調控質粒拷貝數優化生物催化效率的研究進展
2024-04-03 02:14:08李業嚴豪劉夢婷白仲虎
食品與發酵工業 2024年6期
李業,嚴豪,劉夢婷,白仲虎*
1(江南大學 生物工程學院,江蘇 無錫,214122)
2(江南大學,糧食發酵工藝與技術國家工程實驗室,江蘇 無錫,214122)
3(清華大學,深圳國際研究生院生物醫藥與健康工程研究院,廣東 深圳,518000)
質粒是細胞染色體外的雙鏈閉合環狀DNA分子,具有獨立的自主復制能力,廣泛存在于細菌和真菌。質粒本身在生物進化過程中發揮著作用[1]。作為載體,質粒被現代基因工程、合成生物學等領域利用來研究新基因的功能[2-3]。利用質粒作載體攜帶外源基因,通過轉化的方法將其轉入微生物細胞,相比將外源基因整合到宿主基因組來說更加便捷方便。大量研究發現,質粒在微生物宿主細胞內拷貝數與所攜帶基因的表達量關系密切[4-7],優化質粒拷貝數這一方法廣泛在生物化工領域用于優化關鍵基因表達,提高生物催化效率[8]。基于天然質粒構建的優化體系存在諸多局限,近年來已有眾多研究,利用前沿合成生物學手段構建出了更先進的質粒調控優化系統。為此,本文對調控質粒拷貝數優化生物催化效率展開綜述,并探討了通過合成生物學手段進一步優化系統的新研究方向。
1 質粒復制及拷貝數簡介
質粒的復制大致包括復制起始、延伸和終止。復制的起始從復制起點 (origin of replication,replicon) 開始,涉及復雜的蛋白—蛋白、蛋白—DNA相互作用。不同的復制起點序列具有不同的序列結構特征。質粒復制的分子生物學機理已經研究的較為透徹,詳見相關綜述總結[9]。
質粒拷貝數是指每個微生物細胞中質粒相對于染色體的個數[10-11]。對于給定宿主、固定的生長條件,具有特定復制起點序列的質粒通常具有特征拷貝數(或一定拷貝數范圍),這是特定質粒系統通過調節其復制相關元件來實現的。控制系統通過糾正單個細胞中平均拷貝數的偏差,將復制速率維持在穩定狀態,進而決定了質粒拷貝數。相關研究表明,質粒復制起始起作用的負調節因子(抑制劑)參與此調控,主要調控機制包括:a)利用反義RNA (antisense RNA);b)利用轉錄抑制(transcription repression);c)利用復制起點上的迭代子序列(iterons)[12-13]。
質粒拷貝數可以利用基于熒光定量PCR (qPCR)的方法進行絕對定量[14]。拷貝數測量結果表明,可以將質粒大致劃分為高拷貝質粒、低拷貝質粒、單拷貝質粒。a)高拷貝數質粒是指拷貝數較大的質粒,在每個細胞中可達到幾十乃至上千份拷貝,其復制與宿主染色體不同步,僅與質粒本身復制子的控制有關,也稱為松弛型復制控制的質粒[15]。高拷貝質粒一般用作表達目的基因的載體,因為高拷貝的基因通常能導致獲得較高產量的目的基因產物[16];b)低拷貝數質粒是指拷貝數較低的質粒,少到在每個細胞中只有一到幾份拷貝[17],與染色體的復制同步進行,受質粒本身復制子和宿主染色體的雙重調控,也稱為嚴謹型復制控制的質粒,在宿主細胞中表達效率較低,甚至可能導致質粒分配到子細胞失敗[18],因而低拷貝質粒在實際應用中使用較少;c)單拷貝質粒的研究及應用很少。除此之外,宿主的不同、生長條件不同 (培養基成分、溫度),也會影響質粒拷貝數[19-21]。
研究者們對模式原核生物大腸桿菌(Escherichiacoli)和模式真核微生物釀酒酵母 (Saccharomycescerevisiae)的質粒系統研究最為透徹。大腸桿菌及釀酒酵母常見的復制起點、質粒拷貝數以及常用質粒分別如表1及表2所示,其他非模式微生物的質粒系統常以它們為參照開發構建。
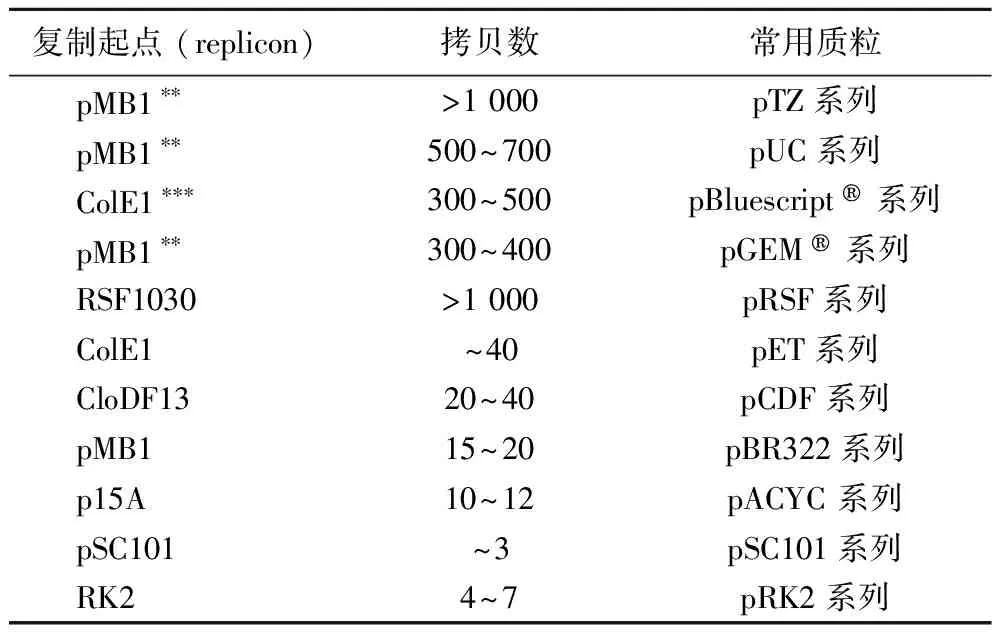
表1 大腸桿菌常見復制起點及拷貝數

表2 酵母常見復制起點及拷貝數
2 天然質粒系統在優化生物催化的應用
大量研究發現,質粒在宿主細胞內拷貝數與所攜帶基因的表達量關系密切[4-7]:一般來說,目的基因的表達量隨重組質粒在宿主細胞中拷貝數的增加而提高,進而提高了目標蛋白的表達量。這種對應關系甚至在質粒拷貝數>300仍存在[9]。不過,當質粒拷貝數超過某個閾值,會嚴重影響系統穩定性,反而使得目標蛋白表達量及目標小分子化合物合成效率降低[22]。上述研究也表明,并非質粒(或目標基因)的拷貝數越高,生物催化的效率也越高。測定不同質粒拷貝數的影響(優化質粒拷貝數),是一種簡單、有效且已經被廣泛應用的優化生物催化效率的策略[8](表3)。
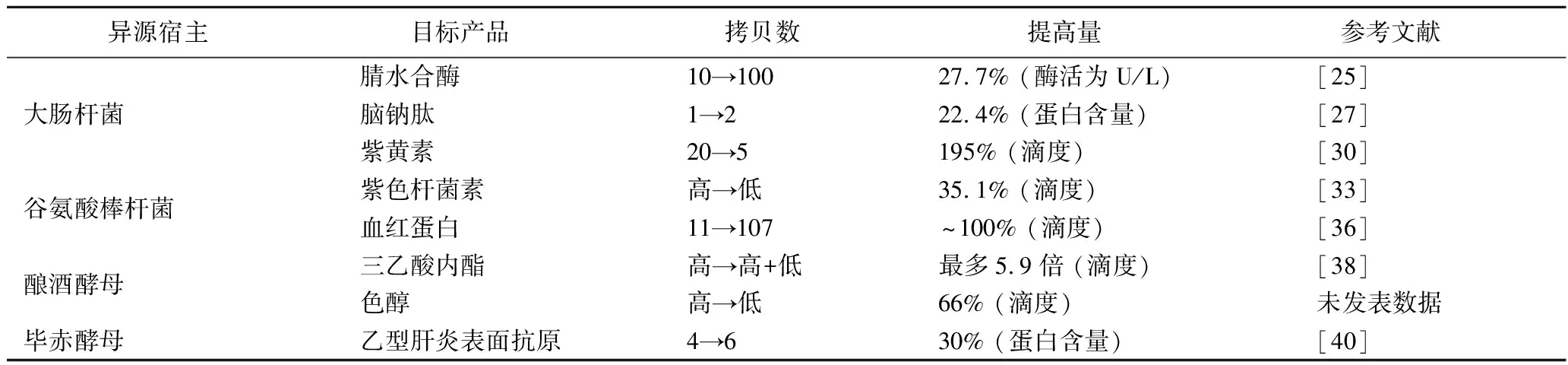
表3 優化天然質粒系統提高生物催化效率
2.1 調控質粒拷貝數原核系統生物催化
2.1.1 在大腸桿菌中調控質粒拷貝數的應用
腈水合酶是一類重要的金屬酶,可以催化腈類化合物水合生成相應的酰胺,在合成酰胺已廣泛應用。但是,目前腈水合酶的數量較少,催化效率低下[23]。為提高腈水合酶的表達量,史悅等[24]在大腸桿菌BL21(DE3)中利用質粒pET28a(+)異源表達諾卡氏菌來源的腈水合酶,比活性可高達450 U/mg,遠高于野生諾卡氏菌,說明了異源多拷貝表達相比野生天然宿主表達具有明顯優勢。王麗燕等[25]在BL21(DE3)中表達Klebsiellaoxytoca來源的腈水合酶,考察了不同拷貝數質粒對腈水合酶表達的影響,發現蛋白濃度(g/L)和粗酶活力(U/L)隨著拷貝數的提高具有增加。使用pCDFDuet或者pETDuet質粒(拷貝數15~20)比使用pACYCDuet質粒(拷貝數10~12) 單位蛋白的粗酶活力(U/g)也隨質粒拷貝數增加而提高,但是使用高拷貝質粒pRSFDuet (拷貝數100~200) 卻對提高酶表達效果并不明顯。因此,優化質粒拷貝數是提高酶異源表達量的簡單可行的辦法。
腦鈉肽(brain natriuretic peptide,BNP)在診斷心力衰竭、調節血壓等方面有很大的醫學作用,尤其是在診斷是否為心力衰竭患者方面,已成為公認的診斷心力衰竭的血漿標志物[26]。異源宿主表達是一種簡便高效合成BNP的有效方法。易俊波等[27]在大腸桿菌DH5α和BL21(DE3)中利用質粒pCW111異源表達了BNP。作者在pCW111上分別克隆單個和兩個BNP的基因表達盒。SDS-PAGE分析發現BNP的表達量隨著目標基因拷貝數增加,菌體總蛋白占比從單拷貝的8.12%增加到兩拷貝的9.94%,這也預示了利用多拷貝質粒進一步增加BNP的潛力。
紫黃素 (violaxanthin) 是一種存在于高等植物和微藻中重要的天然葉黃素,且被證實具有抗氧性、抗癌等生物活性,因而該物質在化妝品、藥品等鄰域具有潛在的應用[28]。紫黃素在微藻中的含量可變的,它會受到外部環境(如光照強度)的影響[29]。將合成紫黃素關鍵合成途徑引入工程菌,通過代謝工程改造構建高產菌,則可以克服上述缺點。DONG等[30]發現嘗試將催化合成紫黃素的玉米黃質環氧化酶(zeaxanthin epoxidase,ZEP)基因克隆到不同拷貝數的質粒上轉入大腸桿菌表達,發現低拷貝質粒(pSC101 ori)效果最好,紫黃素產量相比用高拷貝質粒(pMB1 ori)提高了195%(圖1)。該研究說明了并非質粒拷貝數越高目標產物產量越高,這可能跟高拷貝質粒引起的細胞負擔有關[31]。

A-紫黃質合成途徑;B-使用不同的質粒表達ZEP并應用于類胡蘿卜素的合成
2.1.2 在谷氨酸棒桿菌中調控質粒拷貝數的應用
紫色桿菌素是一種藍紫天然色素,具有抗腫瘤、抗致病菌等多種活性,在食品、印染、化妝品和醫藥行業都有重要的潛在應用價值[32]。產生該色素的天然菌種多為條件致病菌且難于培養,由于紫色桿菌素的前體物質為色氨酸,利用高產氨基酸的異源谷氨酸棒桿菌合成該色素成為一種很好的選擇。盧丹妮等[33]在谷氨酸棒桿菌中構建了紫色桿菌素的合成途徑。研究發現,基于pTET3的低拷貝重組質粒 (p15A復制子,拷貝數~6) 比基于pEC-XK99E的高拷貝重組質粒 (rep/ColE復制子,拷貝數~360)能夠使谷氨酸棒桿菌生產紫色桿菌素的產量提高35.1%。該結果也可能跟前述高拷貝質粒引起的細胞負擔有關。
血紅蛋白廣泛存在于大豆、玉米等植物中,該含有血紅素的蛋白質可被用作顏色增強劑[34]、色素添加劑[35]等。植物血紅蛋白在作為食品添加劑方面具有巨大的價值,且利用微生物可持續生產血紅蛋白是一種有效途徑。WANG等[36]在谷氨酸棒桿菌中實現了植物來源血紅蛋白的高效合成。該研究者們對調控質粒拷貝數相關的起始蛋白編碼基因copA進行了突變,獲得了pXMJ19質粒拷貝數提高的突變體:copAC1033T,copAC1033 del,copAC1032A,copAC1033G,對應測定出的拷貝數分別為107、107、250、318;而表達野生copA時pXMJ19質粒拷貝數僅為11。蛋白表達實驗表明,在一定范圍內蛋白表達量與拷貝數呈正相關,當質粒拷貝數從11增加107,血紅蛋白表達量增加約2倍,但繼續增加質粒拷貝數反而導致產量下降。
2.2 調控質粒拷貝數在真核生物合成生物學中的應用
2.2.1 在釀酒酵母中調控質粒拷貝數的應用
三乙酸內酯 (triacetic acid lactone,TAL) 是一種簡單的聚酮化合物,可在酵母中通過非洲菊III型2-吡喃酮合酶 (2-PS)從乙酰-CoA和丙二酰-CoA合成,被認為是一種可轉化為多種高價值化合物的大宗化學品[37]。YOCUM等[38]通過酵母過氧化物酶體表面展示技術,將TAL合成相關3個酶錨定于細胞質一面,充分利用過氧化酶體流出的乙酰-CoA代謝流合成TAL。研究表明同時利用高拷貝質粒(2 μm)攜帶的自由酶加上低拷貝質粒(CEN/ARS4)攜帶的錨定酶,比僅利用高拷貝自由酶或者低拷貝錨定酶,TAL產量分別提高了3.2倍和5.9倍。
色醇是一種源自L-色氨酸的代謝產物,表現出廣泛的生物活性,并且作為一種關鍵的藥物前體,色醇及其衍生物化學方式轉化為眾多具有藥用價值的化合物。我們的研究表明色醇可以通過釀酒酵母以葡萄糖為碳源合成。分別用基因組整合、低拷貝質粒(CEN/ARS4)或高拷貝質粒(2 μm)表達限速步驟 (TRP2S65R,S76L),發現低拷貝質粒效果最好,基因組整合次之,高拷貝質粒效果最差。這說明色醇的最終產量是基因表達量和細胞負擔兩個因素平衡的結果 (未發表數據)。
2.2.2 在畢赤酵母中調控質粒拷貝數的應用
乙型肝炎表面抗原(HBsAg)是HBV的主要特征蛋白,可以作為感染HBV的診斷指標,在減少HBV感染率且消滅乙肝方面作用很大[39]。乙肝疫苗生產成本高,構建多拷貝質粒并在畢赤酵母中高效表達乙型肝炎表面抗原能減低疫苗成本。實驗表明,乙型肝炎表面抗原在畢赤酵母中的表達量與質粒拷貝數呈正相關,質粒拷貝數從4增加到6,HBsAg在菌體破碎液的含量增加了30%,且拷貝數的提高并未對畢赤酵母的生長產生影響[40]。
3 通過合成生物學手段改造質粒以精確調控拷貝數范圍
綜上研究表明,利用天然復制子序列,可以構建具有不同拷貝數的質粒。構建起一系列不同拷貝數的質粒合集,可以廣泛用于優化關鍵基因表達量,進而提高生物催化效率,提高目標產物產量。前述研究進展也表明,并非質粒拷貝數越高,目標產物也越高,這也凸顯出優化表達系統質粒拷貝數的必要性。
然而,利用天然復制子構建的質粒,拷貝數并不是均勻分散的,還存在著動態調控范圍小等問題。特別是對于真核酵母體系,常用的復制起點只有CEN6/ARS4和2 μm,其拷貝數上限僅為30左右 (表1)。為了克服上述局限,近年來,國內外研究者充分利用合成生物學及分子生物學的前沿技術手段,構建出了質粒拷貝數調控分布更均勻、拷貝數上限更高、可調控范圍更大的系統。這些系統應用于優化生物催化,顯著提高了生物合成的催化效率 (表4)。
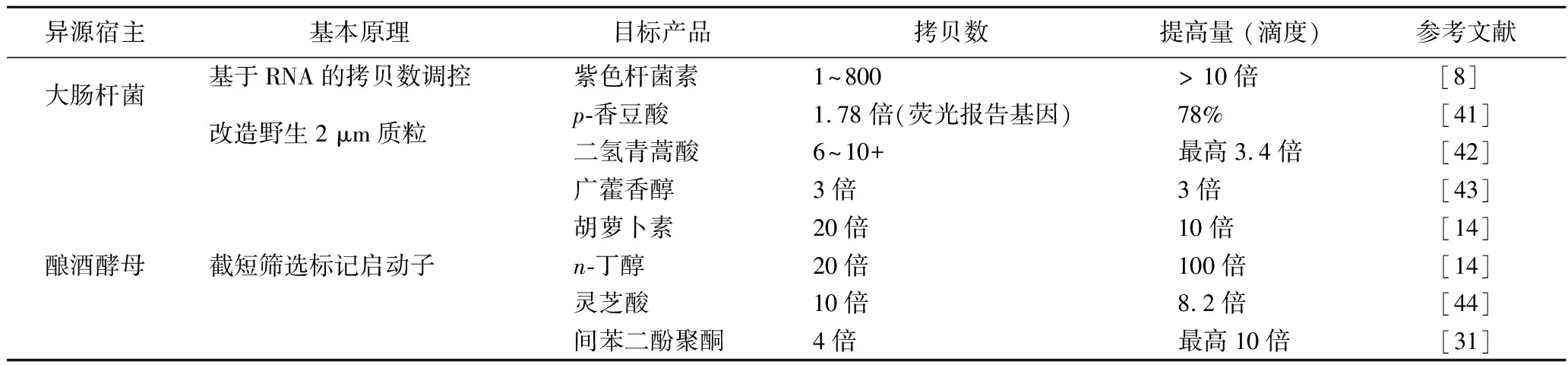
表4 利用合成生物學技術優化質粒拷貝數系統提高生物催化效率
3.1 調控質粒拷貝數在真核生物合成生物學中的應用
大腸桿菌中應用廣泛的、含有ColE1復制起點的質粒,其拷貝數的控制通過兩種RNA實現:a)引發RNA (RNA-p),它以順式方式充當復制起點附近的引物;b)抑制RNA (RNA-i),其轉錄為RNA-p的反義序列,并通過在原點附近雜交之前與引發 RNA 結合而發揮反式作用,RNA-p/RNA-i的比例決定著質粒拷貝數[9]。根據上述機制,ROUCHES等[8]通過調控RNA-p或RNA-i的表達量,構建起了拷貝數可變的2個ColE1質粒系統。在第一個系統中,研究者將Ptet啟動子設置于RNA-p基因前,通過添加無水四環素 (aTc) 控制RNA-p表達量;而RNA-i的表達受ColE1復制子序列自帶啟動子Pi控制。實驗測定在aTc添加濃度為0~100 ng/mL,該系統質粒拷貝數為1.4~50/細胞。在第二個系統中,研究者以ColE1質粒上控制RNA-p的天然啟動子Pp和控制RNA-i的天然啟動子Pi的-35和-10盒子上引入定點和隨機突變,構建出了1 194個RNA-p/RNA-i比例不同(質粒拷貝數不同)基因文庫。序列及拷貝數測定表明質粒拷貝數范圍為1~800/細胞,并且此系統還避免了外源添加aTc。該系統被用于測定質粒拷貝數與細胞負擔的相關性,表明每個質粒會給細胞帶來0.063%的代謝負擔。上述2個系統還被應用于了優化生物合成途徑提高紫色桿菌素產量。第一個系統因為拷貝數變化范圍小、拷貝數較低,紫色桿菌素產量隨aTc添加量的增加而增長;第二個系統由于過高質粒拷貝數帶來的代謝負擔逐漸加重,紫色桿菌素的產量在質粒拷貝數~300時出現峰值。
p15A復制子和ColE1復制子機制類似。LI等根據p15A的復制原理,巧妙設計一套基因回路,構建了受p-香豆酸誘導調控拷貝數的質粒系統。實驗數據表明,該系統能夠動態調控代謝通路中關鍵基因拷貝數,平衡細胞生長與合成目標產物p-香豆酸的代謝通量。與質粒拷貝數不變的靜態系統相比,該動態調控系統p-香豆酸搖瓶產量提高78%,達到1.69 g/L[42](圖2)。
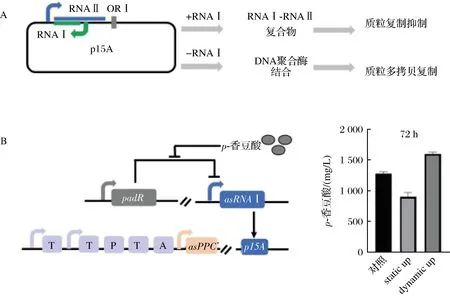
A-RNAI影響質粒復制機理;B-基因拷貝動態控制在p-香豆酸生物合成中的應用
JOSHI等[45]近期開發了原核系統中靈活的、能隨時間變化的動態質粒拷貝數的新系統TULIP (TUnable Ligand Inducible Plasmid)。TULIP的設計基于pSC101復制起點的自動調節機制設計,與前述ColE1復制機制不同。pSC101復制子通過RepA調控蛋白單體-二聚體的平衡使質粒拷貝數控制在3左右。該研究者們利用RepA突變體,設計了新的基因回路,使pSC101固定拷貝數系統變成了受異丙苯酸誘導調控、拷貝數動態調控范圍達到20倍 (3~60拷貝/細胞)的新系統。進一步的表征表明,TULIP在常用的大腸桿菌菌株(NEBStable、DH10B、NEBExpress、BW25113和MG1655) 和生長培養基(M9-葡萄糖、M9-甘油、Lysogeny Broth和Super Optimal Broth)中表現穩定。除了異丙苯酸誘導系統,TULIP也被改造為由其他小分子誘導體系,如N-(3-氧代-己酰基)-L-高絲氨酸內酯[N-(3-oxododecanoyl)-L-homoserine lactone,AHL)],香草酸,異丙基-β-D-硫代半乳糖苷。
3.2 利用合成生物學技術優化真核質粒拷貝數系統
二氫青蒿酸能通過獨立且自發的代謝途徑形成青蒿素[46],且由于青蒿素的醫藥價值不斷被發現,對于青蒿素合成的研究不斷深入。其中利用二氫青蒿酸在釀酒酵母中異源表達,從而進一步合成青蒿素在提高產量方面取得了較好的效果。曾薄軒研究發現,用傳統的2 μm多拷貝質粒表達的關鍵基因ADS、CYP71AV1和DBR2不夠穩定高效,進而影響二氫青蒿酸產量[42]。可以復制2 μm多拷貝質粒的酵母自身還含有內源性的野生2 μm質粒 (wt 2 μm質粒),兩者相比wt 2 μm質粒擁有更高的拷貝數,且在不含任何酵母生長必須基因的前提下,能隨細胞分裂穩定遺傳。因此,在不破壞穩定遺傳所需必須基因的基礎上,作者基于wt 2 μm質粒,設計了利用CRISPR/Cas9基因編輯技術將外源基因插入STB與RAF1基因之間,開發出了pE 2 μm多拷貝質粒系統,可以方便進行外源基因克隆。實驗結果表明,利用pE 2 μm多拷貝質粒表達關鍵基因,相比傳統2 μm質粒系統,在有篩選壓力和無篩選壓力的條件下,二氫青蒿酸產量分別提高到了65%和 3.4倍。類似的,YANG等[47]利用傳統克隆方法在wt 2 μm質粒REP1啟動子和RAF1啟動子之間插入了多克隆位點,方便進行外源基因克隆。研究者們將酪醇合成途徑基因引入該系統,酪醇相比基于傳統2 μm質粒的體系也有提高。
釀酒酵母常用質粒系統具有拷貝數較低、動態變化范圍窄的缺點,很大程度上限制了更多的應用。相關基礎研究發現,酵母質粒拷貝數可以調控其篩選標記基因/蛋白的表達量。CHEN等[43]發現通過將泛素/N-降解決定子標簽 (ubi-tag)融合到篩選標記N可以破壞標記蛋白的穩定性,進而增加載體的拷貝數和同一載體目標蛋白的表達量。這種增加質粒拷貝數的方法使報告蛋白LacZ的活性最高增加100%,將廣藿香醇合成途徑構建進該體系,其產量提高了3倍。
LIAN等[14]隨后系統性利用截短篩選標記基因啟動子的方法來弱化標記蛋白表達量,進而增加質粒拷貝數。他們發現對某些篩選標記 (HIS3,URA3,KanR,HygR),2 μm多拷貝質粒隨篩選標記啟動子截短而增加;但是對另外一些篩選標記 (TRP1,LEU2)則無此相關性,最高拷貝數達到了100 拷貝/細胞。KanR和HygR的系統隨即被用作胡蘿卜素及n-丁醇代謝途徑的優化,產量較野生型最高增加10倍和100倍。類似通過弱化篩選標記基因表達、進而增加篩選壓力來提高質粒拷貝數的方法還用于了重要天然產物靈芝酸[44]。
近期,本課題組將上述拷貝數增加且可調控的方法應用到了雙質粒表達真菌I型聚酮合酶的體系中,用于優化產量極低的間苯二酚類聚酮,因為該類化合物極具藥用價值[32]。通過組合優化,我們將新發現的化合物產量提高最高10倍(圖3)。這加速了此類微量產物的分離純化和結構鑒定,進而促進了結構新穎的非天然真菌聚酮的合成以及加深了對I型真菌聚酮合酶催化機制的理解。

A-組合拷貝數工程優化非天然產物合成;B-通過組合調節還原性聚酮合酶以及非還原性聚酮合酶的質粒拷貝數來優化生產非天然產物
同時有實驗證明,在截短啟動子前提下,不同濃度的抗生素也會影響質粒的拷貝數,且啟動子截短程度較高的對抗生素濃度變化更加敏感[14]。結合啟動子截短工程和抗生素的不同濃度,可以構建拷貝數變化梯度更大的質粒系統。
4 總結與展望
綜上,天然復制子、拷貝數不同的質粒,已經廣泛用于優化關鍵基因表達量,進而提高生物催化效率。然而,天然復制子系統也具有拷貝數分布區間不均勻、動態調控范圍小等局限,這些問題對于真核酵母體系尤為突出。近年來,為了克服上述局限,國內外研究者充分利用合成生物學及分子生物學的前沿技術手段,設計并構建出了質粒拷貝數調控分布更均勻、拷貝數上限更高、可調控范圍更大的系統。這些系統應用于優化生物催化,顯著提高了生物合成的催化效率。但總體而言,目前相關研究不是很多,尤其在真核生物合成生物學領域,這需要對質粒復制機制更深入研究和理解。可以預見,上述領域的進一步進展,可以為合成生物學提供更多解決方案。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
工業設計(2022年8期)2022-09-09 07:43:20
房地產導刊(2022年5期)2022-06-01 06:20:14
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
建材發展導向(2021年12期)2021-07-22 08:06:48
建材發展導向(2021年7期)2021-07-16 07:07:52
中學生數理化(高中版.高二數學)(2021年12期)2021-04-26 07:43:48
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
