大麥馬鈴薯間作模式對大麥苗抗氧化酶活性的影響
2024-03-23 08:04:50楊麗娥楊曉夢普曉英霞官秀嬌楊硯斌楊加珍和習瓊曾亞文
中國種業 2024年3期
趙 塔 楊麗娥 楊曉夢 普曉英 李 霞官秀嬌 楊硯斌 楊加珍 和習瓊 曾亞文
(1 云南大學資源植物研究院,昆明650504;2 云南省農業科學院生物技術與種質資源研究所,昆明 650205;3 云南省麗江市農業科學研究所,麗江674100)
大麥(Hordeum vulgareL.)屬于禾本科作物,具有耐鹽堿、耐干旱、耐貧瘠、適應性廣、分蘗力強等特點[1-2]。作為重要的飼用和釀造原料,大麥不僅有多種生物活性和保健功能[3-5],而且是酶類物質種類和酶活力較高的谷物之一[6],抗氧化酶在大麥的生長發育、抗病和抗脅迫等方面起到重要的調節作用[7]。麥苗內較高的抗氧化酶類物質可以有效清除植物受環境脅迫而積累的有害氧自由基[8],抗氧化酶系統主要包括超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)、多酚氧化酶(PPO)等。而這些抗氧化酶往往與植物體次級代謝物的合成有關,如POD 和PPO 在酚類物質的合成和分解代謝過程中起著重要的作用[6]。因此,分析大麥幼苗抗氧化酶活性對大麥產量及品質保證具有重要意義。
云南屬于青藏高原延伸的低緯高原,是發展大麥及其麥苗生產生態條件最好的地區之一[9],但存在大麥良種缺乏,平均單產較低等問題[10]。而作物之間配套種植可以提高資源利用率、增加作物生物產量以及經濟效益[11],還可以控制或減輕某些病蟲害的發生[12]、改變作物成分及含量[13]、控制雜草生長[14]等。大麥和馬鈴薯具有相似的生態適應性,研究大麥—馬鈴薯間作體系可為緩解大麥原料短缺或功能大麥開發起到積極的作用,保障兩種作物產業的發展[15]。間套作模式下,作物間對水、肥、光、熱資源產生競爭機制,作物體內抗氧化酶系統將作出相應的改變。研究發現,玉米—大豆間作模式下玉米抗氧化酶活性顯著提高,根系抗氧化能力提高,生理活動能力強[16];生姜—葡萄立體間作模式下生姜葉片中抗氧化酶活性顯著提高,可以緩解夏季生姜受到的強光脅迫,降低葉片灼傷率[17]。
目前,關于作物葉片抗氧化酶活性的研究大多是在脅迫條件下進行,且主要集中于小麥、水稻、玉米等作物[18]。李玉賢等[15]已對大麥馬鈴薯套作進行相關研究,但麥薯間作下對大麥抗氧化酶活性影響的研究還未見報道。本研究選用64 個云南大麥品種(系)為試驗材料,對不同種植模式、不同棱型和用途類型分蘗期大麥苗SOD、CAT、POD 以及PPO活性進行比較,分別探究間作—單作、二棱—多棱和啤酒大麥—飼料大麥—青稞之間分蘗期大麥苗抗氧化酶活性的差異,篩選與馬鈴薯間作條件下抗氧化性強的云南大麥品種(系),以期為培育間作模式下抗氧化能力強的大麥新品種提供種質資源。
1 材料與方法
1.1 試驗材料供試材料為64 個云南大麥優良品種(系)(表1),由云南省農業科學院生物技術與種質資源研究所選育或收集,根據棱型分為二棱大麥39 份、多棱大麥25 份;根據用途類型分為啤酒大麥36 份、飼料大麥18 份、青稞10 份。所選用的馬鈴薯品種為麗江市農業科學研究所選育的高花青素含量的紫色中晚熟馬鈴薯品種(編號:12H11-3)。
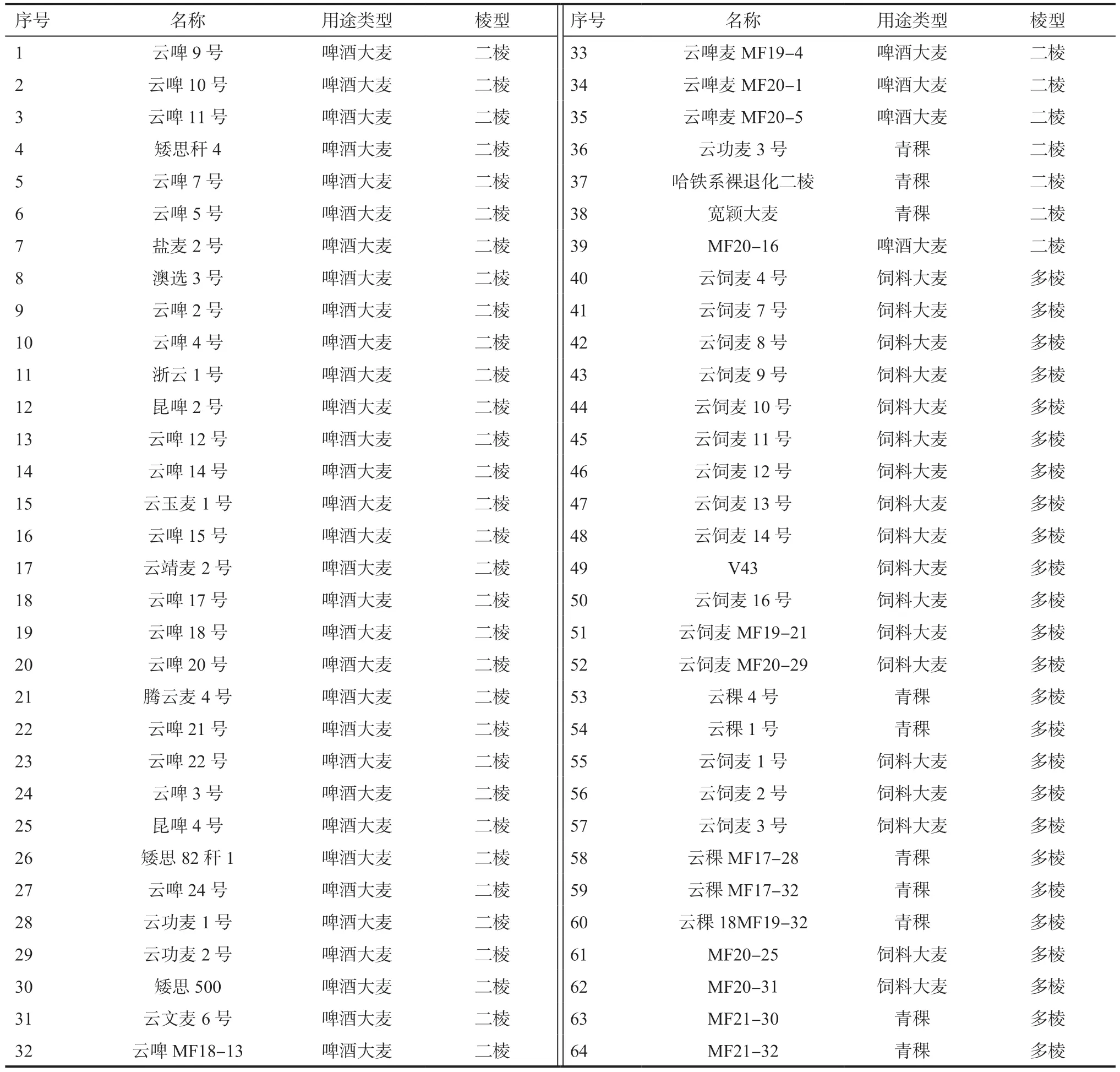
表1 64 份云南大麥優良品種(系)
1.2 試驗設計試驗地點位于昆明市盤龍區滇源鎮南營村試驗基地(海拔1973m),試驗地土壤類型為紅壤土,其理化性質為:pH 值 6.25,有機質含量25.3%,全氮含量1.81%,全磷含量1.47%,堿解氮含量102.31mg/kg,有效磷含量14.32mg/kg,速效鉀含量147.13mg/kg。地勢平坦,肥力均勻,茬口一致,無遮陰,排灌良好,田間水肥按大田正常管理。
種植方式:試驗采用單因素完全隨機設計,以大麥單作為對照,利用64 個不同大麥品種與馬鈴薯進行間作,即大麥單作和大麥—馬鈴薯間作。(1)大麥—馬鈴薯間作,馬鈴薯起壟種植,壟寬0.5m,壟間距為0.3m,馬鈴薯株距為0.24m;每個小區種植1個大麥品種,行數2 行,行長1.44m,行間距0.25m,面積0.36m2,不同品種大麥小區前后間距為0.24m;馬鈴薯壟間種植大麥。(2)大麥單作:其種植規格同間作。大麥與馬鈴薯于2023 年1 月4-5 日同期播種,大麥播種量為150 粒/行。
1.3 樣品采集與測定
1.3.1 樣品采集于分蘗期對每個小區選取標記植株3 株,分別采集新鮮大麥苗,分裝入凍存管中,管壁做標記后立即存放在-80℃低溫冰箱中備用。
1.3.2 樣品測定及方法大麥苗的POD、CAT、SOD 和PPO 活性分別使用POD、CAT、SOD 和PPO 檢測試劑盒(由南京建成生物工程研究所提供)測定,儀器為美國生產的多功能酶標儀(型號:BioTeK SynergyTMHT)。各酶活性測定均按試劑盒步驟,依次配置所需試劑,SOD 活性在450nm 處測定,CAT 活性在405nm 處測定,POD 活性在420nm處測定,PPO 活性在420nm 處測定。根據試劑盒上的公式進行相關計算,試驗數據均為3 次重復的平均值。
1.4 數據分析采用Excel 2016 和SPSS 26.0 軟件進行數據整理分析;隸屬函數值計算方法參照張會麗等[19]的方法,具體如下。
(1)當指標性狀值與抗氧化性呈正相關時:
(2)當指標性狀值與抗氧化性呈負相關時:
(3)總隸屬函數的計算:
式中,i:某個品種;j:某個性狀;n:指標性狀數量;X ij為隸屬函數值;Xij為性狀值;Xj min為性狀最小值;Xj max為性狀最大值;Xi為品種抗氧化隸屬函數平均值。把每個品種的各個性狀具體抗氧化隸屬函數值累加后求平均值,平均值越大,抗氧化性則越強。
2 結果與分析
2.1 不同種植模式大麥苗抗氧化酶活性差異如表2 所示,不同種植模式對大麥苗抗氧化酶活性有顯著影響(P<0.05)。結果表明,64 個大麥品種(系)大麥苗SOD、CAT、POD、PPO 活性呈現出間作高于單作的普遍趨勢(圖1),且差異達到顯著水平。例如間作模式下云啤17 號大麥苗的SOD 酶活性顯著上升了41.34%;云啤18 號大麥苗的CAT 酶活性顯著上升了75.35%;云啤麥MF20-5 大麥苗的PPO 酶活性顯著上升了21.74%;云啤17 號大麥苗的POD 酶活性顯著上升了54.89%。

圖1 不同種植模式下大麥苗抗氧化酶箱形圖
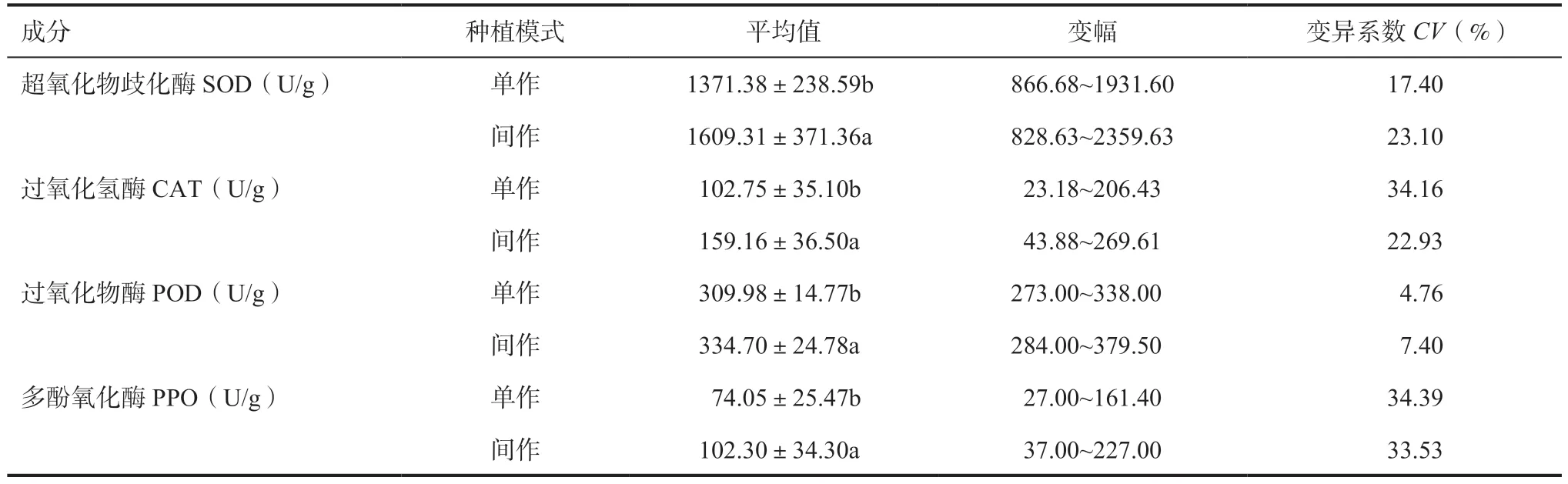
表2 不同種植模式大麥苗抗氧化酶活性
此外,不同品種在相同模式下抗氧化酶活性均存在廣泛的差異。其中,單作時MF20-16 SOD活性最高為1931.60U/g,云飼麥4 號活性最低為866.68U/g,兩者相差2.23 倍;間作時云啤麥MF20-5 SOD 活 性 最 高 為2359.63U/g,云 飼 麥8 號 活 性最低為828.63U/g,兩者相差2.85 倍。單作時云稞18MF19-32 CAT 活性最高為206.43U/g,V43 CAT活性最低為23.18U/g,兩者相差8.91 倍;間作時MF20-16 CAT 活性最高為269.61U/g,云飼麥4 號CAT 活性最低為43.88U/g,兩者相差6.14 倍。單作時浙云1 號 POD 活性最高為338.00U/g,云飼麥9 號活性最低為273.00U/g,兩者相差1.24 倍;間作時云啤麥MF20-5 POD 活性最高為379.50U/g,云飼麥8 號活性最低為284.00U/g,兩者相差1.34 倍。單作時浙云1 號 PPO 活性最高為161.40U/g,云飼麥10 號活性最低為27.00U/g,兩者相差5.98 倍;間作時云啤12 號PPO 活性最高為227.00U/g,云飼麥11 號活性最低為37.00U/g,兩者相差6.14 倍。說明不同品種(系)大麥抗氧化酶活性差異較大。
2.2 不同棱型大麥苗抗氧化酶活性差異試驗結果表明(表3),不同棱型大麥苗SOD、POD 和PPO 活性水平差異顯著,相同種植模式下二棱大麥苗的SOD、POD 和PPO 平均活性顯著高于多棱;SOD 和PPO 活性呈現出二棱間作>二棱單作>多棱間作>多棱單作的趨勢,POD 則表現為二棱間作>多棱間作>二棱單作>多棱單作。相同種植方式下二棱大麥苗的CAT 活性高于多棱,但差異不顯著,間作二棱大麥酶活性最高(165.05U/g),單作多棱大麥酶活性最低(94.17U/g)。總體來看,不同棱型大麥苗抗氧化酶活性表現為間作二棱大麥抗氧化酶活性最高,單作多棱大麥抗氧化酶活性最低(圖2)。
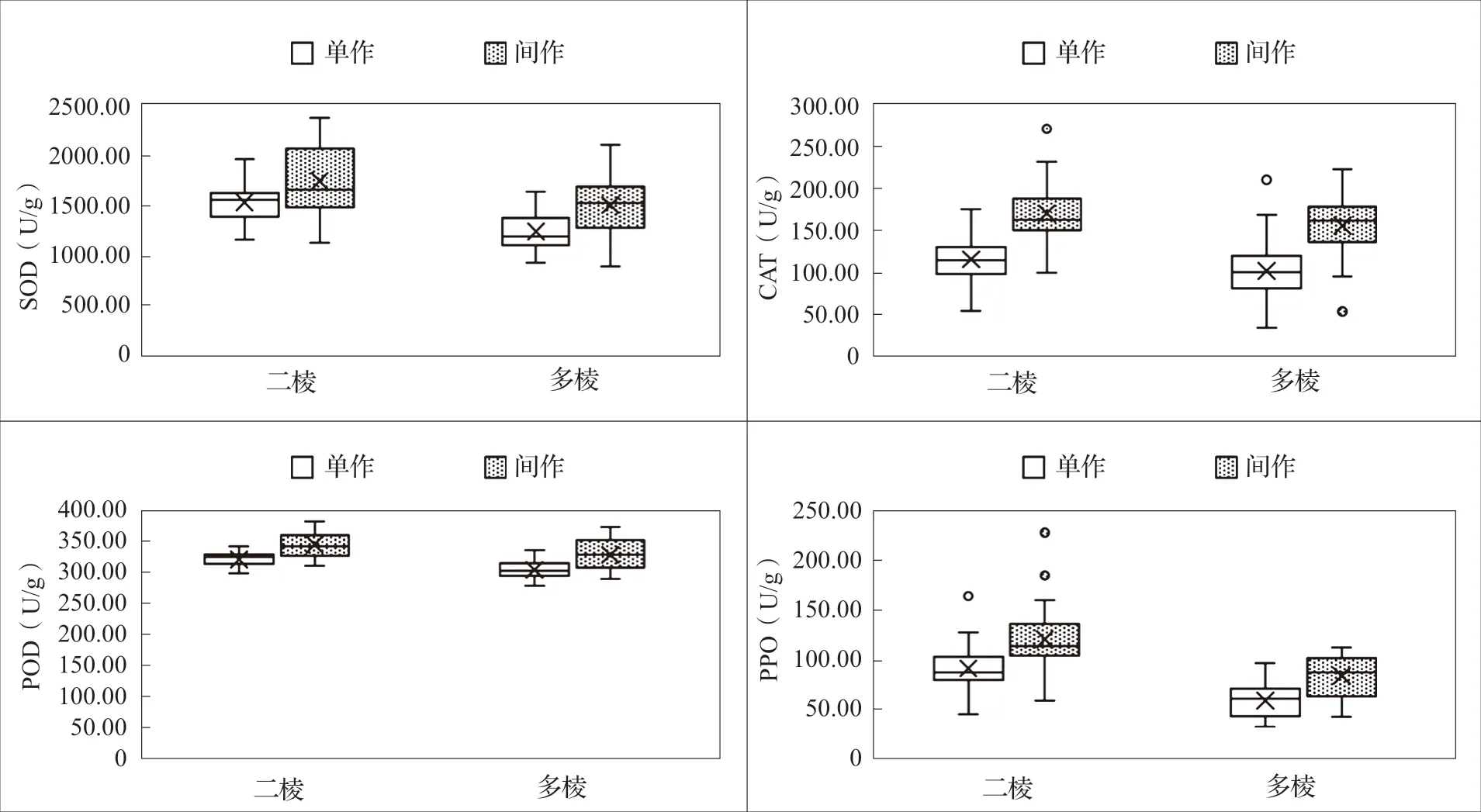
圖2 不同棱型大麥苗抗氧化酶活性箱形圖
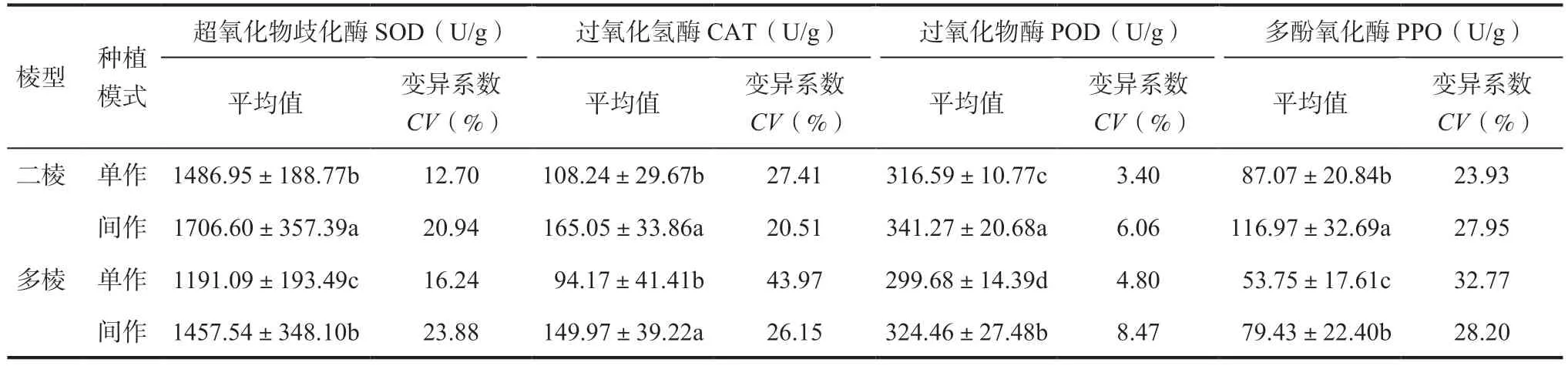
表3 不同棱型大麥苗抗氧化酶活性
2.3 不同用途類型大麥苗抗氧化酶活性差異將供試大麥按用途類型分為啤酒大麥、飼料大麥及青稞,結果表明(表4),不同用途類型大麥SOD 活性水平差異顯著,相同種植模式下飼料大麥SOD 活性顯著低于啤酒大麥和青稞,啤酒大麥和青稞SOD活性水平差異不顯著;其中,間作啤酒大麥SOD 活性含量最高(1715.47U/g),單作飼料大麥SOD 活性含量最低(1130.02U/g),不同種植方式下SOD活性表現出間作啤酒大麥>間作青稞>單作啤酒大麥>間作飼料大麥>單作青稞>單作飼料大麥。

表4 不同用途類型大麥苗抗氧化酶活性
不同用途類型大麥PPO 活性差異顯著,相同種植方式下啤酒大麥PPO 活性顯著高于飼料大麥和青稞,且啤酒大麥、飼料大麥和青稞間差異均達到顯著水平;間作啤酒大麥PPO 活性最高(118.89U/g),單作飼料大麥PPO 活性最低(46.91U/g),兩者相差2.53 倍;不同種植方式下PPO 活性表現出間作啤酒大麥>間作青稞>單作啤酒大麥>間作飼料大麥>單作青稞>單作飼料大麥。
不同用途類型大麥CAT、POD 活性差異不顯著;其中,間作青稞CAT 活性最高(168.87U/g),單作飼料大麥CAT 活性最低(89.24U/g),兩者相差1.89倍;間作啤酒大麥POD 活性含量最高(341.04U/g),單作飼料大麥POD 活性含量最低(296.00U/g),兩者相差1.15 倍;不同種植方式下CAT 活性表現出間作青稞>間作啤酒大麥>間作飼料大麥>單作青稞>單作啤酒大麥>單作飼料大麥;POD 活性表現出間作啤酒大麥>間作青稞>間作飼料大麥>單作啤酒大麥>單作青稞>單作飼料大麥。整體來看,不同類型大麥間作時啤酒大麥品種抗氧化酶活性最高,青稞品種次之,飼料大麥品種抗氧化酶活性最低(圖3)。
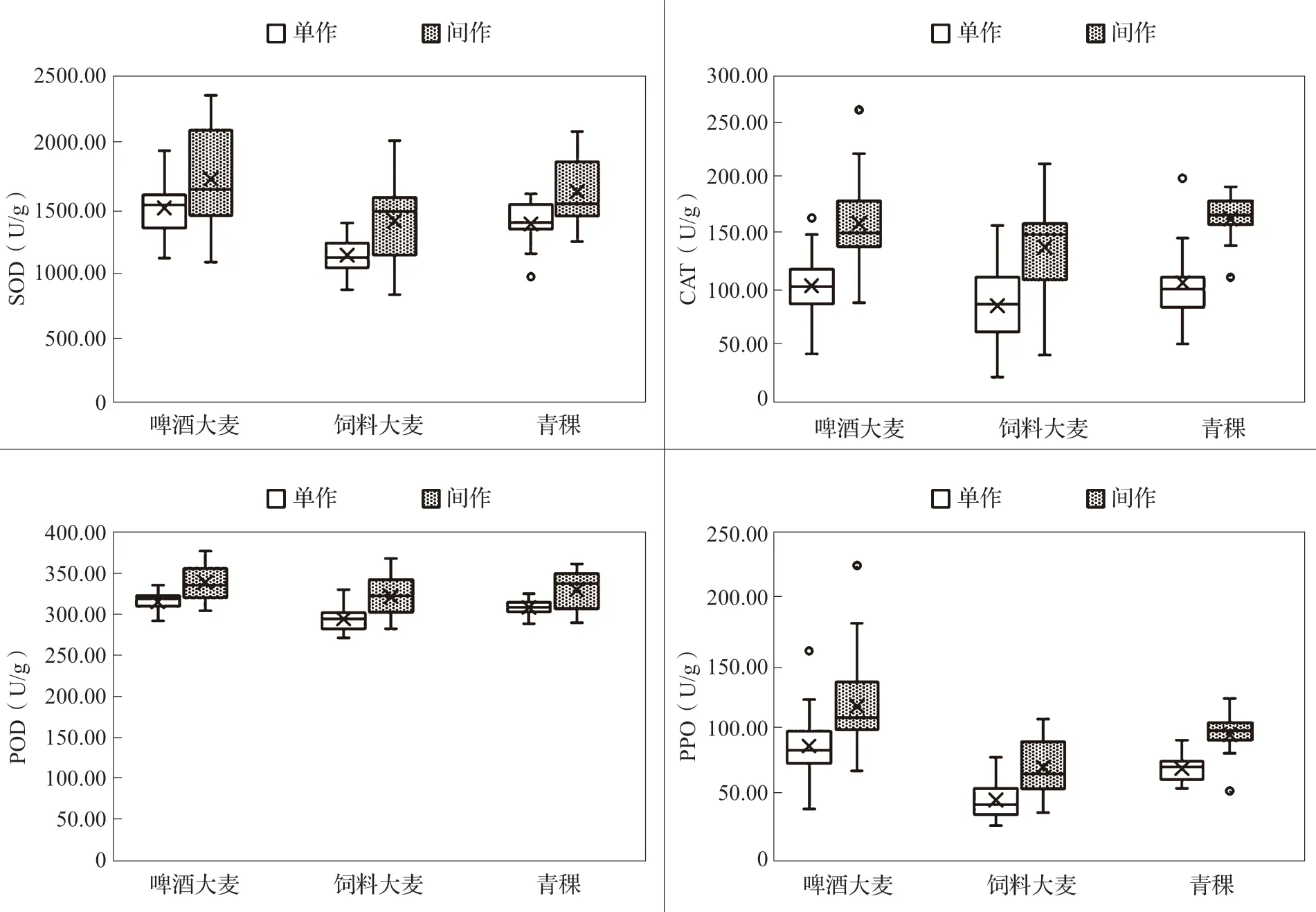
圖3 不同用途類型大麥苗抗氧化酶活性箱形圖
2.4 不同種植模式大麥苗抗氧化酶活性相關性分析相關性分析結果顯示(表5),相同種植模式下抗氧化酶活性相關性趨勢一致。單作種植模式下SOD 與CAT、POD、PPO 活性呈極顯著正相關關系,CAT 活性和PPO 活性呈正相關,相關性不顯著。間作種植模式下除CAT 活性與PPO 活性呈顯著正相關外,其他均呈極顯著正相關。相關性分析結果表明抗氧化酶活性之間相互影響,說明各抗氧化酶共同作用,以響應外界環境的變化,進而保護植物免受氧化應激,并維持正常生長發育。
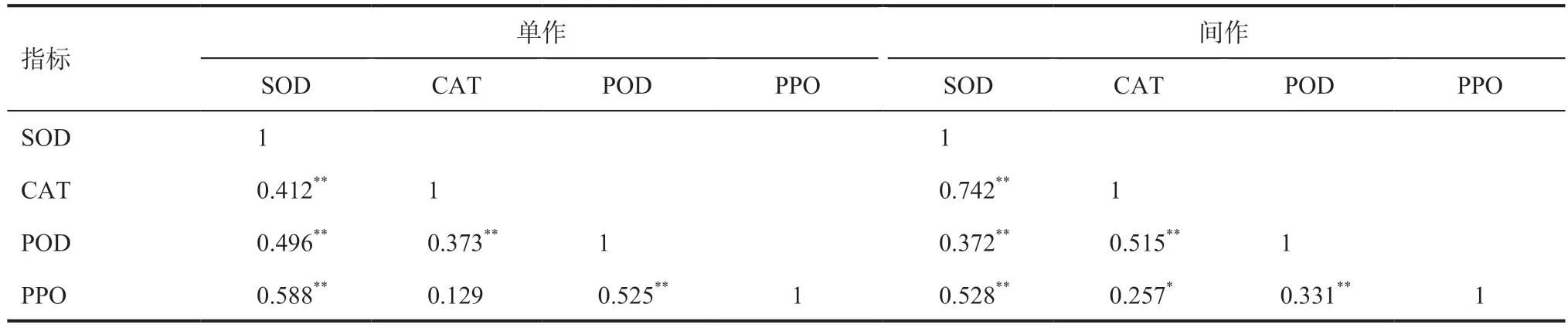
表5 不同種植模式大麥品種抗氧化酶活性相關性分析
2.5 間作種植模式下大麥品種抗氧化性綜合評價對間作種植模式下64 份大麥品種(系)進行隸屬函數值計算,篩選出了抗氧化能力較優的10 個大麥品種(表6)。
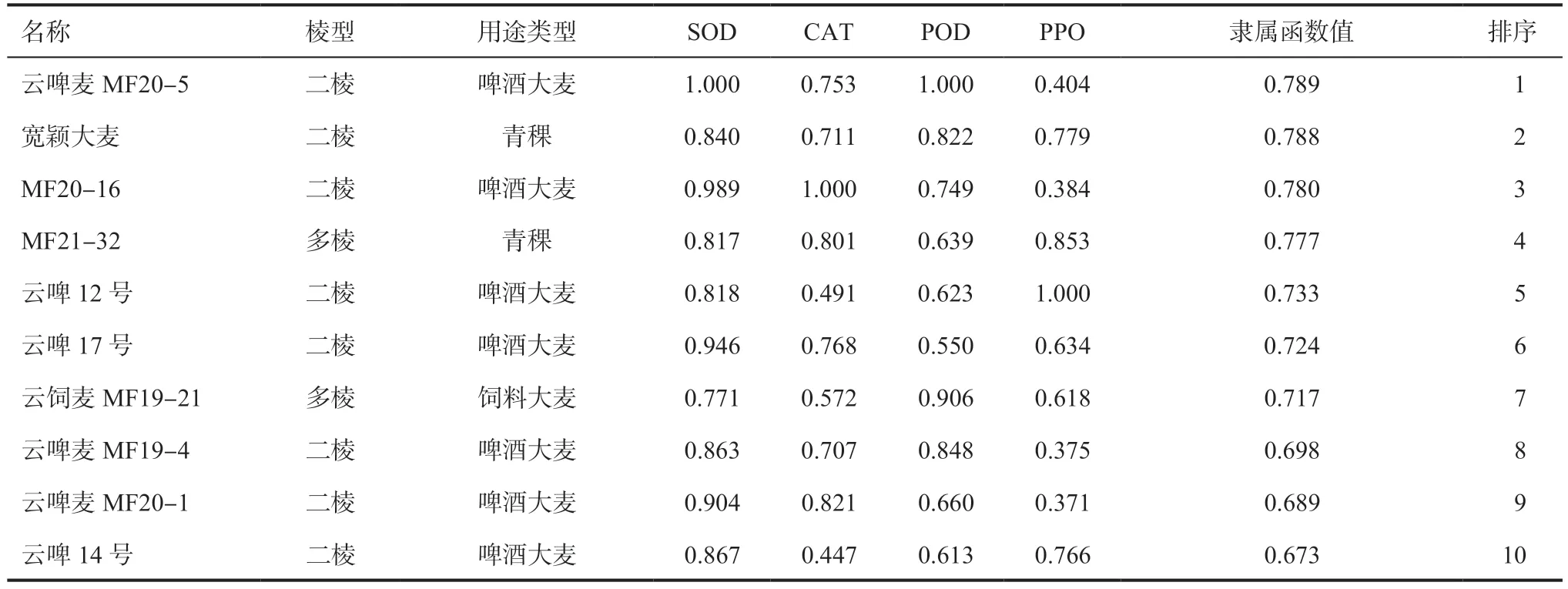
表6 間作種植下大麥品種的綜合抗氧化隸屬函數值(綜合前10)
3 討論
影響植物抗氧化酶活性的主要因素有基因型和外部環境,基因型決定其內在品質,而外部環境主要有生長、種植條件以及氣候等[6]。外部環境改變時,植物體內的防御系統會通過協同抗氧化酶,維持自由基產生與清除的動態平衡,而較高的抗氧化酶活性可以增強大麥植株抵抗外界脅迫的能力[20]。
研究發現,間作體系中由于作物基因型不同、高低不等、受熱受光不均,導致兩種作物在生長過程中存在光熱資源的競爭[21]。本研究中,馬鈴薯間作大麥誘導了分蘗期大麥SOD、CAT、POD、PPO 活性水平顯著升高,這與Assadi 等[22]對大麥鷹嘴豆間作、楊娜娜等[23]對燕麥馬鈴薯間作研究結果一致。大麥—馬鈴薯間作系統中存在地上部光熱水氣資源以及地下部大麥—馬鈴薯根系間營養、水分的資源競爭[24-25]。說明間作種植時可對大麥造成氧化脅迫,從而造成大麥苗活性氧數量增加。大麥苗在外界環境改變時進行脅迫保護,體內的抗氧化酶活性隨之變化。
大麥根據穗的棱型可分為二棱和多棱,不同棱型大麥品種的生長規律、代謝特點存在明顯差異[26],二棱大麥農藝性狀[27]、營養成分含量[28]優于多棱。王美琦等[29]對二棱大麥材料進行抗氧化能力綜合評價,發現大麥種質間的抗氧化性存在顯著差異。有研究表明,多棱大麥多為春大麥,宜播期在3-4 月,而二棱大麥多為冬大麥,宜播期在11 月至翌年1 月左右[30]。本研究發現二棱大麥在不同種植模式下抗氧化酶活性都高于多棱,這可能與播種季節及二棱大麥抗性有關,其具體機制需要進一步研究。
酚類化合物在植物體內積累有助于植物適應強紫外線輻射、溫度極端變化和其他非生物脅迫的惡劣環境[31],也是大麥中的主要生物活性成分之一,會直接影響大麥芽和麥汁的品質,進而會影響成品啤酒的色澤、風味和非生物穩定性[32]。本研究結果表明,啤酒大麥抗氧化酶活性高于青稞和飼料大麥,而抗氧化酶POD 和PPO 在酚類物質的合成和分解代謝過程中起著重要的作用。此外,有研究表明,啤酒大麥根系發達,具有極強的分蘗性,表現出更好的抗旱和豐產穩產能力[33]。青稞由于其特殊的分布環境,使得其具有較好的適應高原惡劣環境的遺傳基礎,在激素信號轉導、抗病性和次生代謝合成等方面優于普通大麥[34-35],這與本研究中青稞抗氧化酶活性高于一般飼料大麥的結論相呼應。
4 結論
本研究通過馬鈴薯—大麥間作試驗,發現間作時分蘗期大麥苗抗氧化酶活性顯著高于單作;間作二棱大麥抗氧化酶活性最高,單作多棱大麥抗氧化酶活性最低;間作時啤酒大麥品種抗氧化酶活性最高,青稞品種次之,飼料大麥品種抗氧化酶活性最低;大麥苗各抗氧化酶活性具有協同作用。綜合考慮,用隸屬函數法對間作大麥苗抗氧化性綜合評價,初步篩選出云啤麥MF20-5、寬穎大麥、MF20-16 等抗氧化綜合評價前10 的品種(系),作為最適宜與馬鈴薯間作且抗氧化性強的云南大麥品種(系)。
