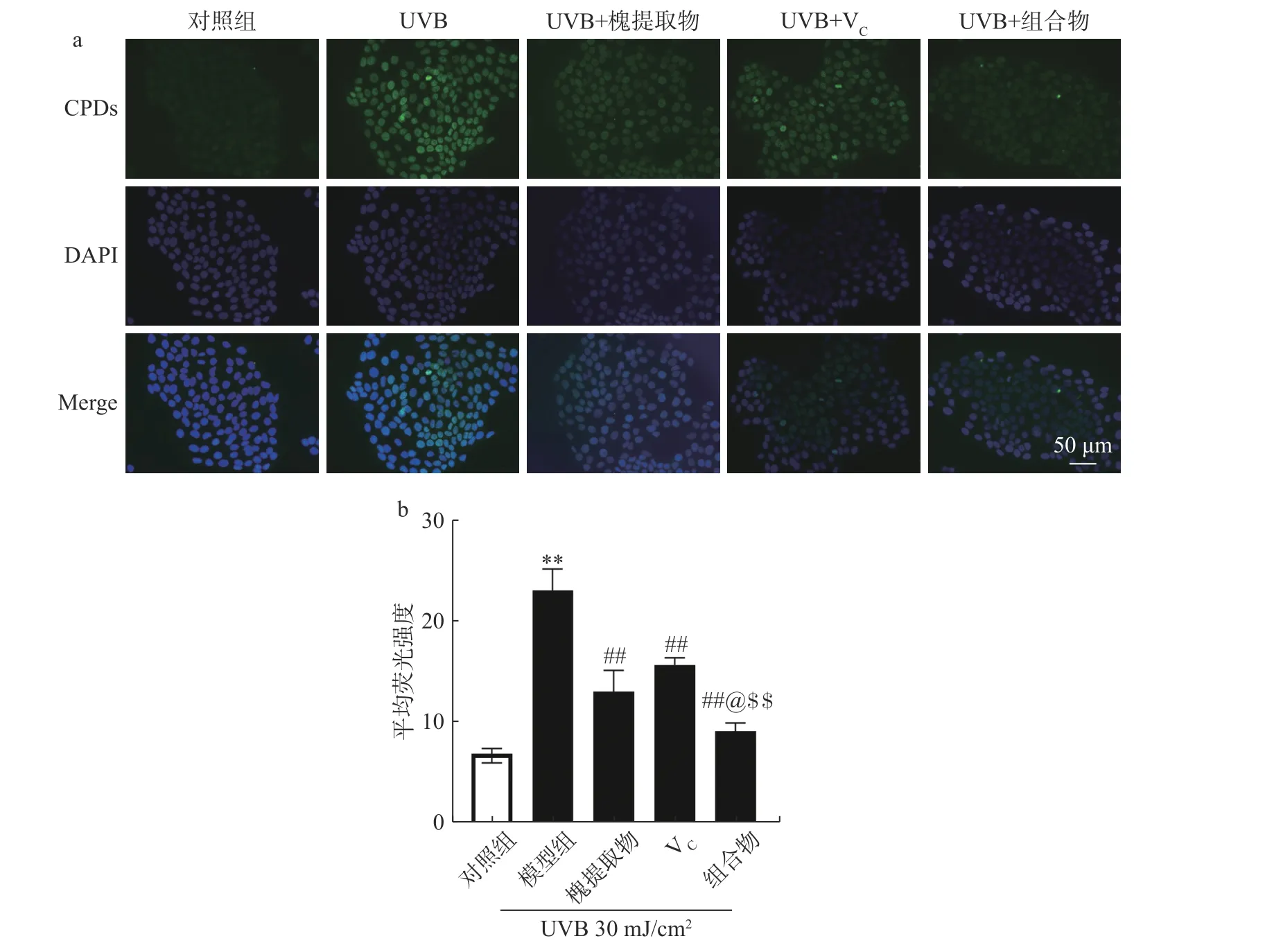
圖3 槐提取物、VC 及組合物對UVB 照射HaCaT 細胞CPDs 含量的影響(n=3)Fig.3 Effects of Sophora japonica L.,vitamin C and combination on CPDs content in UVB-irradiated HaCaT cells (n=3)
皮膚UVR 暴露導致DNA 損傷。CPDs 是UVR誘導DNA 損傷的代表產物,被認為是啟動光致癌的分子觸發器CPDs 的生成主要與UVB 有關,且主要生成于表皮層的細胞中[25-26]。核酸切除修復(NER)是紫外線誘導的DNA 損傷的主要修復途徑,細胞著色性干皮病蛋白A(XPA)在NER 中其中不可或缺的作用[27-28]。研究表明VC可以上調細胞中sirtuin1(SirT1)蛋白的表達,而SirT1 通過與XPA 結合,將XPA 維持在低乙酰化狀態,下調細胞中紫外線誘導的CPDs 的表達,在紫外線的NER 途徑中起到積極作用[29]。蘋果提取物顯著降低導致紫外線照射體外皮膚外植體和3D 組織工程皮膚細胞中CPDs 的形成,這種保護作用可能與蘋果提取物中的蘆丁相關[30]。研究表明蘆丁和VC聯用有增強皮膚細胞的抗UVB 損傷的作用[9]。因此,兩者聯用對UVB 導致的DNA 損傷可能也有一定的加成效果。本研究也發現,UVB 照射則使細胞中的CPDs 含量顯著增加。UVB 照射后給予槐提取物、VC及其組合物,可減少HaCaT 細胞中CPDs 的生成,說明槐提取物、維生素C 及其組合物可促進UVB 誘導的DNA 損傷修復,且組合物具有加成作用。
2.4 槐提取物和維生素C 組合物對HaCaT 細胞內IL-6 水平的影響
UV 作用于HaCaT 細胞,可激活轉錄因子NF-κB,促進多種炎癥因子如IL-1、IL-6、IL-8 等的表達[31]。其中IL-6 是炎癥反應過程中的促發劑,可促進T 細胞增殖與分化,刺激B 細胞分化產生抗體,參與機體的免疫應答[32]。如圖4 所示,與對照組比較,UVA照射組細胞內IL-6 含量極顯著升高(P<0.01);與UVA組比較,槐提取物處理組及組合物處理組細胞內IL-6 含量顯著下降(P<0.05),槐提取物處理組IL-6 水平下降最低(P<0.01),其次組合物處理組(P<0.05)。因此,組合物處理組在UVA 誘導的角質形成氧化應激模型中雖具有抗炎作用,但效果并不優于單用槐提取物處理組抗炎作用。
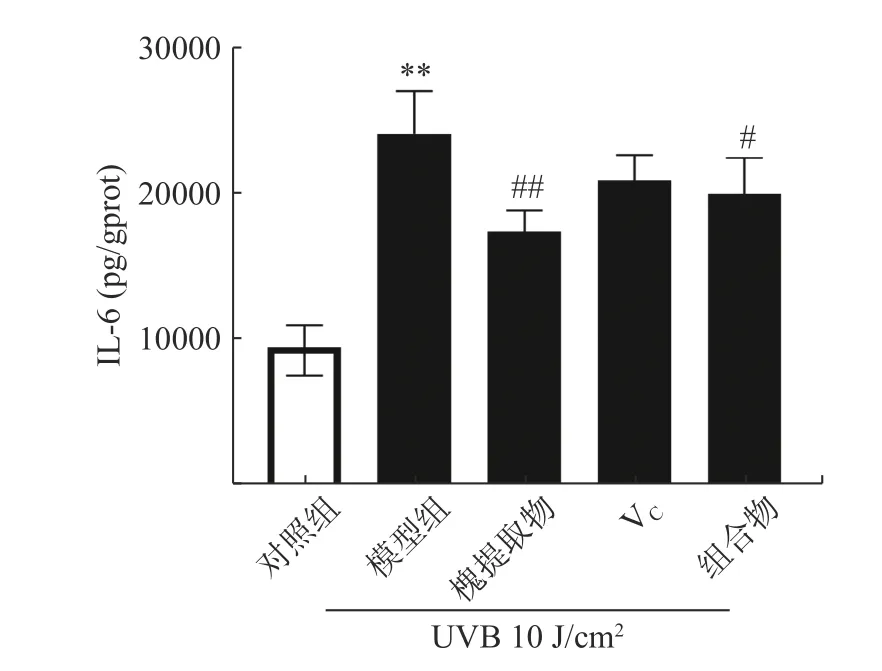
圖4 槐提取物、維生素C 及其組合物對UVA 照射HaCaT 細胞IL-6 水平的影響(n=3)Fig.4 Effects of Sophora japonica L.,vitamin C and combination on IL-6 expression in UVA-irradiated HaCaT cells (n=3)
在生物相關的相互作用機制中,植物提取物聯合使用可通過多靶點效應發揮互補增效的作用,包括介導不同信號通路或同一通路的不同下游環節。本研究發現本研究發現槐提取物和VC均具有抗氧化活性和抗炎作用,VC對抑制ROS、mtROS 的生成方面有優于槐提取物的趨勢,其抗氧化活性更強,而槐提取物的抗炎作用強于VC。氧化應激與炎癥反應間存在的錯綜復雜的調控通路,二者往往相互影響形成正反饋循環[33-34]。本文的研究結果表明,VC和槐提取物組合物抗氧化活性有明顯提高,推測槐提取物和VC分別主要通過拮抗氧化應激和炎癥反應發揮互補增效作用。然而,在抗炎方面本研究表明,VC有抗炎趨勢,槐提取物有較好抗炎效果,VC和槐提取物聯用后雖仍具有較好的抗炎作用,但與槐提取物相比,效果不突出。關于VC和槐提取物聯用后抗氧化效果增強的機制需要進一步進行驗證。
3 結論
本文通過分析槐提取物、VC及其組合物對紫外照射處理的HaCaT 細胞的抗氧化和細胞炎癥因子分泌的影響。結果表明,槐提取物、VC及其組合物具有抑制由UVA 誘導產生的ROS 生成的作用,對細胞ROS 抑制率分別為38.23%±9.23%、27.20%±6.87%及64.13%±4.08%,對mtROS 抑制率分別為50.29%±4.92%、38.81%±8.66%及84.74%±3.68%。組合物對于ROS 的抑制作用優于單一物料處理。槐提取物、VC及其組合物顯著降低由UVB 誘導產生的CPDs 的含量,其中組合物效果最佳。此外,槐提取物及槐提取物和VC組合物顯著減低炎癥因子IL-6 的分泌。該研究可為槐提取物及VC組合物的營養健康產品開發和應用提供實驗支持。而未來的研究將聚焦在槐提取物和VC聯用后抗氧化效果增強的機制進行深入研究與探討。
