溫度與負荷對Anammox 菌自富集啟動的PDA 的影響
2024-03-28 08:06:14馬雨晴李笑迪北京工業大學城鎮污水深度處理與資源化利用技術國家工程實驗室北京市工程技術研究中心北京100124
中國環境科學 2024年3期
馬雨晴,王 博,李笑迪(北京工業大學,城鎮污水深度處理與資源化利用技術國家工程實驗室,北京市工程技術研究中心,北京 100124)
厭氧氨氧化工藝已經被證明是可持續和節約能源的工藝,具有運行能耗低,污泥產量少,溫室氣體排放少以及脫氮效率高等優點[1-2].伴隨著厭氧氨氧化工藝在高氨氮廢水處理中成功應用,研究者們開始致力于探索處理主流城市污水.由于厭氧氨氧化細菌生長緩慢,世代時間長,所以啟動往往需要較長時間.厭氧氨氧化菌已被證明在不同類型的污泥中自富集,如普通活性污泥、短程硝化污泥、反硝化除磷污泥和厭氧消化污泥等,這就為厭氧氨氧化工藝的啟動提供了新的策略.通過短程反硝化提供亞硝酸鹽實現厭氧氨氧化細菌自富集的方法已經在實驗室規模UASB 中得以證明[3].但是由于厭氧氨氧化細菌還存在對溫度、基質濃度和有機物等環境敏感的特性[4],使得厭氧氨氧化工藝在污水處理廠中表現出來的脫氮性能和豐度并不理想[5-6].
厭氧氨氧化細菌最適宜的生存溫度為 30~40℃[7-9].當溫度超過45℃的時候,厭氧氨氧化細菌的活性受到嚴重抑制[10].而當溫度從33℃下降到15℃時,氮去除速率明顯下降[11].側流污水處理過程通常具有較高的水溫(~30 ℃), 因此更有利于厭氧氨氧化活性的維持.相比而言,主流污水溫度隨季節變化較大(10~25 ℃), 這會對厭氧氨氧化細菌的活性產生較大的影響.溫度的降低對短程反硝化過程中亞硝酸鹽的積累影響不大[12-13].當溫度在18.5~28.6℃變化時,硝酸鹽到亞硝酸鹽的轉化率可以保持在90%,這就說明通過短程反硝化可以為厭氧氨氧化更好地提供亞硝酸鹽[14].基質濃度也會影響厭氧氨氧化反應,亞硝酸鹽濃度過高會抑制厭氧氨氧化細菌的活性[15].因此在高氨氮廢水中,通常采用梯度提高進水濃度的方式使厭氧氨氧化細菌適應生活環境.厭氧氨氧化是無需有機碳源的工藝,而主流污水中含有的易降解有機物使得異養菌進行生長代謝[9].相對于厭氧氨氧化相對緩慢的生長速率而言,異養細菌的生長速率較高,這可能使得厭氧氨氧化細菌處于劣勢地位,脫氮效果變差[16-17].因此自富集出的厭氧氨氧化細菌要想在城市污水處理廠中發揮作用,就要擁有相對穩定性,能抵抗氮負荷、有機負荷變化以及溫度變化的沖擊.
本文通過運行厭氧氨氧化細菌自富集啟動的PDA 系統,探究了溫度下降及負荷沖擊下長期脫氮性能的變化;然后通過16S rRNA 基因測序分析了微生物群落的變化;最后基于宏基因組測序分析研究了微生物種群中基因的表達情況.
1 材料與方法
1.1 試驗污泥與進水
試驗采用經過兩月閑置的自富集的短程反硝化-厭氧氨氧化(PDA)污泥,為絮體-海綿填料污泥,其中海綿填料是是聚氨酯材質的,邊長20mm,可以有效持留污泥.污泥接種后反應器中的混合液懸浮固體濃度(MLSS)約為1500mg/L.
用投加氯化銨(NH4Cl),硝酸鈉(NaNO3)和乙酸鈉的人工配水作為進水.進水包含兩部分:含氨氮(NH4+-N)和有機物的原水以及硝酸鹽(NO3--N)廢水,每個階段的進水濃度如表1 所示.進水氮負荷由3.2gN/(m3·d)提高到 4.0gN/(m3·d),有機負荷由64gCOD/(m3·d)提高到80gCOD/(m3·d).礦物培養基的組成包含 KH2PO4(60mg/L) 、 MgSO4·7H2O(30mg/L)、CaCl2·2H2O(14mg/L)和KHCO3(150mg/L).微量元素溶液 A(g/L)含有 6.37EDTA·2Na 和9.15FeSO4·7H2O,而微量元素溶液 B(g/L)含有19.1EDTA·2Na,0.24CoCl2·6H2O, 0.25CuSO4·5H2O,0.22NaMoO4·2H2O,0.19NiCl2·7H2O,0.014H3BO4,0.4 3ZnSO4·7H2O , 0.99MnCl2·4H2O.

表1 系統的運行模式和特點Table 1 Operational mode and characteristics
1.2 反應器設置和運行
采用上流式厭氧污泥床(UASB)反應器,UASB的工作容積為2.5L,材質為有機玻璃.反應器配備蠕動泵和pH/DO 在線監測儀(德國 WTW340i).
UASB長期實驗持續進行了約115d,分為3個階段.第I 階段(1~20d)為短程反硝化-厭氧氨氧化污泥的恢復階段,第II 階段(20~40d)和第III 階段(40~115d)為提高進水濃度階段,具體進水濃度和水力停留時間(HRT)見表 1.除此之外,在第 III 階段中67~72d 中,為檢測厭氧氨氧化細菌活性,停止投加有機物,以NH4+-N 和亞硝酸鹽(NO2--N)為底物進行了原位活性測試,同時通過饑餓的策略來提高厭氧氨氧化的貢獻,降低其他異養細菌的競爭.整個實驗不控制反應器溫度,在自然降溫條件下進行,反應器的溫度由25℃逐漸下降到19 ℃.

圖1 試驗裝置Fig.1 Schematic diagram of experimental device
1.3 分析方法
NH4+-N、NO2?-N 和NO3?-N,混合液懸浮固體濃度(MLSS)、混合液揮發性懸浮固體濃度(MLVSS)采用標準方法檢測,DO、pH 和溫度通過DO 和pH 探頭檢測.取反應器的進水以及出水水樣,所有水樣經0.45μm 的中速濾紙過濾后進測定.
在第34(第II 階段)、74(第III 階段)和114d(第III 階段)取出反應器中的絮體污泥以及海綿填料樣品共6 個,并將填料上的生物膜用沖洗的方式進行剝離,進行4000r/min 離心凍干處理.通過DNA 試劑盒(FastDNA Spin Kit for Soil)提取凍干污泥中的DNA,采用NanoDrop ND-1000 分光光度計對提取DNA 的純度和濃度進行測定.通過16S rRNA 基因測序技術對反應器內不同階段的微生物群落結構及多樣性進行檢測.使用正向引物338F 和反向引物806R 用于擴增細菌 16S rRNA 基因的V3-V4 區.
進行宏基因組學測序,將提取的6 個DNA 樣品被超聲處理成400bp 的片段,通過接頭連接,用于Illumina 測序和進一步的PCR 擴增.構建文庫后,檢測文庫的插入尺寸,并使用Illumina 平臺進行測序.對得到的原始序列進行優化處理,進行物種和功能上的注釋以及分類.16S rRNA 基因測序和宏基因組學數據在上海美吉生物醫藥科技有限公司在線平臺進行處理分析(Shanghai Majorbio Bio-pharm Technology Co.,Ltd).
2 結果與討論
2.1 提高負荷對PDA 系統脫氮性能的影響
在第I 階段,為恢復其短程反硝化以及厭氧氨氧化性能,采用較低的進水濃度,進水 NH4+-N 和NO3?-N 分別為(10.1±0.5)mg/L 和(10.2±0.1)mg/L(圖2).運行前2d,NH4+-N 濃度基本沒有下降,NO3?-N 下降7.0mg/L 左右,出水中并沒有NO2?- N 積累.這就說明在閑置過程中,短程反硝化和厭氧氨氧化細菌活性都被破壞.但是隨著反應器的運行,在第8~20d,出水中NH4+-N 從8.0mg/L 逐漸下降到2.6mg/L,NO3?-N濃度也逐漸下降到0.6mg/L.第I階段的氮去除效率從44.3%恢復到81%,出水總氮低于5.0mg/L.這就說明短程反硝化-厭氧氨氧化污泥活性恢復,并在20d 時間恢復到閑置前水平.

圖2 UASB 中的NH4+-N、NO2?-N、NO3?-N、TN 和NRE 的變化Fig.2 Variations of NH4+-N、NO2?-N、NO3?-N、TN and NRE in UASB
在第II 階段,提高進水總氮和有機物的濃度.進水NH4+-N 和NO3?-N 分別提高到(24.9±0.5) mg/L和(25.7±0.4) mg/L,同時HRT 提高到12h.提高后可以看到在進水總氮濃度提高的沖擊之下,出水中NH4+-N 和NO2?-N 的濃度都上升到8.0~9.0mg/L,短程反硝化-厭氧氨氧化反應的NRE 又從81%下降到50%.但是隨著運行在HRT 延長條件下,反應器內短程反硝化和厭氧氨氧化細菌逐漸適應濃度升高的水質,脫氮效果有所上升.但是脫氮過程消耗的NO3?-N/NH4+-N 的比率異常低,出水NH4+-N 下降到0.5mg/L 時NO3?-N 還剩余10.0mg/L 左右,這可能是由于空氣滲入導致硝化作用發生.
在第III 階段,再次提高進水氨氮,硝態氮和有機物的濃度,進水總氮為100.0mg/L 左右,同時為了保證溫度降低情況下的脫氮效果延長了HRT.此時PDA 系統性能并沒有因為負荷提高的沖擊而破壞,NRE 保持在相對穩定狀態, 這可能也得益于水力停留時間延長對厭氧氨氧化細菌的持留.而且隨著HRT 的延長,也使得有機物對反應器中的厭氧氨氧化細菌的負面影響降低.在綜合作用下,短程反硝化和厭氧氨氧化過程抗沖擊能力有所提升.
2.2 自然降溫對系統脫氮性能的影響
在第I 階段溫度變化幅度較小,在(25±1)℃范圍內波動,因此短程硝化-厭氧氨氧化性能在較短時間內得到恢復,出水總氮濃度最低為3.7mg/L(圖2).在第II 階段溫度繼續下降由24.7℃到24℃,此時出水濃度波動主要是由于負荷的沖擊,溫度逐漸下降過程中脫氮效果較穩定,出水NRE 在逐漸穩定在77%左右.
第III 階段溫度下降比較明顯,運行10d 內溫度從24℃下降到22.2℃,可能是由于溫度下降的沖擊,在第56~66d 出現了出水硝態氮升高的情況.這可能是由于厭氧氨氧化細菌活性受到溫度影響而下降,與此同時異養細菌占據主導地位.為了加強厭氧氨氧化細菌的效果,對反應器中進行饑餓處理.在第66d 停止投加有機物,進水調整為 NH4+-N 和NO2?-N,濃度分別為(50.0±1.6) mg/L 和(50.5±2.4)mg/L.在溫度逐漸由23.6℃下降到19.4℃的過程中,出水中NH4+-N、NO2?-N 和NO3?-N 分別為(8.7±1.1),(9.3±0.9)和(10.8±0.8) mg/L.其中 NH4+-N 和NO2?-N 分別下降 42.3mg/L 和 41.2mg/L,消耗NO2?-N/NH4+-N 為 0.97,低于理論值.而生成的NO3?-N/NH4+-N 的比例為0.25 則比較接近理論值.根據計算可以得第 III 階段厭氧氨氧化的活性67.1mgN/(gVSS·d).饑餓處理完畢后,恢復進水,出水NH4+-N 和NO3?-N 相比之前有所下降.這可能是由于系統已經適應環境溫度,異養細菌活性也在饑餓條件下得到抑制,進而使得厭氧氨氧化脫氮能力增加,出水的NRE 最高達到89.9%.在77~100d,溫度繼續下降到18.7℃,反應器的出水總氮濃度略有增加,NRE 下降到80%左右.在100d 之后NH4+-N 濃度下降到1.0mg/L以下,NO3?-N濃度在15.0mg/L左右,這有可能是厭氧氨氧化性能提高產生了更多的硝態氮.但是總體上,反應器的脫氮性能較好,出水總氮在14.1mg/L 左右,NRE 恢復到85.9%左右.這就說明了通過自富集啟動的厭氧氨氧化工藝在提高進水負荷和溫度變化方面有一定的抵抗能力.
2.3 微生物群落變化分析
2.3.1 污泥樣品形態 本文是絮體和生物填料混合體系.通過粒度儀測得絮體污泥的平均粒徑為374μm,已經形成較大的顆粒.在顯微鏡下對污泥進行觀察,可以看到視野內分布有較大的顆粒,在污泥顆粒內部夾雜著淺紅色(圖3).對海綿中心部位切片后觀察,可以看到一些淺紅色的污泥分布在海綿的骨架上.由于厭氧氨氧化菌代謝過程中依賴于亞鐵血紅素c,則一般呈現為暗紅色[18],這可能是厭氧氨氧化在填料內部中富集的證據.

圖3 絮體污泥和生物填料的形態結構(第110d)Fig.3 Morphological structure of floc sludge and bio-carriers(the 110th day)
2.3.2 多樣性和豐富度的分析 由表2 可知,根據足夠高的覆蓋率(>0.99),采樣深度足夠且真實.Chao1 和ACE 的結果表明,相比于生物填料,絮體污泥中擁有更高的豐富度.此外,從Shannon、Simpson指數可以看出,隨著反應器的運行,填料中生物群落的多樣性增加,而絮體污泥中的多樣性減少.
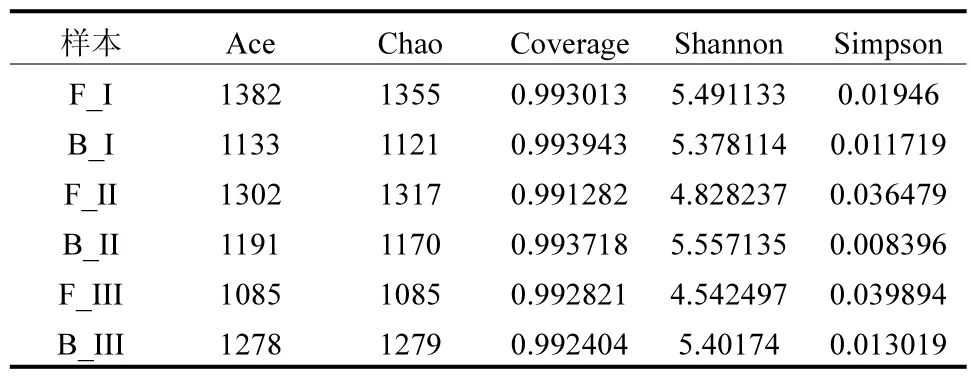
表2 Shannon, Simpson, Ace 和Chao1 指數Table 2 Shannon, Simpson, Ace, and Chao1 indices
2.3.3 微生物群落分析 通過16S rRNA 分析了微生物群落的結構和動態.主要的門為Proteobacteria,Bacteroidetes, Chloroflexi, Planctomycetota(圖4).Proteobacteria 與短程反硝化有關,在絮體污泥中的占比逐漸由34.3%提高到58.7%,在PDA系統中占據絕對優勢[19].其中Thauera 菌屬是將硝酸鹽還原為亞硝酸鹽的主力[20-21],在絮體污泥中的豐度最高可達15.7%,但在生物膜中的豐度僅為0.03%,這是由于相比于生物膜,Thauera 菌更傾向于生長在絮體污泥中,這與其他試驗結果相同[21-22].Chloroflexi 和Bacteroidota 在形成聚集體和生物膜方面發揮了重要作用,在絮體和顆粒污泥中都占一定比例.厭氧氨氧化菌所在的Planctomycetes[23-24],相對豐度從第I階段的7.5%和4.7%下降到第III 階段的3.1%和3.8%,這可能是由于進水濃度或者溫度下降提高對其產生了沖擊. Candidatus Brocadia 作為唯一檢測到的厭氧氨氧化細菌,其在第II 階段生物膜中的相對豐度達到3.7%,這個豐度甚至超過某些接種厭氧氨氧化細菌啟動的反應器中的豐度[25],并且在自富集厭氧氨氧化細菌的反應器中也處于較高水平[26-27].在III 階段可能受到溫度和有機物的影響,豐度有所下降但是其脫氮效果并沒有惡化.重要硝化細菌Nitrosomonas 和Nitrospira 在UASB 中的豐度都低于1%.
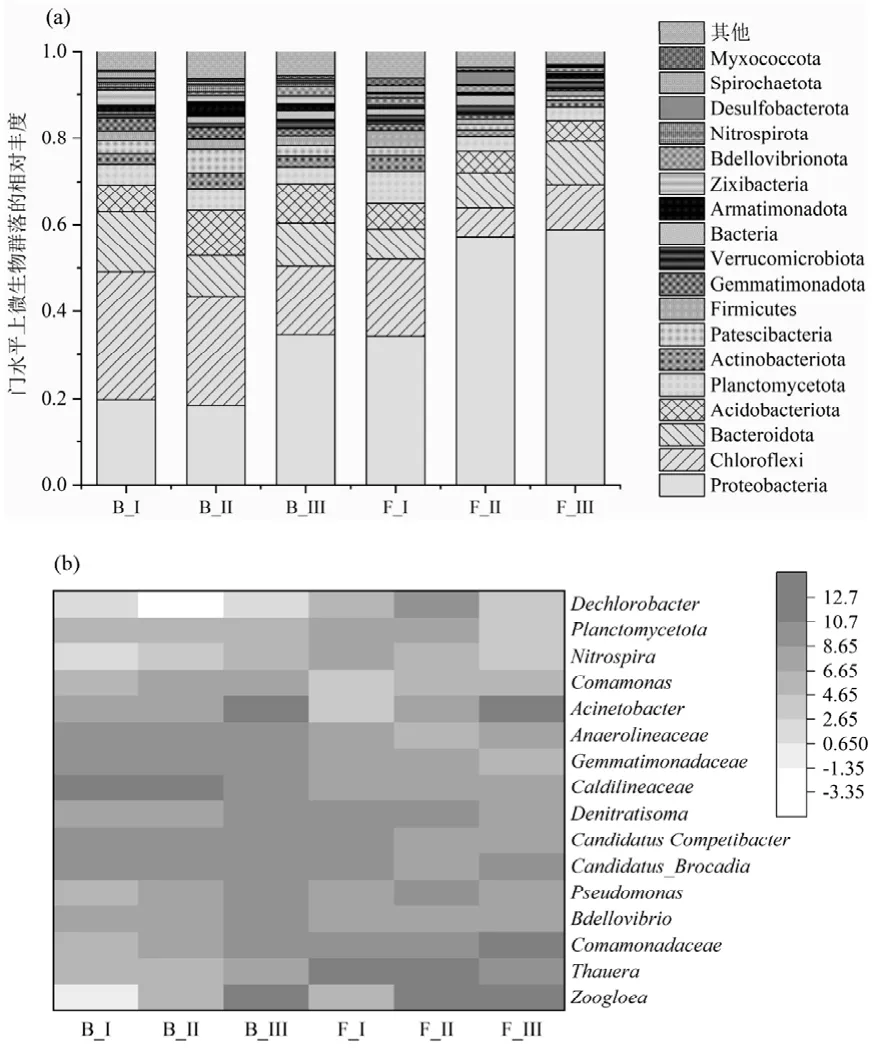
圖4 門和屬水平上的微生物群落分布Fig.4 Microbial community distribution at the phylum and genus levels
2.4 關鍵功能基因分析
圖5 中,與新陳代謝相關的功能基因占比最高,達到了50.0%左右,與氮循環相關的能量代謝功能就被包含在其中.

圖5 系統功能占比以及與氮循環相關基因的表達Fig.5 Proportion of system functions and expression of genes related to nitrogen cycle
在UASB中的生物膜和絮體中都觀察到硝酸鹽轉化為亞硝酸鹽的關鍵基因細胞質硝酸還原酶(Nar)、周質硝酸還原酶(Nap),顯示了硝酸鹽轉化為亞硝酸鹽轉化為厭氧氨氧化富集的潛在途徑. Nar基因根據Reads Number 計算的豐度值之和為83945,說明短程反硝化有關基因表達程度較高.亞硝酸銅還原酶(NirK)和細胞色素cd1 亞硝酸還原酶(NirS)可以將亞硝酸鹽還原為一氧化氮,是發生厭氧氨氧化反應的其中一個重要過程[28].肼脫氫酶(Hdh),肼合酶(Hzs)作為厭氧氨氧化過程中的最重要的兩種酶都可以被檢測到,豐度分別為1118 和12050,這就說明厭氧氨氧化在其中發揮重要作用.NirS 的豐度值為32190,這表明在亞硝酸鹽轉化為一氧化氮的表達程度較高.與反硝化相關的一氧化氮還原酶(NorBC),一氧化二氮還原酶(NosZ)也都可以檢測到,說明反硝化過程在UASB的脫氮過程中也做出了貢獻.與硝化相關的基因AmoABC 都處在非常低的水平,這與微生物群落檢測結果相同,這說明了氨氮主要是通過厭氧氨氧化反應去除的.
2.5 關于自富集PDA 系統穩定性的討論
厭氧氨氧化種泥通常作為接種污泥的一部分,以加速自養脫氮系統的啟動.但它會在一定程度上增加實際應用中的投資和操作的復雜性,而且大量的厭氧氨氧化種泥獲取困難.相比之下,厭氧氨氧化細菌的自富集將是一種更實用、更有效的操作策略.然而,將厭氧氨氧化直接應用于主流工藝通常受到低溫、低氨濃度和高有機物濃度的阻礙.因此自富集的厭氧氨氧化細菌對環境變化的抵抗能力決定了其能否在實際工程中順利應用.
本文中采用厭氧氨氧化菌自富集啟動的PDA系統在主流污水中實現了深度脫氮,總氮去除率達到89.9%,并且發現在溫度以及負荷的變化時自富集啟動的氨氧氨氧化工藝仍然具有較好的穩定性.這可能是因為相比于接種處理高氨氮廢水富集的厭氧氨氧化細菌而言,自富集的厭氧氨氧化污泥更加適應主流污水環境.而且生物填料和絮體相結合的形式有更好的魯棒性,可以更好的抵抗沖擊和低溫的影響.在整個運行過程中亞硝酸鹽的產生充足的同時消耗有機物,消除有機物對厭氧氨氧化的不利影響.此外,整個操作過程中的較長污泥停留時間有效保留了厭氧氨氧化細菌,使得功能微生物Candidatus Brocadia 細菌得到富集.
Wang 等[29]通過16S rRNA 測序發現在水廠的各個單元均有厭氧氨氧化菌的存在,且能夠發揮脫氮作用.Lou 等[30]也發現厭氧氨氧化菌在大規模AAO 氧化溝過程中的富集.因此全規模污水處理廠中檢測到的厭氧氨氧化細菌豐度為將厭氧氨氧化直接整合到主流處理中并實現深度脫氮提供了一定可行性.但是由于厭氧氨氧化細菌自身特性,其在污水處理廠中發揮出的脫氮能力并不能確定.本研究在實驗室規模的反應器中對自富集厭氧氨氧化系統的性能進行的探究取得了較理想的結果.但是對于自富集厭氧氨氧化細菌在污水處理廠中如何發揮深度脫氮的作用仍然需要繼續探究.
3 結論
3.1 在溫度由25℃下降到19℃的條件下,UASB 進水總氮濃度由20mg/L 提高到100mg/L,總氮去除率最高可達到89.9%以上,說明短程反硝化-厭氧氨氧化反應能夠保持一定的穩定性.
3.2 Candidatus Brocadia 和Thaurea 分別為主要的厭氧氨氧化菌和短程反硝化菌.生物填料中厭氧氨氧化細菌相對豐度達到3.7%,絮體污泥中Thaurea相對豐度最高達15.7%.
3.3 與短程反硝化相關的基因Nar,Nap 和與厭氧氨氧化相關的Nir,Hzs 和Hdh 基因表達程度較高.
