固相自養(yǎng)-異養(yǎng)反硝化脫氮同步去除微污染物
2024-03-28 08:05:14劉彩虹陳子惟劉乾亮重慶大學(xué)環(huán)境與生態(tài)學(xué)院三峽庫(kù)區(qū)生態(tài)環(huán)境教育部重點(diǎn)實(shí)驗(yàn)室重慶400044哈爾濱理工大學(xué)化學(xué)與環(huán)境工程學(xué)院黑龍江哈爾濱50040
中國(guó)環(huán)境科學(xué) 2024年3期
關(guān)鍵詞:體系
曹 陽(yáng),劉彩虹*,陳子惟,劉乾亮,何 強(qiáng)(.重慶大學(xué)環(huán)境與生態(tài)學(xué)院,三峽庫(kù)區(qū)生態(tài)環(huán)境教育部重點(diǎn)實(shí)驗(yàn)室,重慶 400044;.哈爾濱理工大學(xué)化學(xué)與環(huán)境工程學(xué)院,黑龍江 哈爾濱 50040)
目前,排放的污水主要包括市政和工業(yè)污水,不同污水中污染物成分差異較大,種類復(fù)雜,且不同污染物及其轉(zhuǎn)化產(chǎn)物之間還會(huì)產(chǎn)生協(xié)同、拮抗等復(fù)合毒性作用,威脅生態(tài)安全和人類健康.其中,以重金屬鉻和高氯酸鹽為代表的有害物污染問(wèn)題不容忽視[1-2],藥物與個(gè)人護(hù)理品(PPCPs)也在水體環(huán)境中頻繁檢出,常見(jiàn)的如雙酚A(BPA)和萘普生(NPX)在污水中檢出濃度為ng/L~μg/L 級(jí)別[3].在城鎮(zhèn)污水廠處理過(guò)程中,以上微污染物常與NO3--N 共存,需同步去除.污水廠進(jìn)水由于碳氮比(C/N)較低,常依賴于外加碳源實(shí)現(xiàn)出水中氮排放達(dá)標(biāo).為克服傳統(tǒng)液體碳源易過(guò)量、易燃、有毒的弊端,近年來(lái)固相反硝化應(yīng)運(yùn)而生,其中以人工可降解多聚物與天然纖維復(fù)合而成的復(fù)合碳源[4]最具優(yōu)勢(shì),但其在運(yùn)行過(guò)程中仍存在脫氮效率不足等問(wèn)題.近年來(lái),有研究[5]表明鐵(Fe2+)自養(yǎng)反硝化可實(shí)現(xiàn)高效脫氮,若將其與異養(yǎng)反硝化結(jié)合,可進(jìn)一步減輕體系中各反硝化還原酶之間對(duì)碳源電子的競(jìng)爭(zhēng),整體提高反硝化效能.此外,硫自養(yǎng)反硝化可通過(guò)異養(yǎng)反硝化產(chǎn)生的堿和硫自養(yǎng)反硝化產(chǎn)生的酸實(shí)現(xiàn)酸堿互補(bǔ)、降低異養(yǎng)反硝化過(guò)程中污泥的產(chǎn)量、降低自養(yǎng)反硝化過(guò)程中SO42-的產(chǎn)量等途徑增強(qiáng)體系中反硝化速率,同時(shí)降解高價(jià)態(tài)微污染物和PPCPs[6].
目前,大部分研究對(duì)于異養(yǎng)反硝化耦合鐵(FeO)自養(yǎng)反硝化的研究較少,對(duì)于將異養(yǎng)-自養(yǎng)復(fù)合反硝化運(yùn)用于痕量污染物(Cr( Ⅵ)、ClO4-、BPA、NPX)與NO3--N 同步去除的研究亦較少.基于此,本文在聚己內(nèi)酯復(fù)合花生殼(PCL/PS)固體碳源的基礎(chǔ)上,考察其分別耦合單質(zhì)硫(S)、氧化鐵(FeO)和同時(shí)耦合二者所構(gòu)建的復(fù)合碳源對(duì)反硝化體系脫氮效能的強(qiáng)化作用,以及同步去除微污染物(以Cr( Ⅵ)、ClO4-、BPA、NPX 為例)的能力,揭示復(fù)合自養(yǎng)/異養(yǎng)反硝化體系的微生物群落結(jié)構(gòu)、功能種群分布的關(guān)聯(lián)性及微觀作用機(jī)制,旨在為尾水深度脫氮及微污染物同步處理工藝的構(gòu)建提供參考.
1 材料與方法
1.1 固體碳源的制備與表征
花生殼作為富含木質(zhì)素和纖維素的天然材料,具有強(qiáng)吸附性,選作復(fù)合碳源的天然成分(采購(gòu)于中國(guó)重慶本地),同時(shí)選用PCL(分子量~50000Da,中國(guó)一順工業(yè)有限公司)作為聚合物基,添加劑均為分析級(jí)采購(gòu).復(fù)合碳源制備方法如下:花生殼經(jīng)蒸餾水洗滌后,在35℃下脫水,研磨成粉末.采用高速混合機(jī)(50KG-25,安博維機(jī)械有限公司,中國(guó))對(duì)共混后的固體碳源進(jìn)行均質(zhì)處理,通過(guò)直徑為2mm 的雙螺桿擠出機(jī)(20R,南京永杰化工機(jī)械有限公司),按3mm的規(guī)格進(jìn)行最后切割.制備的4 種固體碳源的原料組成及質(zhì)量配比如表1 所示,該比例由前期研究[7]并結(jié)合雙螺桿擠出機(jī)中可添加的天然材料的最大百分比確定.將制備的固體碳源在40℃下干燥15h后加入反應(yīng)器.固體碳源采用 SEM(Gemini 300,ZEISS,德國(guó))對(duì)其表面形貌進(jìn)行表征,并對(duì)固體碳源上的微生物形態(tài)進(jìn)行測(cè)定.固體碳源官能團(tuán)利用傅里葉變換紅外輻射(FTIR)光譜儀(Thermo Scientific Nicolet 6700)進(jìn)行分析,采用KBr 壓片法,波長(zhǎng)范圍4000~400cm-1.
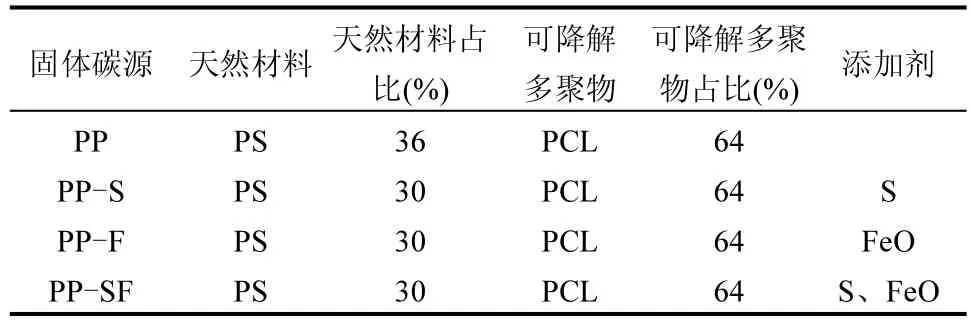
表1 固體碳源組成及質(zhì)量配比Table 1 Solid carbon source composition and mass ratio
1.2 實(shí)驗(yàn)污泥
實(shí)驗(yàn)所用污泥(MLSS=14380mg/L)為重慶雞冠石污水處理廠厭氧池污泥.實(shí)驗(yàn)用水為人工模擬二級(jí)出水,COD、NH4+-N、NO2--N、NO3--N、TN、PO43--P 濃度分別為157.21~212.66,0~0.15,0~0.11,8.42~13.94,10.23~14.71,9.41~11.54mg/L, pH 值為7.5~8.0.其它相關(guān)化學(xué)試劑如BPA(C15H16O2,MW=228.29)、重鉻酸鉀、高氯酸鈉、NPX、1,5-二苯基碳酰二肼、丙酮等均為分析純級(jí)別.所有溶液和儲(chǔ)備液均采用超純水配制.
1.3 實(shí)驗(yàn)方法
實(shí)驗(yàn)共分4 組(A~D,分別以Cr( Ⅵ)、ClO4-、BPA、NPX 為目標(biāo)去除微污染物),考察每組在4 種固體碳源體系下(PP、PP-S、PP-F、PP-SF)對(duì)微污染物的去除情況,并設(shè)置3 組平行. 原水中NO3-N 濃度為5mg/L,Cr( Ⅵ)、ClO4-、BPA、NPX 濃度分別為0.5mg/L,20mg/L,20μg/L 和20μg/L.
以500mL 厭氧瓶為反應(yīng)容器,實(shí)驗(yàn)開(kāi)始前,在每個(gè)厭氧瓶分別加入80g固體碳源和400mL實(shí)驗(yàn)配水,曝氮?dú)?5min,以排出配水和厭氧瓶?jī)?nèi)原有的氧氣,再接種40mL 污泥.所有厭氧瓶瓶口均用丁基橡膠塞加鋁蓋密封,在25℃、40r/min 的溫水搖床中運(yùn)行.根據(jù)相關(guān)研究[8-9],設(shè)置運(yùn)行周期為14d.實(shí)驗(yàn)開(kāi)始后每24h 采樣1 次,測(cè)定NO3-、NO2-、微污染物(Cr( Ⅵ)、ClO4-、BPA、NPX)及TOC 的含量.
1.4 分析儀器與測(cè)試方法
每24h 對(duì)反應(yīng)器的出水取樣并測(cè)試污染物濃度,常規(guī)水質(zhì)指標(biāo)測(cè)試前先經(jīng)0.45μm 微孔的濾膜過(guò)濾,按照《水和廢水監(jiān)測(cè)分析方法》[10]進(jìn)行檢測(cè).Cr( )Ⅵ濃度測(cè)定前水樣用0.22μm 微孔濾膜二次過(guò)濾,依據(jù)《二苯碳酰二肼分光光度顯色法》(GB7467-1987)使用紫外-可見(jiàn)分光光度計(jì)(UV-2600,日本)進(jìn)行檢測(cè).采用Dionex Aquion(AQ)離子色譜進(jìn)行ClO4-的檢測(cè)分析.BPA 和NPX 使用高效液相色譜儀(HPLC)進(jìn)行檢測(cè),色譜柱為CNW Athena C18-WP,5μm, 100A,直徑為4.6mm,長(zhǎng)度為250mm;柱溫40℃;流動(dòng)相比例甲醇:超純水=75:25(V/V),柱溫30℃,流速0.1mL/min,進(jìn)樣量10μL,保留時(shí)間3min,檢測(cè)波長(zhǎng)254nm.
1.5 微生物樣品分析
實(shí)驗(yàn)結(jié)束后,在各組燒瓶的下層固態(tài)混合物中采集生物膜樣本,進(jìn)行微生物分析.將生物膜置于0.1mol/L 磷酸鹽緩沖鹽水(PBS)中,以225r/min 的轉(zhuǎn)速振蕩40min,使附著的生物膜懸浮.然后,將樣品在10000r/min 下離心10min 后收集沉淀,并使用E.Z.N.A.土壤DNA 試劑盒(Omega Biotek,Norcross,GA,United States)進(jìn)行DNA 提取和純化.以27F(5'-AGAGTTTGATCCTGGCTCAG-3') 和 338R(5'-TGCTGCCTCCCGTAGGAGT-3')為引物,對(duì)V1~V2區(qū)域進(jìn)行擴(kuò)增,隨后對(duì)純化后的PCR 產(chǎn)物進(jìn)行16S rDNA 高通量測(cè)序.基于KEGG 數(shù)據(jù)庫(kù)中的16S rDNA數(shù)據(jù),利用PICRUSt2 預(yù)測(cè)與微生物代謝相關(guān)的功能酶/基因,采用R 軟件進(jìn)行Pearson 相關(guān)性分析、IBM SPSS Statistics 26.0 軟件進(jìn)行組內(nèi)和組間方差分析(ANOVA)、CANOCO5 軟件進(jìn)行主成分分析(PCA).
2 結(jié)果與討論
2.1 固體碳源的表征
如圖1 所示,PP 表面呈現(xiàn)粗糙的層狀結(jié)構(gòu),可能是由于花生殼中富含木質(zhì)纖維素,由于微生物附著通常會(huì)隨載體表面粗糙度的增加而增加,PP 的層狀結(jié)構(gòu)將利于微生物的黏附.隨著放大倍數(shù)增加,PP-S表面一些單質(zhì)S 分子聚集形成團(tuán)聚體堆積在PP 的表面及孔隙中,耦合了FeO 的固體碳源表面較為平整、有細(xì)小孔洞分布.二者同時(shí)耦合時(shí),固體碳源表面既有S 粒聚集又有細(xì)小孔洞分布.從EDS 結(jié)果可知,耦合后材料的元素主要有C、O、Fe、S.從表面形貌的結(jié)果來(lái)看,S 已成功耦合在PP-S、PP-SF上,FeO 已成功耦合在PP-F、PP-SF 上,表明成功制備PP-S、PP-F、PP-SF3 種新型改性固體碳源.

圖1 固體碳源表面的SEM 和EDS 表征結(jié)果Fig.1 SEM and EDS characterization results of solid carbon source surface
2.2 固體碳源反硝化及同步去除微污染物效能分析
2.2.1 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系同步去除Cr( Ⅵ)由圖2 可知,前期PP-S 平均脫氮率最高,反硝化性能最好.第3~5d,PP 和PP-S 對(duì)NO3--N 的去除率穩(wěn)定在90%~95%,NO3--N 濃度低至0.29mg/L.但隨著碳源的消耗,NO3--N 濃度逐漸上升,推測(cè)可能是前期異養(yǎng)反硝化作用較強(qiáng),碳源減少,抑制微生物的代謝,迫使微生物逐步進(jìn)入內(nèi)源呼吸階段.而PP 的回升幅度大于PP-S,推測(cè)是單質(zhì)硫作為電子供體可以在系統(tǒng)電子供體相對(duì)不足時(shí)(C/N 下降)通過(guò)硫自養(yǎng)反硝化作用參與脫氮,彌補(bǔ)反應(yīng)器中因碳源消耗造成的電子供體不足,同時(shí)生成H+分擔(dān)部分堿度,保證脫氮效率穩(wěn)定.該結(jié)果表明,單質(zhì)硫作為系統(tǒng)備用電子供體,在碳源不足時(shí)可作為異養(yǎng)反硝化的補(bǔ)充,維持穩(wěn)定的脫氮效率.然而,PP-F 的NO3--N 濃度一直保持較高水平,平均脫氮率僅有17.2%,可能是Fe2+在細(xì)胞外被氧化,生成的鐵垢滯留在細(xì)胞周圍,阻礙養(yǎng)分的吸收和代謝產(chǎn)物的外流,導(dǎo)致細(xì)胞活性下降甚至死亡[11].對(duì)于Cr( Ⅵ),4 組實(shí)驗(yàn)反應(yīng)器均具有較好去除能力,Cr( Ⅵ)平均去除率分別為85%(PP)、82%(PPS)、75%(PP-F)、75%(PP-SF).其中,PP、PP-S、PP-SF中的最高去除率均可達(dá)92%.在整個(gè)實(shí)驗(yàn)周期內(nèi),每個(gè)反應(yīng)器效能的變化趨勢(shì)相似,表明各反應(yīng)器的性能穩(wěn)定且可重復(fù).隨實(shí)驗(yàn)進(jìn)行,PP、PP-S、PP-SF 去除率略有上升最后穩(wěn)定在85%左右,同時(shí),NO3--N 去除率略有下降,推測(cè)后期NO3--N 與Cr( Ⅵ)對(duì)電子存在競(jìng)爭(zhēng)關(guān)系,且Cr( Ⅵ)對(duì)電子供體具有更強(qiáng)的利用能力.

圖2 投加微污染物為Cr( Ⅵ)時(shí)復(fù)合自養(yǎng)-異養(yǎng)反硝化體系對(duì)NO3--N 和Cr( Ⅵ)的去除Fig.2 The removal of NO3--N and Cr( Ⅵ) by the composite autotrophic-heterotrophic de nitrification system when the micropollutant was Cr(VI)
由此可見(jiàn),PP 的反硝化脫氮同步去除Cr( Ⅵ)的能力最優(yōu),PP-S 其次,耦合自養(yǎng)反硝化后并未對(duì)體系內(nèi)微污染物Cr( Ⅵ)的去除起到促進(jìn)作用.但值得指出的是,PP-S 的脫氮速率較高,這主要是由于硫自養(yǎng)反硝化對(duì)反應(yīng)進(jìn)行了電子的補(bǔ)給.
2.2.2 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系同步去除ClO4-由圖3 可知,在ClO4-同時(shí)存在的情況下,4 種固體碳源對(duì)NO3--N 均可在5d 內(nèi)實(shí)現(xiàn)90%的去除率.PP-S、PP-F 和PP-SF 于第3d 最先實(shí)現(xiàn)對(duì)NO3--N 的最大去除,NO3--N 濃度到達(dá)最低點(diǎn),分別為0.67,1.22,0.51mg/L,而PP 則于第5d 降解NO3--N 最低至0.52mg/L,其中以PP-SF 的脫氮速率最快.體系對(duì)ClO4-的去除發(fā)生在NO3--N 被基本去除之后,前期NO3--N 濃度較高時(shí)ClO4-幾乎無(wú)變化,當(dāng)NO3--N 濃度低于1~2mg/L 時(shí),ClO4-才開(kāi)始呈現(xiàn)較明顯的降解,這與Giblin 等[12]的研究結(jié)論一致.ClO4-濃度從第3d開(kāi)始逐漸減少,且在第4~5dNO3--N 濃度降至最低時(shí)ClO4-的降解速率最快,并在第13~14dClO4-濃度降至最低(0.12mg/L),此時(shí)反應(yīng)器對(duì)ClO4-去除率達(dá)到99.4%.這些現(xiàn)象說(shuō)明與ClO4-相比,NO3--N 更易作為電子受體而被還原去除,同時(shí)NO3--N 幾近完全降解時(shí),可促進(jìn)ClO4-的降解,這與謝宇軒[13]的結(jié)論一致.通過(guò)對(duì)比PP 的表現(xiàn),耦合鐵自養(yǎng)反硝化后反應(yīng)器對(duì)高氯酸鹽的去除性能明顯有了極大提升,PP-F、PP-SF 的去除率分別高達(dá)99.4%、96.2%.與此同時(shí),投加了FeO 的反應(yīng)器中溶液變黃,且檢測(cè)到有大量的Fe3+產(chǎn)生,說(shuō)明FeO 為反應(yīng)提供了電子,通過(guò)化學(xué)還原作用主導(dǎo)了高氯酸鹽的降解.根據(jù)已有研究結(jié)果可知,鐵自養(yǎng)反硝化過(guò)程中,Fe(II)的氧化由化學(xué)作用主導(dǎo)[20],Fe(II)作為電子供體協(xié)助去除ClO4-可能的化學(xué)反應(yīng)式為:7Fe2++ClO4-+4H2O→7Fe3++Cl-+8OH-.由于S 在微生物作用下發(fā)生歧化反應(yīng):4S+4H2O→H2SO4+3H2S,產(chǎn)生大量H+,pH 值降低較快,抑制微生物對(duì)高氯酸根的降解[14],因此PP-SF 的去除率略低于PP-F.在后文微生物群落分析中,梭菌屬數(shù)目的增加也證實(shí)此種情況,除FeO 的化學(xué)作用外,生物作用也不容忽視.
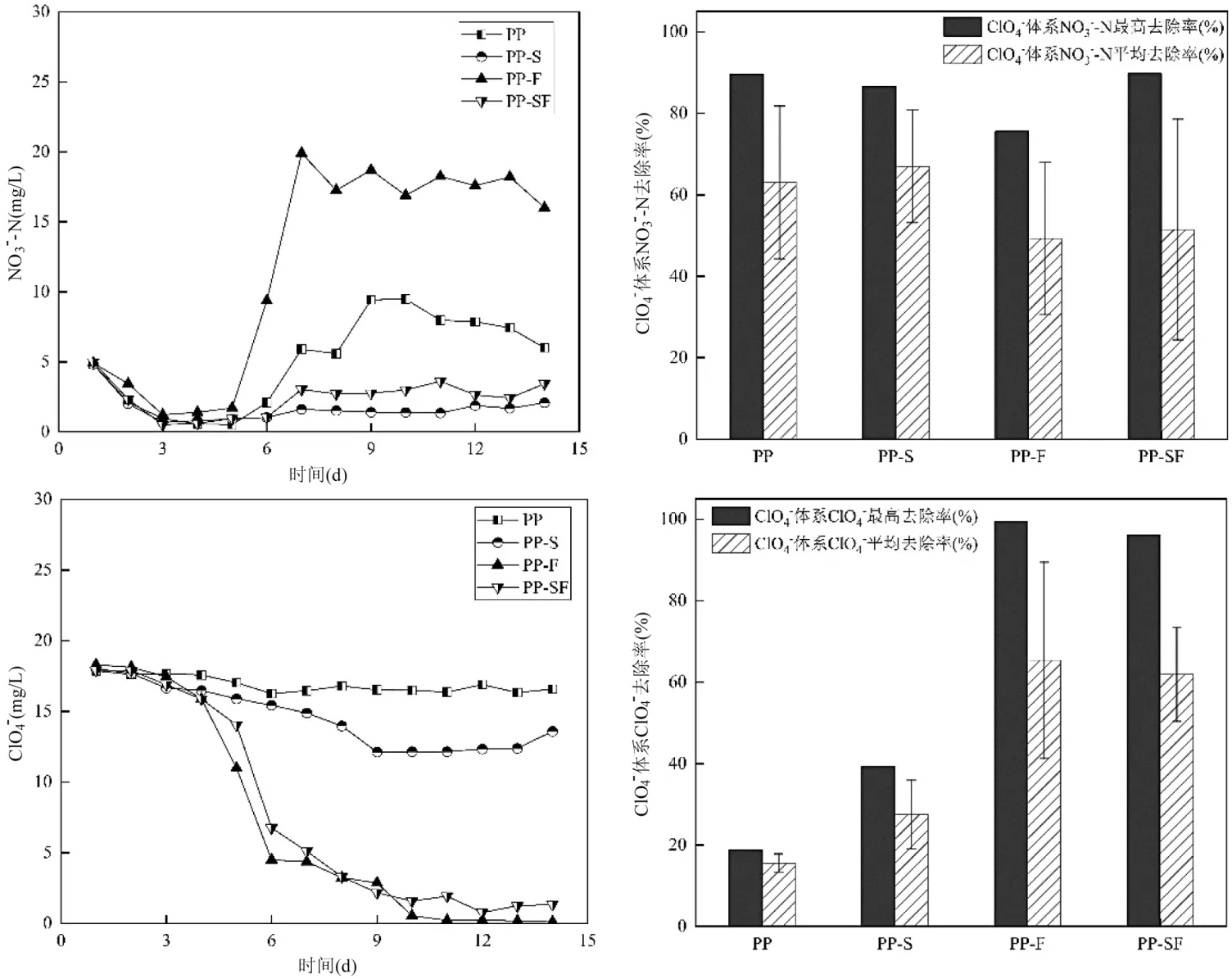
圖3 投加微污染物為ClO4-時(shí)復(fù)合自養(yǎng)-異養(yǎng)反硝化體系對(duì)NO3--N 和的ClO4-去除Fig.3 The removal of NO3--N and ClO4- by the composite autotrophic-heterotrophic denitrification system when the micropollutant was ClO4-
總體評(píng)估,PP-SF 反硝化脫氮同步去除ClO4-的能力最優(yōu),其脫氮表現(xiàn)優(yōu)于含鉻體系,這可能是在高氯酸鹽的存在下,NO3--N 更易被還原去除.耦合了鐵自養(yǎng)反硝化后對(duì)體系內(nèi)微污染物ClO4-的去除有了極大促進(jìn),但PP、PP-S 去除ClO4-的表現(xiàn)不佳,表明該體系中Fe(II)可能在促進(jìn)ClO4-去除中起到了重要的作用.
2.2.3 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系同步去除BPA從圖4 可知,PP 的反硝化性能最好,對(duì)NO3--N 去除率高達(dá)100%,在PP 中添加S 或FeO 后,對(duì)NO3--N在前期有一定去除,后期產(chǎn)生抑制作用,尤其是添加了FeO 后,這可能是鐵對(duì)微生物的抑制作用導(dǎo)致.此外,4 種碳源對(duì)BPA 表現(xiàn)出良好的去除能力,平均去除率皆為99%以上.結(jié)合BPA 和NO3--N 的去除趨勢(shì)分析,可能源于PPCPs 的非生物轉(zhuǎn)化,即在生物脫氮體系中,PPCPs 會(huì)與NH2OH、NO2-、NO3-等氮活性物質(zhì)直接發(fā)生化學(xué)反應(yīng).Yu 等[15]研究發(fā)現(xiàn)向體系內(nèi)加入的NH2OH(5μmol/L)能與體系內(nèi)磺草靈直接反應(yīng),使其氮雜環(huán)上的-NH2發(fā)生羥基化反應(yīng),或氧化為NO2,并將鄰位的-H 取代為-OH.此外,鄭杰蓉等[23]發(fā)現(xiàn)相同時(shí)間內(nèi),投加乙酸鈉后反硝化菌群對(duì)磺胺嘧啶(SDZ)的降解率提高40%左右,表明SDZ作為碳源與乙酸鈉共同被反硝化菌群利用.由此推斷,本實(shí)驗(yàn)體系內(nèi)可能也存在異養(yǎng)菌將PPCPs 作為碳源利用的情況.

圖4 投加微污染物為BPA 時(shí)復(fù)合自養(yǎng)-異養(yǎng)反硝化體系對(duì)NO3--N 和BPA 的去除Fig.4 The removal of NO3--N and BPA by the composite autotrophic-heterotrophic denitrification system when the micropollutant was BPA
通過(guò)總體評(píng)估,PP 的反硝化脫氮同步去除微污染物BPA的能力最優(yōu).各組固體碳源的脫氮同步去除BPA 表現(xiàn)中主要在反硝化脫氮方面有所差距.耦合自養(yǎng)反硝化后對(duì)體系效能無(wú)顯著提升,但值得指出的是耦合硫自養(yǎng)后體系效能發(fā)揮更為穩(wěn)定.
2.2.4 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系同步去除NPX 由圖5 可知,實(shí)驗(yàn)運(yùn)行3~4d 時(shí),各組均實(shí)現(xiàn)了對(duì)NO3--N 的最高去除,去除率分別為80.2%、87.4%、65.6%、95.4%,PP-SF 對(duì)NO3--N 的去除效果最好.各組均在9~10d 時(shí)實(shí)現(xiàn)了對(duì)NPX 的最高去除,去除率分別為100%、99.96%、97.35%、99.95%,這與 Marchlewicz 等[17]的研究結(jié)果相似.Marchlewicz 等從波蘭某化工廠的土壤中分離出了可降解萘普生的菌株,當(dāng)外加碳源對(duì)NPX 進(jìn)行共代謝降解的過(guò)程中,萘普生的降解率顯著提高,可達(dá)90%以上.但NPX 可能對(duì)自養(yǎng)微生物生長(zhǎng)代謝產(chǎn)生一定干擾,因此耦合了自養(yǎng)反硝化后并未對(duì)異養(yǎng)反硝化去除微污染物產(chǎn)生促進(jìn)作用.
通過(guò)綜合評(píng)估,PP-SF 反硝化脫氮同步去除NPX 的能力最優(yōu),PP-S 次之.與BPA 體系類似,各組碳源主要在反硝化脫氮方面有所差距,耦合自養(yǎng)反硝化后體系效能有所提升并更為穩(wěn)定.
2.2.5 固體碳源的優(yōu)選與分析 通過(guò)對(duì)比4 種污染物體系中4 種固體碳源的反硝化脫氮與同步去除微污染物結(jié)果:(1)固體碳源不同,其所擅長(zhǎng)降解的微污染物種類不同.經(jīng)綜合考評(píng),本實(shí)驗(yàn)得到的脫氮與同步去除微污染物性能較好的固體碳源為PP 和PP-SF,其中PP 的脫氮及同步去除Cr(Ⅵ)和BPA 的能力較好,而PP-SF 則在去除ClO4-和NPX 時(shí)表現(xiàn)最好.
(2)在ClO4-的存在下,NO3--N 更易被還原去除,且體系中去除ClO4-的主要貢獻(xiàn)者為鐵自養(yǎng)反硝化反應(yīng).在Cr(Ⅵ)、BPA、NPX 存在時(shí),各組固體碳源反硝化脫氮及同步去除微污染物的綜合性能主要取決于反硝化脫氮方面的差距,而耦合了FeO 的固體碳源表現(xiàn)不太理想.這可能是由于Fe2+在反應(yīng)過(guò)程中具有較強(qiáng)的還原性,極易失電子而被氧化,降低生物的利用率,從而出現(xiàn)鈍化現(xiàn)象而降低脫氮效率[18],且Fe2+在酸性條件下較穩(wěn)定,但大多數(shù)鐵自養(yǎng)反硝化菌的生物活性會(huì)受到抑制.
(3)耦合了S 的固體碳源在脫氮和去除微污染物的表現(xiàn)中都取得了較穩(wěn)定的成績(jī),其優(yōu)勢(shì)在于:產(chǎn)生適當(dāng)酸度,平衡體系pH 值,與有機(jī)碳源一同為微生物提供了穩(wěn)定的生長(zhǎng)環(huán)境;硫自養(yǎng)過(guò)程中,S 元素由0 價(jià)變?yōu)?6 價(jià),大大提高了對(duì)單位底物的供電子能力;在碳源消耗而減少時(shí),硫自養(yǎng)反硝化承擔(dān)部分NO3--N 負(fù)荷,彌補(bǔ)了系統(tǒng)反硝化效率不足的問(wèn)題,同時(shí)也保證出水有機(jī)物含量較低.雖然耦合硫自養(yǎng)技術(shù)后呈現(xiàn)了明顯的優(yōu)勢(shì),但自養(yǎng)菌較異養(yǎng)菌相比生長(zhǎng)繁殖速度慢,容易在底物(本實(shí)驗(yàn)為NO3--N、高價(jià)污染物)競(jìng)爭(zhēng)中處于劣勢(shì),導(dǎo)致耦合體系穩(wěn)定性不佳.因此,應(yīng)深入研究如何控制實(shí)驗(yàn)條件使自養(yǎng)菌、異養(yǎng)菌之間更好地協(xié)作,促進(jìn)脫氮和微污染物去除的效能提升.
2.3 微生物群落結(jié)構(gòu)分析
在實(shí)驗(yàn)結(jié)束時(shí),提取各組底部生物膜進(jìn)行高通量測(cè)序,采用ACE、Chao1、Shannon 和Simpson4個(gè)指標(biāo),分析各反應(yīng)器和接種污泥中的微生物群落多樣性,同時(shí)對(duì)物種不同分類水平上的群落結(jié)構(gòu)進(jìn)行分析.通過(guò)表征微生物多樣性及物種組成,考察各系統(tǒng)反硝化及同步去除微污染物的規(guī)律、機(jī)理及差異性.
2.3.1 Cr( Ⅵ)復(fù)合自養(yǎng)-異養(yǎng)反硝化體系 ACE 和Chao1 指數(shù)表示菌群豐富度且與菌群豐富度呈正相關(guān),Shannon 和Simpson 指數(shù)表示菌群物種多樣性程度,Shannon 指數(shù)與菌群多樣性呈正相關(guān),Simpson 指數(shù)與菌群多樣性呈負(fù)相關(guān).如表2 所示,與對(duì)照組相比,添加了S 或FeO 的體系中擁有更高的微生物豐富度和多樣性,其中以PP-F最為顯著,這可能是鐵對(duì)反硝化有一定抑制作用,使活性污泥整體系統(tǒng)沒(méi)有向以反硝化微生物為優(yōu)勢(shì)的方向發(fā)展,從而未能形成以優(yōu)勢(shì)種群領(lǐng)導(dǎo)的群落結(jié)構(gòu).
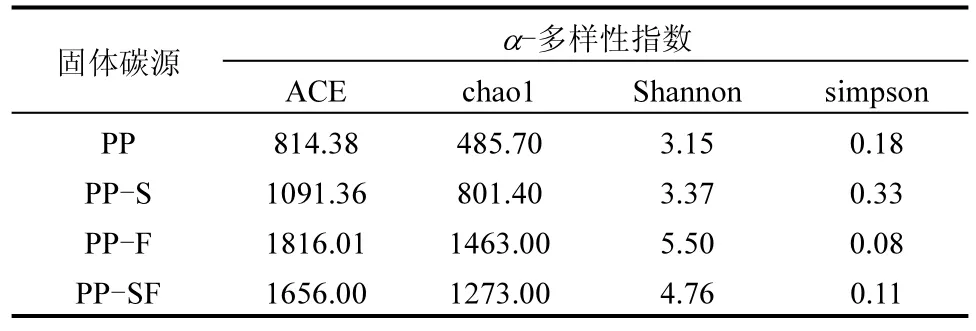
表2 Cr(Ⅵ )復(fù)合自養(yǎng)-異養(yǎng)反硝化體系中的生物多樣性指數(shù)Table 2 Biodiversity index in Cr(VI) complex autotrophicheterotrophic denitrification system
由圖6(a)可知,4 組固體碳源的優(yōu)勢(shì)門均為Proteobacteria(變形菌門)、Firmicutes(厚壁菌門);添加了FeO的PP-F、PP-SF具有獨(dú)特的優(yōu)勢(shì)門Bacteroidetes(擬桿菌門),添加了S 的PP-S、PP-SF 具有獨(dú)特的優(yōu)勢(shì)門Patescibacteria(髕骨菌門).其中Proteobacteria 是固相反硝化系統(tǒng)(如PCL、PBS 等)中的優(yōu)勢(shì)種群,多數(shù)異養(yǎng)和自養(yǎng)反硝化菌均歸屬于Proteobacteria,具有強(qiáng)化脫氮作用[19],Firmicutes 和Bacteroidetes 則通常與有機(jī)物的水解、發(fā)酵及產(chǎn)酸有關(guān),通常作為異養(yǎng)菌和脫氮功能菌參與到污水處理中[20].Patescibacteria 有同步硝化和反硝化功能,是厭氧氨氧化耦合反硝化啟動(dòng)的主要菌門之一[21-22].對(duì)比各組固體碳源對(duì)反硝化和微污染物去除的表現(xiàn)來(lái)看,PP 因具總量最多的反硝化主要菌門Proteobacteria和Firmicutes(二者相對(duì)豐度分別為36.2%、63.2%,共99.4%),在反硝化脫氮中表現(xiàn)最好,而耦合了S/FeO 自養(yǎng)反硝化后雖擁有豐富的微生物多樣性但卻并未對(duì)異養(yǎng)反硝化實(shí)現(xiàn)強(qiáng)化作用,可能與自養(yǎng)反硝化菌與異養(yǎng)反硝化菌存在一定的競(jìng)爭(zhēng)關(guān)系有關(guān).

圖6 Cr(Ⅵ)(a)、ClO4-(b)、BPA(c)、NPX(d)復(fù)合自養(yǎng)-異養(yǎng)反硝化體系微生物在門(左)、屬(右)水平上的種類及分布Fig.6 Species and distribution of microorganisms in Cr(Ⅵ)(a),ClO4-(b),BPA(c),NPX(d) complex autotrophic-heterotrophic denitrification system at phylum(left) and genus(right) levels
由圖6(a)可知,在4 組固體碳源體系內(nèi)均發(fā)現(xiàn)大量異養(yǎng)反硝化菌,如Clostridium_sensu_strito(梭菌屬)、Lactococcus(乳球菌屬)、Novosphingobium(新鞘氨醇桿菌屬);添加了 FeO 的體系內(nèi)還含有Prevotella_9(普雷沃氏菌屬),屬于擬桿菌門,可產(chǎn)生木質(zhì)纖維素降解酶[23],促進(jìn)纖維素降解.對(duì)照組PP中排名前三的優(yōu)勢(shì)屬分別為 Clostridium_sensu_strito_1、Acetobacter(醋菌屬)、Lactococcus,均屬于厚壁菌門,相對(duì)豐度分別為34.6%、26.2%、21.9%,共計(jì)82.7%,遠(yuǎn)高于其他3 組,反硝化性能領(lǐng)先.其中,Lactococcus 代謝產(chǎn)生大量酸質(zhì),能迅速降低反應(yīng)體系內(nèi)的pH 值以防亞硝酸鹽積累,同時(shí)促進(jìn)Cr( Ⅵ)的去除,且能抑制其他有害微生物的繁殖[24].據(jù)研究[25],Lactococcus 還對(duì)難降解有機(jī)物具有降解能力.PP 中還發(fā)現(xiàn)了少量Nitrospira(硝化螺旋菌屬),可參與亞硝酸鹽氧化過(guò)程[26]產(chǎn)生硝酸鹽,并利用硝酸鹽為電子受體進(jìn)行反應(yīng)[27].
2.3.2 ClO4-復(fù)合自養(yǎng)-異養(yǎng)反硝化體系 如表3 所示,PP-S、PP-F、PP-SF 的ACE、Chao1 和Shannon指數(shù)均顯著大于PP,simpson 指數(shù)均小于PP,說(shuō)明這三者具有更高的菌群豐富度和物種多樣性,其中最高者為PP-SF,可能是硫自養(yǎng)反硝化、鐵自養(yǎng)反硝化與異養(yǎng)反硝化耦合后反硝化細(xì)菌不斷增長(zhǎng)和富集的緣故.

表3 ClO4-復(fù)合自養(yǎng)-異養(yǎng)反硝化體系中的生物多樣性指數(shù)Table 3 Biodiversity index in ClO4- complex autotrophicheterotrophic denitrification system
由圖 6(b)可知,4 組固體碳源的優(yōu)勢(shì)門為Proteobacteria(變形菌門)、Firmicutes(厚壁菌門),這與前文中Cr( Ⅵ)復(fù)合自養(yǎng)-異養(yǎng)反硝化體系群落結(jié)果相似.不同的是,添加了S/FeO 的PP-S、PP-F、PP-SF 中的優(yōu)勢(shì)門Bacteroidetes(擬桿菌門)遠(yuǎn)高于PP,而PP 中的優(yōu)勢(shì)門Patescibacteria(髕骨菌門)則多于前三者.有研究表明[28],這些菌群對(duì)生物膜反應(yīng)器中污染物的去除起主要作用,Proteobacteria 和Firmicutes 可在厭氧環(huán)境下通過(guò)分解有機(jī)物來(lái)獲取營(yíng)養(yǎng)物質(zhì),與脫氮作用有關(guān),且大多數(shù)高氯酸鹽還原菌都屬于變形菌Proteobacteria[29];Bacteroidetes 包括3 大類細(xì)菌:Bacteroides 、 Flavobacillus 和Sphingomycetes,可降解花生殼中豐富的纖維素[30].總體而言,4 組碳源中反硝化菌門Proteobacteria、Firmicutes、Bacteroidetes的總相對(duì)豐度為86%~93%,因此反硝化脫氮差異不大.
由圖 6(b) 可知,Clostridium_sensu_strito 和Lactococcus是4組固體碳源體系已知的前兩位優(yōu)勢(shì)屬,均為具有反硝化脫氮、有機(jī)物降解功能的微生物.其中Clostridium(梭菌屬)中包含一些可降解ClO4-的細(xì)菌、硫酸鹽還原菌以及鐵還原菌[31],在中性、中溫條件下對(duì)ClO4-的去除率達(dá)90%以上[29],可在NO3--N濃度較低時(shí)將SO42-和Fe3+作為電子受體[32].在 PP 中發(fā)現(xiàn)了相對(duì)豐度為 5.4%的益生菌Acetobacter(醋菌屬),而其他3 組中則含有相對(duì)豐度最高的自養(yǎng)反硝化菌Prevotella(普雷沃氏菌屬),包括Prevotella_1、Prevotella_7、Prevotella_9.在PP-F中發(fā)現(xiàn)了3.56%的Parabacteroides(副桿狀菌屬),是一種典型厭氧菌,可通過(guò)糖酵解作用產(chǎn)生反硝化電子供體煙酰胺腺嘌呤二核苷酸(NADH)[33].此外,各組碳源體系均存在少量Denitratisoma(脫氮單胞菌屬),為典型異養(yǎng)厭氧反硝化菌,與厭氧氨氧化菌之間存在一定協(xié)同效應(yīng)[34].
2.3.3 BPA 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系 如表4 所示,PP-S、PP-F、PP-SF 的ACE、Chao1 和Shannon指數(shù)均大于PP,simpson 指數(shù)均小于PP,說(shuō)明這三者比對(duì)照組具有更高的菌群豐富度和物種多樣性,其中以PP-F、PP-SF 最為顯著,推測(cè)FeO 可能為影響群落微生物多樣性的最大因素.

表4 BPA 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系中的生物多樣性指數(shù)Table 4 Biodiversity index in BPA complex autotrophicheterotrophic denitrification system
由圖6(c)可知,4 組固體碳源的優(yōu)勢(shì)門均為Firmicutes(厚壁菌門)、Proteobacteria(變形菌門),其中PP-S、PP-F、PP-SF 中的優(yōu)勢(shì)門Bacteroidetes(擬桿菌門)多于PP,這與前文中ClO4-復(fù)合自養(yǎng)-異養(yǎng)反硝化體系的群落結(jié)果相似.不同的是,PP-S 中的優(yōu)勢(shì)門Patescibacteria(髕骨菌門)多于另外三者.總體上看,4 組固體碳源中脫氮功能菌的總相對(duì)豐度均高達(dá)95%以上,但各自脫氮能力有所差異.此外,生物脫氮中混合菌群的協(xié)同作用能實(shí)現(xiàn)PPCPs 較高去除,一方面,異養(yǎng)菌能直接對(duì)PPCPs 實(shí)現(xiàn)共代謝降解[35];另一方面,自養(yǎng)菌可將PPCPs 降解成簡(jiǎn)單產(chǎn)物,進(jìn)一步被異養(yǎng)菌作為碳源利用,實(shí)現(xiàn)完全降解.同時(shí),在復(fù)合反硝化體系中還發(fā)現(xiàn)了與硫自養(yǎng)反硝化有關(guān)的Epsilonbacteraeota(ε-變形菌門),可深度降解廢水中難降解有機(jī)物[36].
由圖6(c)可知,各固體碳源體系內(nèi)優(yōu)勢(shì)菌屬主要有Lactococcus、Novosphingobium、Clostridium_sensu_strito、Prevotella、Enterobacte(腸桿菌屬)等,大多數(shù)為具有異養(yǎng)反硝化脫氮、污染物降解功能的微生物.其中,腸桿菌屬在降解有機(jī)污染物過(guò)程中能釋放生物表面活性劑,提高對(duì)污染物的生物利用率[37].同時(shí),在PP-SF 中出現(xiàn)了2.62%的Pseudomonas(假單胞菌),兼具異養(yǎng)和自養(yǎng)反硝化及聚磷作用,并具有電化學(xué)活性[38].史江紅等[39]分離出的一株香茅醇假單胞菌(Pseudomonas citronellolis)能實(shí)現(xiàn)對(duì)E1、E2 和EE2(均為甾體類雌激素,一種環(huán)境內(nèi)分泌干擾物)的近100%降解,由此推測(cè)該菌屬也可能對(duì)BPA 的降解有相當(dāng)貢獻(xiàn).在PP 體系中,發(fā)現(xiàn)了相對(duì)豐度為3.82%的Acinetobacter(不動(dòng)桿菌屬)和2.41%的Kluyvera(克呂沃爾氏屬),Acinetobacter 能降解芳香族化合物,Kluyvera 是變形桿菌在屬水平上的異養(yǎng)反硝化優(yōu)勢(shì)菌種.研究表明,BPA 會(huì)抑制活性污泥脫氫酶活性,降低微生物的活菌比例[40],結(jié)合前文中各組碳源的反硝化脫氮同步BPA 降解的效能和菌屬豐度差異,推測(cè)BPA 的存在降低了Lactococcus、Novosphingobium、Acinetobacter、Kluyvera 等菌屬豐度,影響系統(tǒng)的脫氮性能.
2.3.4 NPX 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系 如表5 所示,PP-S、PP-F、PP-SF 的ACE、Chao1 和Shannon指數(shù)均大于PP,simpson 指數(shù)均小于PP,說(shuō)明這三者具有更高的菌群豐富度和物種多樣性,其中PP-F 最高,表明FeO 是影響群落微生物多樣性的最大因素,同時(shí)對(duì)比PP-S、PP-SF 的多樣性指數(shù),推測(cè)鐵自養(yǎng)微生物和硫自養(yǎng)微生物在該體系下可能存在一定競(jìng)爭(zhēng)關(guān)系.

表5 NPX 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系中的生物多樣性指數(shù)Table 5 Biodiversity index in NPX complex autotrophicheterotrophic denitrification system
由圖6(d)可知,4 組固體碳源的優(yōu)勢(shì)門均為Firmicutes(厚壁菌門)、Proteobacteria(變形菌門),其中PP-S、PP-F、PP-SF 的優(yōu)勢(shì)門Bacteroidetes(擬桿菌門)多于PP,而PP-S 的優(yōu)勢(shì)門Patescibacteria(髕骨菌門)則多于另外三者,這與前文中BPA 復(fù)合自養(yǎng)-異養(yǎng)反硝化體系的群落結(jié)果一致.此外,在添加了S/FeO 的固體碳源體系中也發(fā)現(xiàn)了一定數(shù)量的Epsilonbacteraeota 和Nitrospirae(硝化螺旋菌門)、Actinobacteria(放線菌門)、Acidobacteria(酸桿菌門)、Chloroflexi(綠彎菌門),均與反硝化作用有關(guān),且PP-F中存在Actinobacteria(放線菌門,能夠參與鐵循環(huán))和Deferribacteres(脫鐵桿菌門,一種能厭氧降解芳烴的鐵還原菌)[41].結(jié)合前文脫氮及同步去除微污染物的研究結(jié)果來(lái)看,PP-SF 因擁有最高豐度的脫氮功能菌而表現(xiàn)突出,其中 Bacteroidetes、Firmicutes、Proteobacteria 的相對(duì)豐度分別為43.74%、39.34%、14.03%,總豐度為97.11%.
由圖6(d)可知,各固體碳源體系內(nèi)的優(yōu)勢(shì)菌屬為Clostridium_sensu_strito,其具有較強(qiáng)的有機(jī)物降解和無(wú)機(jī)物還原能力,易在厭氧環(huán)境中富集.通過(guò)對(duì)比,Lactococcus 和Lactobacillus 在PP 和PP-S 體系中的相對(duì)豐度明顯高于PP-F 和PP-SF,而PP-F 和PP-SF 體系中的Prevotella_9 則遠(yuǎn)多于PP 和PP-S.另外,在 PP-F 中存在相對(duì)豐度為 5.02%的Parabacteroides(副桿狀菌屬),可進(jìn)行糖酵解作用產(chǎn)酸,這與前文ClO4-體系中的結(jié)論一致.此外,PP-S 中發(fā)現(xiàn)有 8.50%的 Prevotella_7.結(jié)合前文研究發(fā)現(xiàn),NPX 的添加雖然降低了PP-SF 體系中的微生物多樣性,但增加了優(yōu)勢(shì)種群Clostridium_sensu_strito和Prevotella的相對(duì)豐度,且NPX本身或其降解產(chǎn)物可能對(duì)微生物有毒害作用,因此PP-SF 更有助于微生物在NPX 的環(huán)境中適應(yīng)并繁殖.
2.3.5 各體系內(nèi)微生物群落結(jié)構(gòu)對(duì)比 微生物群落結(jié)構(gòu)分析表明S 和FeO 的加入增加了固相反硝化體系中微生物的多樣性和豐富度,使微生物群落結(jié)構(gòu)趨于復(fù)雜,增加了Firmicutes、Actinobacteria、Proteobacteria 等與木質(zhì)纖維素降解和自養(yǎng)反硝化相關(guān)菌屬,以及Pseudomonas 等好氧菌屬的相對(duì)豐度,一定程度上提高了反硝化體系中有機(jī)質(zhì)的降解和物質(zhì)轉(zhuǎn)化,但當(dāng)體系中微生物多樣性過(guò)高且各占有一定豐度時(shí),反而不利于優(yōu)勢(shì)菌的形成,進(jìn)而對(duì)體系反硝化性能造成負(fù)面影響. Clostridium_sensu_strito、Lactococcus 和Prevotella 是4 個(gè)固體碳源體系的主要優(yōu)勢(shì)菌屬,影響體系脫氮作用,其總體相對(duì)豐度為PP>PP-SF>PP-S>PP-F,與實(shí)際反硝化脫氮和微污染物的去除表現(xiàn)大致一致,但由于其并非影響生物脫氮的唯一菌種,如 Novosphingobium、Enterobacte、Nitrospira、Kluyvera、Denitratisoma等也會(huì)對(duì)體系性能造成影響,因此PP 和PP-SF 在去除各微污染物時(shí)偶有起伏.同時(shí),體系中微污染物的存在也會(huì)對(duì)微生物的種群豐富度和多樣性產(chǎn)生影響,且在運(yùn)行過(guò)程中,當(dāng)條件受限時(shí),由于一些菌種同時(shí)存在脫氮或者降解同種有機(jī)污染物,體系中反硝化脫氮和微污染物去除可能存在一定競(jìng)爭(zhēng)關(guān)系,加之其去除機(jī)理復(fù)雜,包括自養(yǎng)菌轉(zhuǎn)化、物理化學(xué)轉(zhuǎn)化、異養(yǎng)微生物轉(zhuǎn)化以及各菌群的協(xié)同作用等,還與進(jìn)水組成、處理工藝工況、微生物活性和污染物質(zhì)自身理化特性密切相關(guān),其中哪種機(jī)理占主導(dǎo)作用還需進(jìn)一步深入研究.
3 結(jié)論
3.1 固體碳源對(duì)微污染物降解的選擇性不同.異養(yǎng)反硝化對(duì)于添加了Cr( Ⅵ)、BPA 的污水具有更好的反硝化脫氮和同步去除微污染物性能,如PP 對(duì)NO3--N、Cr( Ⅵ)的去除率分別為 94%、92%,對(duì)NO3--N、BPA 的去除率均可達(dá)99%以上;復(fù)合自養(yǎng)-異養(yǎng)生物反應(yīng)體系可高效脫氮同步去除ClO4-、NPX,其中PP-SF 中的去除率最高,性能最穩(wěn)定,其對(duì)NO3--N、ClO4-的去除率分別為 90%、96%,對(duì)NO3--N、NPX 的去除率分別為96%、99%,而PP-S和PP-F 的反硝化脫氮和同步去除微污染物性能較不穩(wěn)定.
3.2 自養(yǎng)反硝化帶來(lái)更豐富多樣的微生物.硫自養(yǎng)反硝化和鐵自養(yǎng)反硝化的耦合增加了固相反硝化體系中微生物的多樣性和豐度,相關(guān)微生物主要來(lái)自Proteobacteria、Chloroflexi、Deferribacteres、Actinobacteria 等門類,同時(shí)與有機(jī)質(zhì)降解相關(guān)的菌屬及好氧菌屬的數(shù)目有所增加,提高了對(duì)污染物的降解效能.對(duì)于 4 組固體碳源, Clostridium_sensu_strito、Lactococcus 和Prevotella 是決定反硝化脫氮的主要優(yōu)勢(shì)菌屬,同時(shí),體系中微污染物的存在和微生物群落間的相互作用也會(huì)對(duì)實(shí)驗(yàn)結(jié)果產(chǎn)生影響.
猜你喜歡
商品與質(zhì)量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀(jì)智能(數(shù)學(xué)備考)(2020年11期)2021-01-04 00:38:16
中國(guó)外匯(2019年17期)2019-11-16 09:31:14
中國(guó)衛(wèi)生(2015年12期)2015-11-10 05:13:40
現(xiàn)代企業(yè)(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11
