大洋硅藻席沉積與冰期旋回中的碳-氮循環(huán)
2024-02-22 14:52:46蔡雯沁黃恩清柳雙權(quán)田軍
海洋地質(zhì)與第四紀地質(zhì) 2024年1期
蔡雯沁,黃恩清,柳雙權(quán),田軍
1. 同濟大學海洋地質(zhì)國家重點實驗室,上海 200092
2. 中國科學院深海科學與工程研究所,三亞 572000
“中生代海洋浮游生物革命”之后,海洋浮游真核藻類出現(xiàn)三大分支,即顆石藻、溝鞭藻和硅藻,它們共同構(gòu)成了現(xiàn)代大洋顆粒有機碳泵以及大洋輸出生產(chǎn)力的主體[1]。其中硅藻輻射分異的最晚,在新生代始新世、漸新世之交,即東南極冰蓋擴張之后,化石記錄中的硅藻多樣性程度才出現(xiàn)第一次的顯著增加[2]。但硅藻演化相當迅速,目前已經(jīng)分異出數(shù)百個屬和大約十萬個種[3]。硅藻發(fā)展出一系列特殊的本領(lǐng)。首先,生活在開放大洋環(huán)境的硅藻,發(fā)育占了細胞40%體積以上的大液泡,這個液泡可以儲存高濃度的硝酸鹽和磷酸鹽[4]。因此,在營養(yǎng)鹽存在頻繁脈沖式供應的地區(qū),例如在近岸上升流或者高緯紊流發(fā)育的海區(qū),硅藻具有明顯的生存競爭優(yōu)勢。一方面,當營養(yǎng)鹽供應突然增加時,硅藻可以高效利用、儲存并快速勃發(fā);另一方面,當營養(yǎng)鹽供應突然消失時,液泡儲存的營養(yǎng)鹽讓硅藻在短時間內(nèi)可以不依賴于外界的供應[2]。其次,由于通過基因水平交換獲得了其他真核生物體的緣故(即二次內(nèi)共生),硅藻存在類似動物的“尿素循環(huán)”[5]。這種功能可以加快細胞內(nèi)碳和氮的周轉(zhuǎn)和運移,促進新陳代謝過程所需的含氮化合物的合成(例如多氨和谷氨酰胺)。因此,當水體環(huán)境出現(xiàn)營養(yǎng)鹽供應時,“尿素循環(huán)”可以幫助硅藻迅速增殖,從長期氮限制環(huán)境中快速恢復和繁盛[5]。
正是硅藻在細胞尺度上的特殊本領(lǐng),使得硅藻在全球規(guī)模的碳、氮循環(huán)中扮演重要角色。據(jù)估算,現(xiàn)今硅藻貢獻了整個大洋約40%的凈生產(chǎn)力和高達50%的有機碳輸出[6]。每年全球硅藻固定二氧化碳的規(guī)模,與整個熱帶雨林相當[3]。硅藻也是著名的大洋“鐵肥實驗”(Iron Fertilization)的研究焦點。在南大洋“高營養(yǎng)鹽低葉綠素海區(qū)”撒入溶解二價鐵之后,會出現(xiàn)小型硅藻的勃發(fā),并且大約一半的勃發(fā)生物量能夠沉降到水深超過1 000 m 的海水中,由此可以推測硅藻勃發(fā)在“碳封存”中的重要角色[7]。
在地質(zhì)時間尺度上,顆石藻和溝鞭藻在白堊紀—古近紀繁盛,而硅藻在漸新世之后崛起。推測在“暖室期”氣候條件下,高緯異常溫暖,經(jīng)向能量差異小,全球風場和洋流較弱,相對靜水環(huán)境有利于顆石藻和溝鞭藻生長。而漸新世之后極地冰蓋體積快速增大,高緯顯著降溫,高緯海區(qū)風場、紊流的發(fā)育有利于硅藻的繁盛[2,8]。但硅藻并非環(huán)境變化的被動產(chǎn)物,硅藻崛起很可能深刻塑造了地球的表層環(huán)境。有學者推測硅藻繁盛導致海洋生物泵效率提高,造成大氣二氧化碳濃度的持續(xù)降低,推動地球氣候從“暖室期”進入“冰室期”[3]。這一推測目前還未獲得地質(zhì)記錄的佐證,但是硅藻與氣候系統(tǒng)的相互作用已經(jīng)有了明確證據(jù)。晚新生代硅藻多樣性程度的幾次顯著增加,與始新世、漸新世之交的C3草原擴張,中中新世C4草類出現(xiàn)以及晚中新世C4草原擴張,在時間上都存在著良好的對應關(guān)系[2]。推測草類植被可以加速陸地硅酸鹽風化,而入海溶解硅通量的增加促進了硅藻的分異和繁盛。反過來,海洋硅藻勃發(fā)加劇了碳埋藏以及氣候變冷趨勢,而氣候干旱化和季節(jié)性的增強又有利于草原植被替代森林植被[2,9-10]。因此,氣候、硅藻、陸地草原生態(tài)系統(tǒng)的協(xié)同演化是晚新生代地球表層圈層相互作用的一個精彩例子。
除了在動蕩、富營養(yǎng)鹽水體中快速勃發(fā)的小型硅藻,還有一個重要類群是在層化水體或者洋流鋒面交界處勃發(fā)的大型硅藻。這種大型硅藻生長周期長達數(shù)月,死亡之后可以快速通過水柱沉降在海底,形成限制底棲生物活動的席狀或網(wǎng)狀沉積物,稱為硅藻席沉積(Diatom-mat Deposits)。半個世紀以來這種硅藻席沉積物在全球大洋都有發(fā)現(xiàn)。近年來隨著我國遠洋科考航次的增多,多家海洋研究單位在熱帶-亞熱帶西太平洋水深超過5 000 m 的數(shù)十個站位,都發(fā)現(xiàn)了末次冰期以及冰消期的硅藻席沉積[11]。中文文獻中,對該學術(shù)主題也做過介紹[12-13]。據(jù)估算,大型硅藻勃發(fā)對大洋初級生產(chǎn)力以及有機碳、硅輸出的貢獻并不亞于小型硅藻勃發(fā)[14-15]。但無論現(xiàn)代觀測還是地質(zhì)記錄,對大型硅藻勃發(fā)儲碳作用的研究程度都不充分,這導致許多碳循環(huán)模型和生物地球化學模型并不包含硅藻席模塊。鑒于此,本文從硅藻席沉積時空分布和勃發(fā)機制等角度綜述該主題的研究進展,并嘗試討論硅藻席沉積對軌道時間尺度上全球碳-氮循環(huán)的潛在影響。
1 硅藻席沉積屬種、時空分布及其勃發(fā)機制
1.1 硅藻席屬種
硅藻席主要由大型硅藻組成(圖1),包括海毛藻Thalassiothrixspp.、根管藻Rhizosoleniaspp.、半管藻Hemiaulusspp.、角毛藻Chaetocerosspp.、掌狀冠蓋藻Stephanopyxis palmeriana、圓篩藻Coscinodiscusspp. 和大篩盤藻Ethmodiscus rex等。其中E. rex是熱帶深海硅藻軟泥的常見屬種,根據(jù)Kolbe 的記載[16],最早由科林伍德在孟加拉灣發(fā)現(xiàn)。1952 年英國“挑戰(zhàn)者”號科考船在馬里亞納海溝南部水深10 505 m地方再次獲得含這種硅藻碎片的沉積物,Wiseman與Hendey 將其正式命名為Ethmodiscus rex(Rattray)Hendey[17]。

圖1 常見成席硅藻屬種A:海毛藻屬Thalassiothrix spp..[14], B:角毛藻屬Chaetoceros spp.[18], C:根管藻屬Rhizosolenia spp.(拍攝者Lars Edler, http://nordicmicroalgae.org),D:半管藻屬Hemiaulus spp.[19], E:圓篩藻屬Coscinodiscus spp.[20], F:大篩盤藻Ethmodiscus rex(本次研究,樣品來自西北太平洋馬里亞納海溝挑戰(zhàn)者深淵)。Fig.1 Images of typical mat-forming diatomsA: Thalassiothrix spp.[14], B: Chaetoceros spp.[18], C: Rhizosolenia spp.(photo by Lars Edler, http://nordicmicroalgae.org), D: Hemiaulus spp.[19],E: Coscinodiscus spp.[20], F: Ethmodiscus rex (this study; samples were taken from the Challenger Deep, the Mariana Trench).
不論小型硅藻勃發(fā),還是大型成席硅藻沉降,都容易形成具有紋層構(gòu)造的沉積物(圖2)。這有兩方面的原因:首先,硅藻和陸源輸入物通量隨時間發(fā)生交替變化,同時陸源輸入的營養(yǎng)鹽也會促使硅藻的優(yōu)勢種屬發(fā)生更迭,這些會導致層狀沉積特征的形成。其次,如前所述,成席硅藻大規(guī)模快速沉降在海底形成席狀或者網(wǎng)狀結(jié)構(gòu),即便在底層水充氧條件良好狀況下也會限制底棲生物的掘穴和擾動活動,因此有利于原始層狀沉積構(gòu)造的保存。例如,在加利福利亞灣全新世沉積剖面里,發(fā)現(xiàn)Thalassiothrix longissima硅藻殼體相互鉸接,形成了硅藻席沉積的紋層構(gòu)造[21-22]。

圖2 大洋硅藻席沉積形成的紋層構(gòu)造a:加利福尼亞灣(站位JPC56, 27°30′N、112°6′W, 水深818 m), 主要硅藻屬種是Thalassiothrix spp.[22];b:赤道西太平洋(站位TS01-B10, 11°11.7′N、141°48.7′E, 水深8 638 m),主要硅藻屬種是E.rex(本次研究)。Fig.2 Laminated structure of diatom-mat depositsSamples are from (a) Guaymas Basin, Gulf of California (JPC56, 27.5°N,112.1°W, 818 m water depth). The dominant diatom species was Thalassiothrix spp.[22]; (b) the equatorial Western Pacific (TS01-B10,11°11.7′N, 141°48.7′E, 8638 m water depth). The main diatom species is E.rex (this study).
1.2 硅藻席沉積的時空分布
現(xiàn)代表層沉積物調(diào)查表明,硅藻軟泥(Diatom Ooze,硅藻殼體占比超過30%,由小型硅藻、大型硅藻或者二者混合物構(gòu)成)主要分布在高緯海區(qū),其中3/4 的沉積量分布在南緯50°寬約900~2 000 km的緯向環(huán)帶中;其次分布在北緯40°以北的太平洋阿拉斯加灣、白令海、鄂霍次克海、日本海等海區(qū)[23]。硅藻席沉積除了分布于上述高緯海域外,在中低緯以及赤道海域也大量出現(xiàn)(圖3)。不同海域硅藻席的主要構(gòu)成種屬也存在差別。
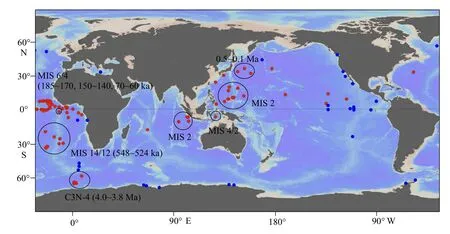
圖3 目前發(fā)現(xiàn)的全球硅藻席沉積的時空分布其中紅色圓點為E. rex 硅藻席, 藍色圓點為其他種屬的硅藻席[14,35-49]。有明確年齡的E. rex 硅藻席沉積用圓圈標注。熱帶-亞熱帶西太平洋和熱帶印度洋E. rex 硅藻席的年齡主要集中在末次冰期[13,17,24-27,50-61], 東太平洋的E. rex 硅藻席沉積大致在上新世和更新世[53,62-64], 大西洋E. rex 硅藻席沉積發(fā)生在中—晚更新世冰期[28-31,65-68], 南極洲附近的E. rex 硅藻席沉積, 根據(jù)古地磁年齡判斷約為4.0~3.8 Ma[32,69-71]。Fig.3 Spatiotemporal distribution of global diatom-mat deposits discovered so farRed dots denote diatom-mat deposits comprised of E. rex, and blue dots represent those comprised of other diatom species[14,35-48]. The E. rex diatom-mat deposits with reliable chronology constraints are circled. The E. rex blooming events in the tropical-subtropical western Pacific and tropical Indian Ocean occurred during the last glacial period[13,17,24-27,50-61], and the blooming in the eastern Pacific occurred in both Pliocene and Pleistocene[53,62-64], while those in the Atlantic occurred across the Mid-Late Pleistocene ice ages[28-31,65-68]. The E. rex blooming event close to Antarctica occurred during 4.0 ~ 3.8 Ma, constrained by paleomagnetic dating[32,69-71].
由于鉆孔調(diào)查資料的限制,目前發(fā)現(xiàn)的硅藻席沉積主要集中在晚第四紀。其中更新世冰期時全球熱帶、亞熱帶海區(qū)出現(xiàn)大規(guī)模的E. rex硅藻席沉積。目前具有可靠定年數(shù)據(jù)的沉積區(qū)域,包括熱帶-亞熱帶西太平洋(氧同位素期次4 和2,MIS 4 和2)[11,24-26]、赤道東印度洋(MIS 2)[27]和熱帶東大西洋(MIS 6 和4)[28-29]以及30°S 附近的亞熱帶南大西洋(MIS 14 和12)[30](圖3)。地質(zhì)歷史時期寡營養(yǎng)熱帶海區(qū)出現(xiàn)大范圍E. rex硅藻席沉積的現(xiàn)象,被稱為“E. rex難題”[31],它的成因迄今仍在爭論。此外,E.rex硅藻席沉積在南極威德爾海區(qū)域也有發(fā)現(xiàn),古地磁方法給出的約束年齡約為4 Ma[32]。
在更老的地層中,硅藻席沉積也有零星報道。例如在更新世北非季風強盛期,地中海出現(xiàn)硅藻席勃發(fā)和腐泥層沉積,主要由rhizosolenid diatom(廣義根管藻,主要包括Rhizosoleniaspp.、Probosciaspp.和Pseudosoleniaspp.等多個屬種)和Hemiaulusspp.組成[33]。15~4.4 Ma 期間,赤道東太平洋區(qū)域深海鉆探鉆孔里也出現(xiàn)數(shù)次成規(guī)模的硅藻席沉積,主要由Thalassiothrixspp.組成[34]。
1.3 大型硅藻勃發(fā)機制
目前通過遙感技術(shù)、沉積物捕獲器以及現(xiàn)場觀測等技術(shù)手段,發(fā)現(xiàn)大型成席硅藻勃發(fā)與沉降常常發(fā)生在兩種水文環(huán)境中,即層化水體或者水團鋒面交匯處(圖4)。熱帶-亞熱帶大洋環(huán)流圈(tropicalsubtropical oceanic gyres)占據(jù)了全球大洋約60%的面積,區(qū)域內(nèi)水體溫度較高,層化較強,次表層營養(yǎng)鹽很難通過垂向運移輸送到表層水體中。再加上遠離大陸,缺乏陸源營養(yǎng)鹽供應,絕大部分熱帶-亞熱帶環(huán)流圈的營養(yǎng)鹽含量和生物量較低[72-73]。但是成席硅藻的一系列特殊本領(lǐng)卻讓它們在這種層化水體中獲得了生存競爭優(yōu)勢。首先,部分成席硅藻可以在弱光環(huán)境下生長,如Rhizosoleniaspp. 和Stephanopyxisspp.[74],因此可以利用深層葉綠素高值區(qū)(Deep Chlorophyll Maxima, DCM,一般在100 m以下水深)中的營養(yǎng)鹽。由于它們生活在表層水團的底部,因此也可以利用風暴等強擾動事件打破溫躍層障礙上涌的營養(yǎng)鹽[14,20,75]。其次,成席硅藻的細胞尺寸較大,可以主動調(diào)節(jié)自身浮力,使其在深層營養(yǎng)源和透光層之間垂向移動,因此既可以利用次表層海水中營養(yǎng)鹽,又可以利用表層海水中的光照條件[76-77]。例如Rhizosoleniaspp.個體可達175 μm,通過相互連接,長度可達30 cm,通過調(diào)控浮力下沉到營養(yǎng)躍層攝入營養(yǎng)鹽后,再上浮到表層進行光合作用[78]。E. rex也可以下潛至水深200 m 左右次表層海水中[31]。可以在弱光條件下生存或者具有浮力機制的成席硅藻也被稱為“樹蔭種”硅藻(Shade Flora Diatoms)[20]。第三,成席硅藻都具有巨大的液泡,可以儲存所攝取的營養(yǎng)鹽,能夠應付長期營養(yǎng)不足的環(huán)境[79]。最后,部分成席硅藻可以與固氮藍細菌共生,獲得氮供應(下文詳述)[80]。因此,在相對寡營養(yǎng)、層化的海洋環(huán)境中,大型成席硅藻可以占據(jù)優(yōu)勢的生態(tài)位[14,20,75,81]。
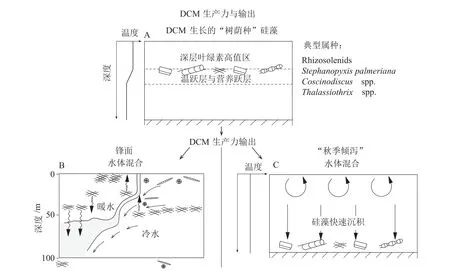
圖4 “樹蔭種”硅藻的生長和沉降機制A:生長于深層葉綠素高值區(qū)的典型“樹蔭種”硅藻, B:鋒面匯聚帶水團相互作用導致的成席硅藻生長和沉降, C:季節(jié)性海洋層化結(jié)構(gòu)破壞造成的硅藻席沉積(改自Kemp and Villareal[15])。Fig.4 Mechanisms for the growth and the deposition of shade flora diatomA: Schematic showing the growth of typical shade flora diatom in the “Deep Chlorophyll Maxima” zone, B: the growth and the deposition of mat-forming diatom in the oceanic frontal convergence zone, C: seasonal breakdown of stratification results in a massive deposition of giant diatom (after Kemp and Villareal)[15].
Kemp 等[14]對季節(jié)尺度海洋表層環(huán)境變化與硅藻勃發(fā)的關(guān)系做過詳細描述。在加利福尼亞灣和地中海,春季時海水混合增強,水體上涌帶來了豐富的營養(yǎng)物,導致小型硅藻迅速生長。春末夏初海洋層化加強,表層營養(yǎng)成分逐漸耗盡,小型硅藻勃發(fā)結(jié)束并快速沉降,以上過程稱為“春季勃發(fā)”(Spring Bloom)[14]。從早夏到晚秋,海洋表層水由于溫度變暖而密度減小,形成較穩(wěn)定的層化結(jié)構(gòu),進入成席硅藻占據(jù)優(yōu)勢的季節(jié)[75]。秋末冬初時隨著上層水體的擾動,成席硅藻大量死亡并造成大規(guī)模堆積的“秋季傾瀉”現(xiàn)象(Fall Dump)[14-15,75]。
此外,Yoder 等[82]在赤道太平洋水團鋒面交匯處也觀測到成席硅藻的勃發(fā)和沉積事件。赤道太平洋葉綠素濃度最高的水體并不在赤道上升流區(qū)域,而是位于與熱帶不穩(wěn)定波(tropical instability waves, TIWs)傳播路徑相吻合的水團鋒面交匯處。鋒面南側(cè)為水溫較低的南赤道流,北側(cè)為水溫較高的北赤道逆流。根管藻(Rhizosoleniaspp.)在暖水一側(cè)大量富集,導致水體呈暗綠色,而冷水一側(cè)硅藻數(shù)量相對較少,水體為淺色。這種水體顏色分界線可以延伸數(shù)百千米。推測大型硅藻可能在鋒面冷水側(cè)吸收和存儲上升流帶來的營養(yǎng)鹽物質(zhì),然后主動遷移到暖水一側(cè)進行生長和勃發(fā)(圖4B),導致顆粒有機碳輸出在熱帶不穩(wěn)定波的活躍期間顯著增強[14,82-84]。
如前所述,晚第四紀冰期熱帶-亞熱帶大洋出現(xiàn)規(guī)模巨大的E. rex勃發(fā)(圖2),但每個海區(qū)的勃發(fā)機制存在差異。西太平洋和赤道東印度洋E. rex勃發(fā)都與大洋層化水體相關(guān)。冰期時亞洲冬季風可以向西太平洋供應風塵硅[11],但印度洋的營養(yǎng)鹽來源存在爭議。Broecker 等認為通過印度尼西亞貫穿流,太平洋溫躍層水團可以向東印度洋輸送硅藻席勃發(fā)所需的營養(yǎng)物質(zhì)[27]。De Deckker 等則認為由于冰期季風降雨減少有助于東印度洋表層形成高鹽度和高營養(yǎng)鹽的環(huán)境,從而促進了硅藻席勃發(fā)[61]。
熱帶-亞熱帶大西洋E. rex席勃發(fā)與洋流鋒面位置的關(guān)系更加密切。在熱帶大西洋,北大西洋中央水團(North Atlantic Central Water, NACW)與南大西洋中央水團(South Atlantic Central Water, SACW)在16°~25°N 位置、水深100~500 m 的地方相遇,形成了佛得角鋒面(Cape Verde Frontal Zone)。該鋒面在冰期時向赤道方向移動了10~15 個緯度[85]。由于西非岸外大量硅藻席沉積站位與該鋒面位置重合,推測硅藻席很可能在鋒面兩側(cè)通過位置遷移發(fā)生勃發(fā)。此外,非洲河流可以向近岸地區(qū)輸入硅藻勃發(fā)所需的硅酸鹽[29]。在亞熱帶南大西洋,E.rex硅藻席勃發(fā)一般被認為與阿古拉斯洋流(Agulhas Current)輸送的營養(yǎng)鹽相關(guān)[86]。
2 現(xiàn)代大洋成席硅藻與碳-氮耦合循環(huán)
據(jù)推測,大型硅藻勃發(fā)可能貢獻了整個大洋約20%的初級生產(chǎn)力和25%的顆粒有機碳輸出[14-15]。雖然有硅藻外殼的保護,硅藻勃發(fā)產(chǎn)生的絕大部分有機碳還是發(fā)生了降解,并不能進入地質(zhì)儲庫中[87]。在全球大洋沉積物調(diào)查中也常常發(fā)現(xiàn),高生物硅含量的深海沉積物往往只有均值水平的有機質(zhì)含量[88]。成席硅藻勃發(fā)可能是通過影響大洋“呼吸碳庫”的方式來調(diào)控碳循環(huán)(詳見第3 部分)。由于成席硅藻碳循環(huán)和硅利用方式已經(jīng)有文獻的詳細介紹,本節(jié)主要聚焦前人較少關(guān)注的氮循環(huán)部分。
2.1 現(xiàn)代大洋氮循環(huán)與同位素分餾
海洋氮循環(huán)可以調(diào)控大洋硝酸鹽和銨鹽的濃度水平,因此是決定海洋初級生產(chǎn)力和生物泵效率的關(guān)鍵要素之一,并與碳循環(huán)過程緊密相連。海洋氮循環(huán)主要包括固氮作用和反硝化作用,此外還包括生物同化作用、再礦化作用和硝化作用等。氮在海洋中的滯留時間約為3 000 年,以硝酸鹽為主,儲量約為6 000 億t[89]。生物固氮作用是大洋氮輸入的最重要途徑,主要發(fā)生在寡營養(yǎng)開闊大洋的表層海水中,依靠自養(yǎng)型的藍細菌完成,每年固氮總量達到約1.63 億t[90]。相比于大氣氮氣的同位素值(定義為0),生物固氮產(chǎn)物的氮同位素相較于大氣氮氣偏負(其值約-2‰~0)[91],低于大洋硝酸鹽的平均氮同位素值(5‰)[92]。
海洋反硝化作用主要發(fā)生在赤道東太平洋、熱帶東南太平洋和阿拉伯海等缺氧海區(qū),將硝酸鹽還原為氮氣[89]。水柱反硝化過程中氮同位素分餾效應比較顯著,分餾幅度達到約25‰,但沉積物中反硝化作用幾乎不產(chǎn)生同位素分餾效應[91]。每年從水柱和沉積物中通過反硝化途徑離開海洋的氮分別約為0.68 和1.32 億t[89-90]。
2.2 成席硅藻與固氮藍細菌共生體
如前所述,部分成席硅藻可以利用次表層海水中的營養(yǎng)鹽,另一部分成席硅藻可以通過與藍細菌共生獲得氮營養(yǎng)[81]。目前已經(jīng)發(fā)現(xiàn)可以形成硅藻-固氮生物共生體(Diatom-diazotroph Associations,DDAs)的成席硅藻包括半管藻、根管藻、角毛藻以及掌狀冠蓋藻等屬[93-94]。固氮藍細菌將大氣中的氮氣轉(zhuǎn)化為可以被寄主硅藻利用的含氮化合物,幫助硅藻突破寡營養(yǎng)海域氮營養(yǎng)的限制。培養(yǎng)實驗發(fā)現(xiàn)共生體中藍細菌約22%的碳元素由宿主硅藻提供,宿主硅藻78%~91%的氮元素來自藍細菌。硅藻細胞越大,固氮和固碳的速率越高[95]。此外,藻類-藍細菌共生體的固氮/固碳效率還受到多種因素的影響,在一定范圍內(nèi),光照、溫度與固氮效率呈正相關(guān)關(guān)系[95]。加入鐵元素和風塵也可以加強藍細菌UCYN-A 對氮氣的固定[96]。硅藻與藍細菌共生體的固氮量十分可觀,在熱帶北大西洋海域,硅藻-藍細菌共生體(Hemiaules-Richelia)與藍細菌中的束毛藻Trichodesmiumspp.一起為表層水提供了近25%的氮營養(yǎng)來源[97]。
2.3 硅藻沉積物氮同位素指標與海洋生產(chǎn)力
氮同位素是追蹤地質(zhì)歷史時期氮循環(huán)過程的最重要工具。但是直接測量海洋沉積物全巖樣品獲得的氮同位素(δ15Nbulk)是一個混合信號,很多時候并不能有效地追蹤氮循環(huán)過程,這是因為從海洋上層掉落的顆粒氮,在水柱中發(fā)生了降解和再礦化作用。在氧化條件下,富含14N 的氨基酸更易被分解,產(chǎn)生的NH4+被黏土吸附,或替代層狀硅酸鹽礦物中的鉀離子[91,98]。這些過程會導致氮同位素發(fā)生分餾。但是在還原條件下,有機質(zhì)降解的同位素分餾效應很小,可以較好地保存海洋有機質(zhì)的初始δ15N 組成[98]。此外,沉積物樣品中有機氮有多個來源,包括陸源輸入和多種海洋浮游藻類的生產(chǎn)力貢獻。
在富含硅藻的沉積物,尤其是硅藻席樣品中,有機質(zhì)主要由勃發(fā)的硅藻產(chǎn)生,來源相對單一,并且硅藻產(chǎn)生的蛋白石可以作為保護有機質(zhì)的儲層,使得硅藻殼體內(nèi)嵌合的有機質(zhì)在沉降和埋藏過程中相對不容易受到成巖作用的影響。因此,富含硅藻沉積物常常是氮同位素測試的良好載體。利用硅藻沉積物樣品,可以測試有機氮同位素(δ15Norg)、葉綠素(Chlorophyll)氮同位素(δ15Nchl)、硅藻內(nèi)嵌(Diatom-bound)有機質(zhì)氮同位素(δ15Ndb)等指標。根據(jù)室內(nèi)培養(yǎng)實驗結(jié)果,發(fā)現(xiàn)δ15Norg與δ15Ndb變化趨勢一致,但二者同位素值存在偏移,δ15Ndb總是小于δ15Norg,偏差幅度在1.0‰~14.0‰之間。綜合來看,利用同一種屬硅藻測試的δ15Ndb指標,是反演表層水體硝酸鹽利用程度以及古生產(chǎn)力變化最為可靠的指標[99]。
利用硅藻沉積物氮同位素已經(jīng)成功回答了地質(zhì)歷史時期海洋生產(chǎn)力變化的一些重要問題。例如,東地中海海底存在著名的腐泥層沉積,有機碳含量可以達到5%。一般認為這些腐泥層形成于北半球夏季太陽輻射量的高值期和北非季風的強盛期(例如早全新世和末次間冰期),一方面非洲尼羅河泛濫和營養(yǎng)鹽輸入導致地中海生產(chǎn)力勃發(fā),另一方面淡水注入后打斷了地中海表層海水的潛沉,底層海水通風減弱逐漸失去氧氣,導致有機質(zhì)在缺氧條件下得到較好的保存[33]。進一步分析發(fā)現(xiàn),腐泥層沉積中事實上存在大量的成席硅藻Hemiaulusspp.和rhizosolenid 硅藻[100],硅藻勃發(fā)是當時海洋生產(chǎn)力的主力。通過測試氮同位素,發(fā)現(xiàn)在上層海水層化條件加強的情況下,成席硅藻是通過利用次表層營養(yǎng)鹽實現(xiàn)大規(guī)模勃發(fā)的,并與固氮藍細菌共生形成廣泛的固氮作用[15,101]。這糾正了之前的認識,即非洲季風強盛并沒有造成地中海表層水的富營養(yǎng)化[101]。值得一提的是,白堊紀時特提斯洋洋底也沉積了大規(guī)模富含有機碳的黑色頁巖,同樣也是成席硅藻Hemiaulusspp.和rhizosolenid 硅藻勃發(fā)造成的[102]。
3 大規(guī)模硅藻席沉積與大洋碳-氮循環(huán)
3.1 冰期熱帶硅藻席沉積與全球碳循環(huán)
第四紀大氣CO2濃度在冰期-間冰期旋回過程中有80~100 p.p.m.v (Parts Per Million by Volume,百萬分比濃度)波動,其變化機制是學術(shù)界長期探索的課題。一般認為,大氣CO2濃度變化主要是通過調(diào)節(jié)深海碳儲量來實現(xiàn)的[103]。由于冰期大洋內(nèi)部通風速率下降,再礦物作用產(chǎn)生的“呼吸碳”(Respired Carbon),可以更長久地滯留在大洋內(nèi)部,引起了大洋無機碳含量上升和溶解氧濃度下降[104-105]。冰消期時,在南大洋附近隨著深部海水上翻,大洋內(nèi)部“老碳”釋放并再次進入大氣圈中[106]。
過去認為冰期深海內(nèi)部發(fā)生降解的顆粒有機碳主要來自表層生產(chǎn)力輸出,但晚第四紀冰期熱帶大洋廣泛存在的E. rex成席硅藻勃發(fā)事件(圖3),說明在大洋寡營養(yǎng)海區(qū)的次表層海水中,也存在規(guī)模可觀的生產(chǎn)力輸出事件。據(jù)估算,在熱帶西太平洋,E. rex勃發(fā)事件期間的海洋初級生產(chǎn)力與近岸上升流地區(qū)水平一致[87]。不過硅藻席沉積物的有機碳含量僅為0.2%~0.3%[87],說明硅藻勃發(fā)產(chǎn)生的絕大部分有機質(zhì)在水柱以及水-沉積物界面處發(fā)生降解,成為冰期大洋內(nèi)部的“呼吸碳”,因此冰期E.rex勃發(fā)充當了從大氣向深海內(nèi)部運輸碳的重要角色。但成席硅藻勃發(fā)在碳循環(huán)中的角色長久被忽略,未來需要進一步調(diào)查,厘定冰期E. rex勃發(fā)事件規(guī)模及其對大洋碳存儲的影響。
此外,在冰期-間冰期時間尺度上,海洋固氮作用與反硝化作用的比例波動調(diào)控了海洋氮儲庫總量和大洋初級生產(chǎn)力,它們也可以影響到大氣CO2濃度[2,89,107]。一般認為,冰期時大陸架暴露、赤道上升流增強等因素,可以增加磷元素供應,刺激固氮作用[107-109]。同時冰期水體混合增強,溶解氧含量增加,總體上會削弱反硝化作用[110]。因此,冰期大洋氮儲庫總量水平上升。模擬實驗也表明,相比于全新世,末次冰盛期時全球海洋氮儲庫最多可能增加了一倍,即從6 000 億t 提高到1.2 萬億t[111]。同樣地,這些研究忽略了冰期E. rex勃發(fā)事件對大洋固氮作用的影響。目前對冰期E. rex勃發(fā)機制和營養(yǎng)鹽來源還存在爭論。在熱帶西太平洋,硅同位素示蹤研究結(jié)果表明,E. rex勃發(fā)利用的是亞洲內(nèi)陸的風塵硅輸入[11]。根據(jù)現(xiàn)代生態(tài)調(diào)查,E. rex并不能與固氮藍細菌共生,很可能是通過浮力機制利用了次表層水體中的營養(yǎng)鹽,包括硝酸鹽和磷酸鹽等,因此獲得了生存競爭優(yōu)勢并觸發(fā)了勃發(fā)事件。冰期熱帶大洋E. rex大規(guī)模勃發(fā)提高了上層海洋營養(yǎng)鹽的周轉(zhuǎn)速率和利用水平,因此可能刺激廣大寡營養(yǎng)海區(qū)固氮微生物的生產(chǎn)力,從而提升大洋的固氮作用。
3.2 硅藻席沉積與大洋碳同位素重值現(xiàn)象
新生代以來,大洋無機碳碳同位素(δ13C)最顯著的軌道周期就是40 萬年長偏心率周期[112]。最近5 Ma 以來,全球大洋δ13C 波動一共出現(xiàn)了約13 次長偏心率周期,在每個偏心率低值期,都會出現(xiàn)δ13C 偏重現(xiàn)象,被命名為碳同位素重值期(δ13Cmax)。其中,近1.6 Ma 以來,大洋δ13C 波動不再嚴格跟隨天文長偏心率周期,而是拉長成為延續(xù)時間達到50 萬年的事件[113-114]。最近兩次碳同位素重值事件分別發(fā)生在氧同位素13 期(δ13Cmax-II, 0.53~0.47 Ma)和3-1 期(δ13Cmax-I, 0.05~0 Ma)(圖5)。先前研究已經(jīng)注意到,這兩次碳同位素重值事件都伴隨著大洋硅藻席勃發(fā)事件。首先,氧同位素14 和12 期時亞熱帶南大西洋發(fā)生E. rex勃發(fā)事件[30],對應于δ13Cmax-II事件。其次,末次冰期熱帶西太平洋-印度洋以及赤道東大西洋E. rex勃發(fā)事件[11,24-29],對應于δ13Cmax-I事件(圖5)。目前還未查清E. rex勃發(fā)與大洋碳同位素重值信號是否存在機制上的關(guān)聯(lián)。
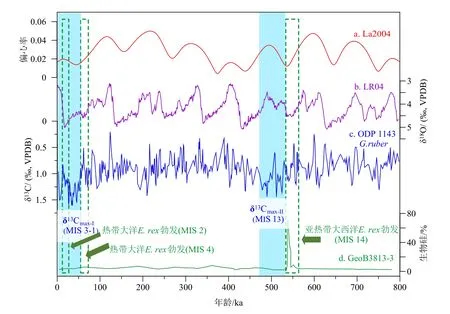
圖5 80 萬年以來兩次大洋碳同位素重值事件與成席硅藻勃發(fā)事件的可能關(guān)聯(lián)a:地球偏心率變化來自La2004[115], b:全球底棲有孔蟲氧同位素合成曲線來自LR04[116], c:南海ODP 1143 站位浮游有孔蟲δ13C 記錄(9°22′N、113°17′E, 水深2 772 m)[117], d:亞熱帶南大西洋GeoB3813-3 站位生物硅含量變化(32°16′S、 21°58′W, 水深4 331 m)[118]。藍色陰影標示碳同位素重值事件,綠色虛線框標示大洋硅藻席勃發(fā)事件。Fig.5 Potential linkage between oceanic δ13Cmax events and diatom blooming events over the past 800 thousand yearsa: The Earth’s orbital eccentricity changes (from La2004) [115]; b: global stack of benthic foraminiferal δ18O, LR04[116]; c: the δ13Cmax events represented by planktonic foraminifera δ13C of ODP Site 1143, southern South China Sea (9°22′N、113°17′E, water depth 2 772 m)[117]; d: biogenic silica concentration(SiO2%) of core GeoB3813-3, South Atlantic (32°16′S、 21°58′W, water depth 4 331 m)[118]. Blue rectangles indicate the duration of the δ13Cmax events and green dotted line boxes show the mat-forming diatom blooming events.
為了解釋大洋δ13C 的長偏心率周期,先前研究提出“溶解有機碳假說”[114]。現(xiàn)在全球海洋溶解有機碳庫總儲量為6 620 億t,與大氣碳儲庫7 500 億t接近[119]。并且海洋溶解有機碳庫中的95%為惰性溶解有機碳[120],在海洋中滯留長達6 000 年[121]。因此,海洋溶解有機碳庫變化可以大幅度影響到大氣CO2濃度以及海洋無機碳δ13C 值。根據(jù)假說,在偏心率低值期,由于熱帶季節(jié)性和季風減弱,入海營養(yǎng)鹽輸入和海洋真核藻類生產(chǎn)力減弱,導致大洋生產(chǎn)力以原核類生物為主,海洋溶解有機碳庫儲量擴大,扣押了大量12C,導致海洋無機碳δ13C 值上升[114]。
由于硅藻席勃發(fā)時,大量有機質(zhì)在水柱中發(fā)生了降解,只有極少部分有機質(zhì)進入埋藏的海洋沉積物中[87]。推測硅藻席有機質(zhì)降解產(chǎn)物中,除了一部分形成無機形式的“呼吸碳”,還有一部分被微生物轉(zhuǎn)化,形成溶解有機碳。這兩種碳的賦存形式對大洋無機碳δ13C 值的作用剛好相反。“呼吸碳”的同位素值較輕,會降低大洋無機碳的δ13C 值;但如果形成溶解有機碳,相當于從其他儲庫中帶走12C,最終使得大洋無機碳δ13C 值升高。因此,硅藻席勃發(fā)如何影響偏心率低值期的大洋無機碳δ13C 重值信號,還需要大量記錄的約束以及模擬的驗證。上的關(guān)聯(lián)。未來更深入的研究需要對地質(zhì)歷史時期的硅藻席勃發(fā)和沉積事件進行更廣泛的調(diào)查,再結(jié)合模型來定量約束硅藻席勃發(fā)事件在軌道尺度的大洋碳-氮循環(huán)中的作用。
4 結(jié)論與展望
(1)成席硅藻在層化或鋒面環(huán)境中勃發(fā)可能與其獨特的生理結(jié)構(gòu)密切相關(guān)。硅藻在海洋浮游藻類演化序列上出現(xiàn)和分異的時間較晚,導致硅藻具備了與藍細菌共生和尿素循環(huán)等特殊功能。成席硅藻更是進一步發(fā)展出大液泡,可以通過浮力調(diào)控機制利用次表層水體中的營養(yǎng)鹽,從而在面積廣闊的熱帶-亞熱帶層化和寡營養(yǎng)海洋中獲得了生存競爭優(yōu)勢,形成大規(guī)模勃發(fā)事件。
(2)硅藻席勃發(fā)和沉積過程可能促進了冰期大氣二氧化碳濃度的降低。一方面,成席硅藻席勃發(fā)過程中,由于絕大部分有機質(zhì)在水柱中發(fā)生了降解,推測可以貢獻冰期深海內(nèi)部的“呼吸碳庫”。另一方面,硅藻席勃發(fā)還可以提高海洋上層營養(yǎng)鹽的周轉(zhuǎn)速率,很可能導致冰期大洋氮儲庫容量和表層生產(chǎn)力的擴增。這兩個過程都可以促進冰期大氣二氧化碳濃度的降低。
(3)成席硅藻勃發(fā)事件的氣候意義并沒有得到充分的重視和理解。在晚第四紀冰期階段,成席硅藻E. rex在熱帶-亞熱帶大洋發(fā)生大規(guī)模的勃發(fā)和埋藏,向大洋深部和海底沉積物輸送了大量有機碳和生物硅,但現(xiàn)在的碳循環(huán)模型以及海洋生物地球化學模型都缺少硅藻席模塊。
(4)低緯大洋E. rex的勃發(fā)事件可能與大洋無機碳碳同位素重值事件相關(guān)。50 萬年以來海洋無機碳碳同位素出現(xiàn)兩次重值事件,期間都伴隨低緯大洋E. rex的成規(guī)模勃發(fā),推測兩者之間存在成因
