毛烏素沙地鹽池地區固定沙丘主要固沙灌木的空間分布格局與空間關聯性研究
2024-04-28 05:50:34牛程旭張定海張志山王艷莉
生態科學 2024年1期
牛程旭, 張定海,*, 張志山, 王艷莉
毛烏素沙地鹽池地區固定沙丘主要固沙灌木的空間分布格局與空間關聯性研究
牛程旭1, 張定海1,*, 張志山2, 王艷莉3
1. 甘肅農業大學理學院數量生物研究所, 蘭州 730070 2. 中國科學院西北生態環境資源研究院, 沙坡頭沙漠研究試驗站, 蘭州 730000 3. 甘肅農業大學林學院, 蘭州 730070
種群的空間分布格局與關聯性的研究對預測種群的演替趨勢和植被重建具有重要意義。基于毛烏素沙地固定沙丘上固沙灌木的地理位置和生長發育階段數據, 采用單變量、雙變量成對相關函數和Monte-Carlo擬合檢驗方法, 對油蒿()、楊柴()和檸條()三種優勢種群進行空間分布格局和關聯性分析。結果表明: (1)三個種群在小尺度下呈現聚集分布, 大尺度下呈現隨機或均勻分布; 楊柴和檸條種群在研究尺度范圍內主要表現為聚集分布。(2)種間關系表明: 油蒿和楊柴在小于5 m尺度下負相關, 其他尺度下基本不具有相關性。(3)種群不同生長發育階段之間的種內關聯關系表明: 在小尺度下, 3個種群的種內關系基本呈正相關; 在大尺度下, 油蒿種內關系呈負相關, 楊柴和檸條的種內關系不存在相關性。
毛烏素沙地; 固沙灌木; 點格局分析; 分布格局; 空間關聯性
0 前言
自然界中生物個體、種群和群落的生存都需要一定的空間, 不同的生物個體生存所需的空間大小存在一定差異。種群是由個體組成, 個體在空間中的位置或布局, 即為種群的空間分布格局。種群的空間關聯性是指不同種群在空間上的相互聯結性, 同時也是種群間相互關系的一種表現形式[1–2]。分析種群的空間分布格局與空間關聯性對于理解種內和種間的競爭關系、依賴關系、外界環境對種群的干擾、種群的生物學特性以及種群和環境的關系具有十分重要的意義[3–5]。
種群空間分布的類型主要分為均勻分布、隨機分布和聚集分布。種群的空間分布格局與空間尺度存在密切的關系, 種群可能在較小的空間尺度下為聚集分布, 但在較大的空間尺度下卻有可能呈現出隨機分布或均勻分布。種群空間關聯性也有3種情形, 空間正相關、空間負相關和空間無相關性[6]。種群空間分布格局與空間關聯性關系密切, 聚集分布與空間正相關體現出種群內部互惠依賴的生態關系, 均勻分布與空間負相關表明種群內部存在競爭的生態關系, 隨機分布與空間無相關性則反映出種群內部的生態關系不明確[7–8]。
對種群空間分布格局傳統的研究方法有: 方差均值比例法[9]、以密度為基礎的樣方法和以距離為基礎的無樣方法[10]、頻次檢驗法、分布型指數法和最近鄰分析法等[11], 其中以最近鄰分析法最為流行。但是, 最近鄰分析法只能檢驗個體分布的隨機性, 不能反映種群空間分布格局隨著空間尺度變化的情況[12]。總體上, 空間格局傳統的分析方法常常限定在特定的空間尺度上進行, 忽略了種群空間分布格局與空間尺度之間的依賴性, 不能全面準確地闡釋種群空間分布格局隨尺度動態變化的情況[13]。目前, 為突破傳統方法的局限性, 已經出現了許多針對不同空間尺度的空間格局研究方法, 如大尺度上的趨勢面分析法, 中等尺度上的雙向軌跡方差法, 小尺度上的種毗連法等[14]。
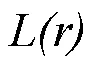
1 材料與方法
1.1 研究區概況
毛烏素沙地是中國四大沙地之一, 研究區位于寧夏鹽池縣毛烏素沙地生態系統國家定位觀測研究站(37°27′N, 107°13′E), 平均海拔1550 m, 年均降水量約為291 mm, 80%的降水主要集中在6—9月, 且多為暴雨集中降落, 固沙植被的降水利用率較低[19, 20], 年均溫度8.34 ℃[21]。研究區的地形主要是高低起伏的丘陵、灘地、谷地、臺地、梁地等, 大部分固定、半固定和流動沙丘主要覆蓋在灘地和臺地上。主要固沙灌木種群為油蒿(, 縮寫為AO)、楊柴(, 縮寫為HM)和檸條(, 縮寫為CK)等。草本有賴草()、白草()、針茅草()等[22]。
1.2 樣地調查
2014年6月沿地形在研究區(37°27′N, 107°13′E)的固定沙丘上設置調查樣地, 調查樣地面積為40 m×128 m, 樣地包含迎風坡、背風坡、丘頂和丘底 4 種沙丘地貌類型。將調查樣地劃分成4 m×4 m的正方形樣方, 調查樣方內所有的固沙灌木, 調查指標包括固沙灌木的類型、生長發育階段、冠幅、株高、固沙灌木所處位置的經緯度和海拔以及樣地的土壤水分和表層土壤養分含量。其中, 固沙灌木的位置利用GPS-RTK定位儀(南京鼎吉測繪儀器有限公司, S86T)測量。
1.3 研究方法
1.3.1 種群空間分布格局分析
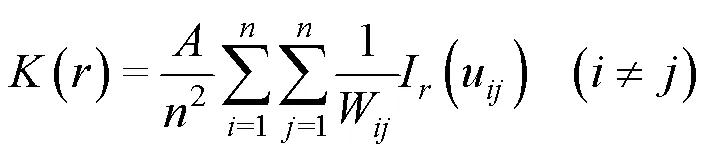
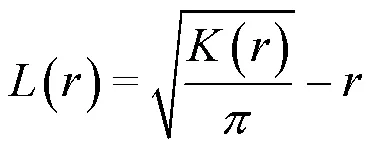
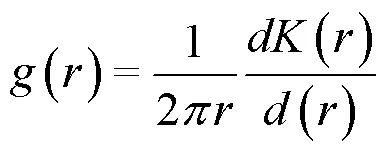
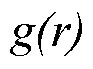
1.3.2 種群空間關聯性分析
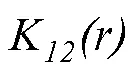
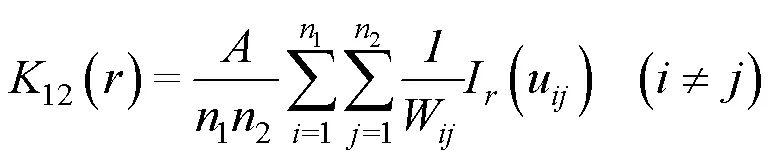
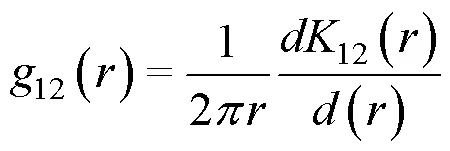
2 結果與分析
2.1 種群的結構組成
研究區固定沙丘調查樣地中廣泛分布著3個固沙灌木種群, 3個種群共有固沙灌木4098株。其中, 油蒿、楊柴和檸條分別有3682株、369株和48株, 占比分別為89.8%、9%和1.2%。將個體的不同生長發育階段分為4種類型, 分別記為幼株(簡記為Y)、營養株(簡記為N)、生殖株(簡記為R)和死株(簡記為D)。3個種群不同生長發育階段的個體數量的占種群數量的比例如圖1所示, 不同生殖階段個體的比例關系均表現出相同的種群結構, 即3種群中營養株, 生殖株、幼株和死株的占比依次減少, 營養株和生殖株的總和占到整個種群的90%以上, 但楊柴的生殖株在種群中的占比較少。同時, 需要指出的是, 樣地內檸條種群中幼株僅有2株, 沒有發現死株, 因此, 在后續的分析中沒有分析檸條種群中幼株和死株的空間格局和空間關聯性。
2.2 種群的空間分布格局
2.3 種群的空間關聯性
2.3.1 不同種群的種間關聯性
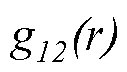
2.3.2 種內不同發育階段的空間關聯性
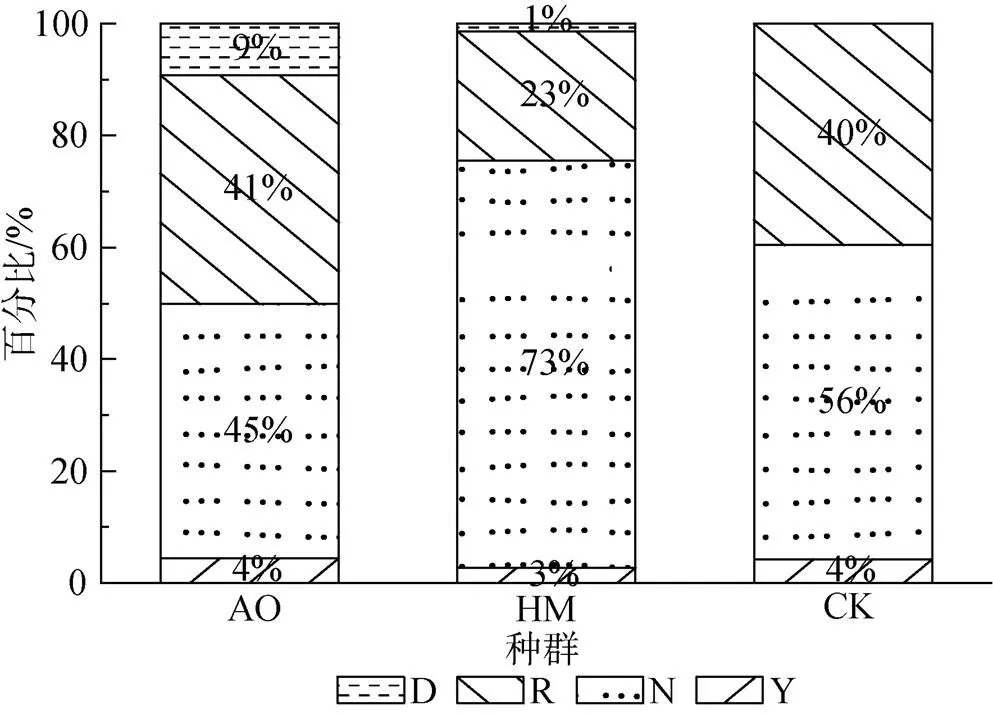
圖1 毛烏素固定沙丘種群的結構組成
Figure 1 Structure composition of fixed dunes population in Mu Us sandy land
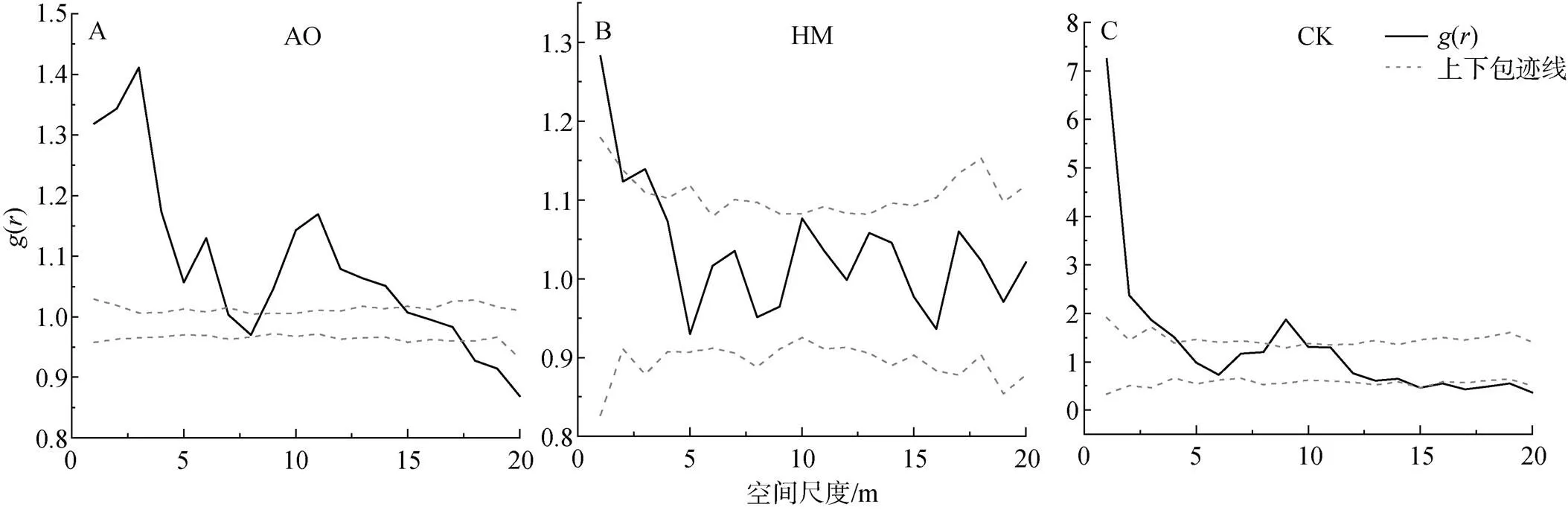
圖2 基于完全隨機模型(CSR)的種群的空間分布格局
Figure 2 Spatial distribution pattern of population based on completely random model
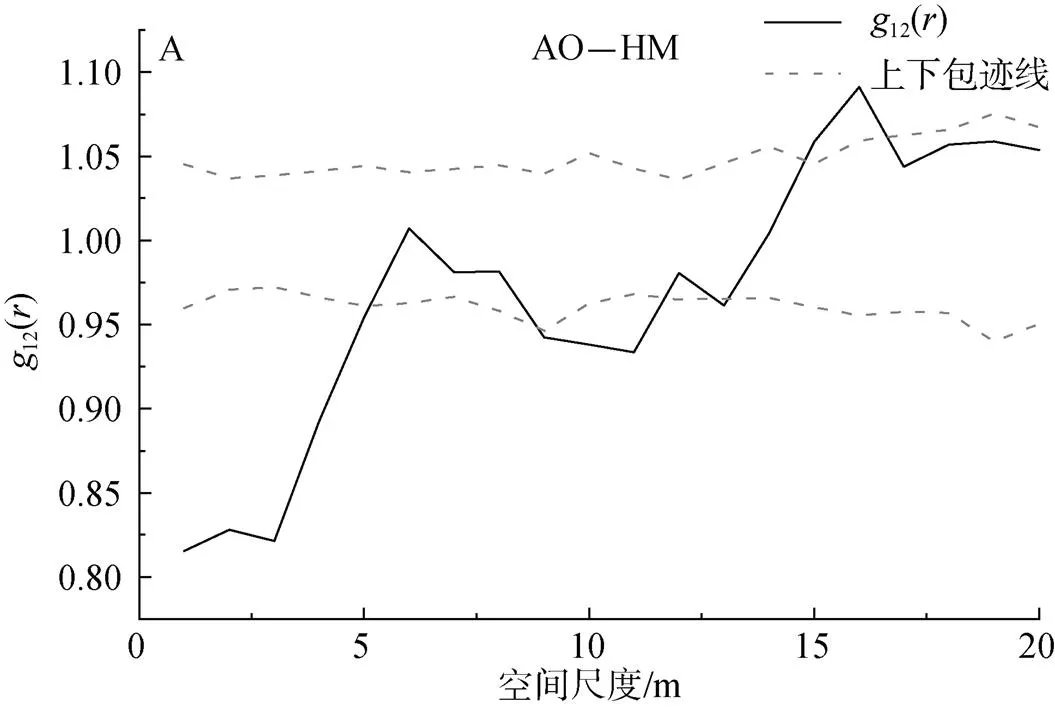
圖3 種間的空間關聯性
Figure 3 Interspecies spatial association
對不同生長發育階段的檸條種群而言(圖5D), 在小尺度下呈現正相關, 大尺度上呈現無關聯或負關聯。具體而言, 檸條種群中營養株和生殖株在小于4 m的尺度范圍內呈現正相關, 在4—18 m的尺度范圍內不存在相關性, 當尺度進一步增大, 大于18 m時, 兩者則表現為負相關。
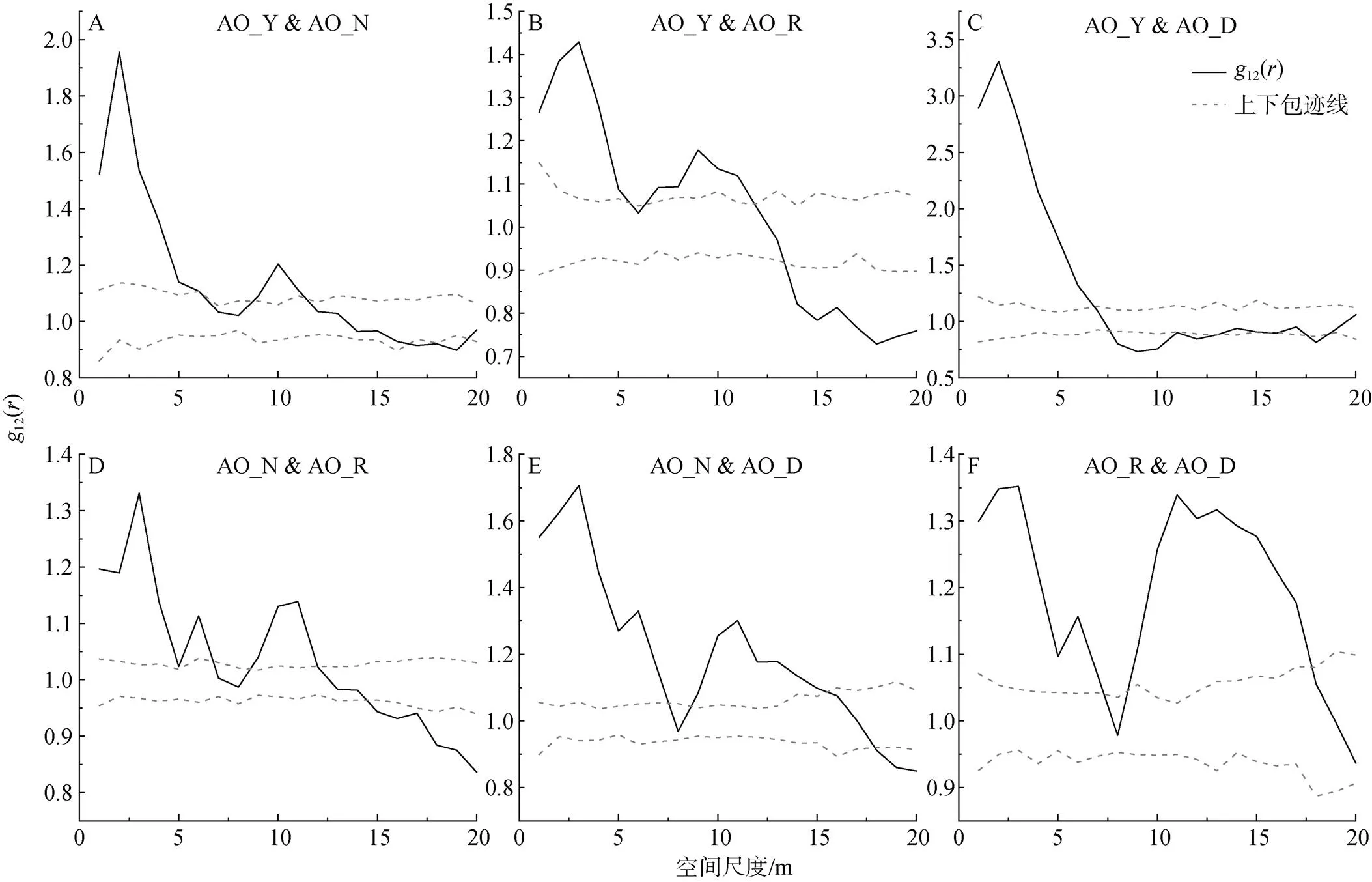
圖4 油蒿不同生長發育階段個體之間的關聯性
Figure 4 Correlation between individuals at different growth stages of AO
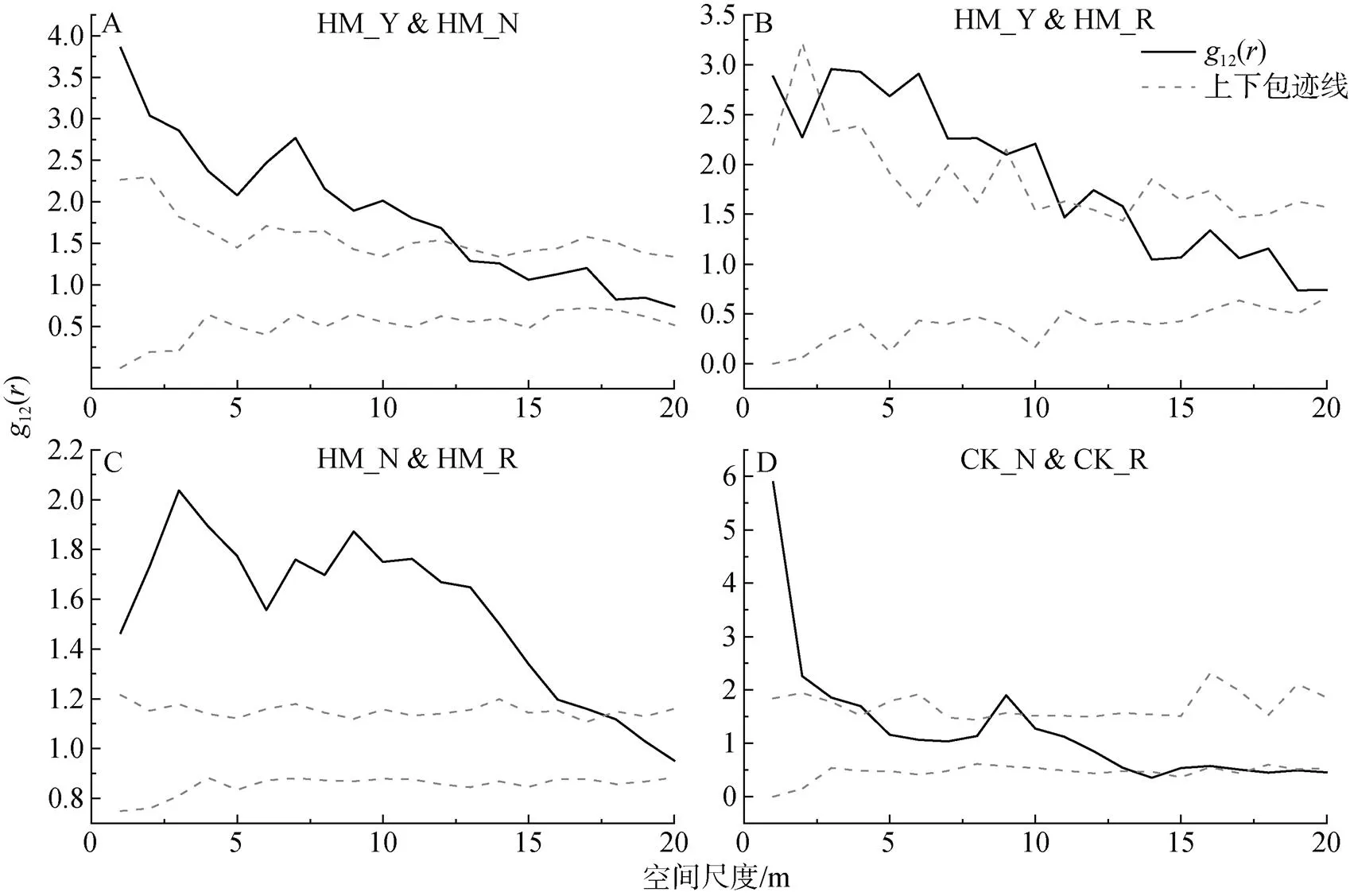
圖5 楊柴和檸條不同生長發育階段個體之間的關聯性
Figure 5 Correlation between individuals at different growth stages of HM and CK
3 討論
3.1 空間尺度與空間格局
種群的空間分布格局通常由種群的特性、生存環境、種內種間的相互關系所決定[24]。不同種群的聚集強度和空間分布情況的變化是種群為適應生態環境, 從而能夠更好地生存的一種策略[25]。油蒿、楊柴和檸條種群的空間分布格局與空間尺度大小存在緊密的聯系。三個種群在研究尺度范圍內多表現為聚集分布, 隨著尺度增大, 聚集強度逐漸降低, 最后趨近于隨機分布或均勻分布。這與許多種群的空間格局的研究結果一致, 種群通常在小尺度下呈現出聚集分布, 在大尺度下多為隨機或均勻分布[6, 7, 16]。產生這種現象, 可能主要與種群的生物學特性和環境因素兩個方面有關[15]。一方面, 油蒿的繁殖方式主要以種子繁殖為主, 但種子繁殖會一定程度下導致播種的成活率較低, 因此在種子擴散過程中, 由于傳播距離的限制和風力風速的影響, 使油蒿個體在20 m尺度下存在密切的關聯關系, 呈現出聚集的分布格局[7, 26–27]。另一方面, 在小尺度下同一個種群對于環境的偏好和資源(水分、土壤、地形等)的需求基本相似, 種群會利用聚集的分布方式產生群體效應, 從而能夠有效抵抗其他物種的競爭和入侵, 抵御惡劣生態環境的影響, 提高種群存活率, 維持種群穩定性[13, 28]。
隨著種群不斷發展, 種群對生存資源的需求不斷增加, 特別是干旱生態系統中, 種群受到水分的脅迫, 其對資源利用的方式發生變化, 從而導致種內和種間競爭加劇, 處于競爭劣勢的個體死亡, 種群密度降低, 個體間距逐漸增加, 最終在大尺度下種群逐漸趨向于隨機或均勻分布[13]。本研究中, 油蒿種內不同發育階段個體之間在尺度增大時普遍出現負相關的排斥作用, 導致幼年株發育受阻, 一定程度證實了種內競爭加劇的現象存在。
楊柴和檸條在研究尺度范圍內主要呈現出的是隨機分布, 這可能是由于楊柴和檸條的根系較深, 主要利用深層土壤水分, 受環境因素和資源競爭的影響較小, 從而主要表現為隨機分布或均勻分布[29]。同時, 楊柴屬于克隆生長的植物, 其具有合軸型分枝類型和游擊型基株構型, 在小尺度上也會呈現聚集分布的特性[30]。
3.2 空間尺度與種間關聯
種群間的相關關系, 通常是在生物學特性、種內種間的相互作用(競爭、依賴等)、生存環境等多種因素的共同作用下, 種群為適應生態環境并合理利用資源占據生態位所產生的結果[31]。在不同空間尺度下, 分析不同種群個體之間的關聯性, 對于研究種群間相互作用機制和群落動態具有重要的意義。
油蒿和楊柴是在較小尺度下負相關性較強, 隨著尺度增大, 兩者主要表現為沒有明顯的關聯關系。不同物種間的相關關系表現為負相關性, 可能有以下兩個方面的原因: 一方面, 不同種群對環境的適應能力不同, 生態位重疊程度較小, 對于生境內資源的利用方面存在差異; 另一方面, 單位面積內資源有限, 不同種群間的對于生存資源(水分、土壤養分等)的競爭需求相同, 在數量上占絕對優勢的種群對其他劣勢種群的競爭排斥作用增強[31, 33]。同時油蒿耐干旱性較強, 在有性繁殖與無性繁殖方面均具有良好的生物學特性, 這使油蒿在相對較差的氣候條件下, 仍然成為種間競爭的優勝者, 逐漸演替為較為穩定的種群[34]。在小尺度下, 這種較為激烈的競爭排斥作用導致油蒿與楊柴之間呈現出負相關關系, 但隨著尺度增大, 油蒿與楊柴均逐漸呈現出隨機或均勻分布, 種群的空間分離格局降低了種群的相遇幾率, 同時生存所需的空間和資源競爭減弱, 因此油蒿與楊柴種群最終呈現出無關聯關系的穩定狀態。
油蒿之所以能在生態系統中占據一定優勢地位,在研究區固定沙丘上與檸條等灌木形成較大數量差異, 主要歸因于以下幾個方面: 一是油蒿的種子產量、種子庫和幼苗密度顯著地高于檸條等灌木[27]; 二是固沙植被演替過程中所形成的生物土壤結皮具有較高的持水能力, 會阻止降水對深層土壤的補給, 導致根深系灌木數量減少直至退出生態系統, 鞏固了油蒿的地位; 同時生物土壤結皮會阻止其他灌木種子補給到土壤種子庫, 油蒿種子由于具有體積較小和見光萌發的特點, 更有利于萌發和幼苗成長[27, 35–36]; 三是油蒿比楊柴和檸條等灌木的根系分布更淺[37], 對于水分的競爭能力更強, 可以更快、更充分地利用降水[38]。
3.3 種內不同生長發育階段之間的相關關系
種內關聯性是指同一種群不同生長發育階段個體之間在空間分布位置上的相互關聯性, 其在一定程度上反映了種群的生長發育現狀和種群內的相互關系[39]。種內空間正相關和負相關分別反映了種群內互利互惠和競爭排斥的生態關系, 空間無關聯反映了沒有明顯的生態關系[6]。同一個種群在一定區域內生長發育具有相同的適應性, 其不同生長發育階段之間的關聯關系, 會受到種群數量和體積大小、生存環境、種群之間的依賴和競爭關系、歷史因素以及其他因素的影響[24]。
本研究發現油蒿與楊柴不同生長發育階段的種內關聯關系在較廣泛的尺度范圍內表現為正相關, 隨著尺度增大關聯性逐漸降低, 表現為無相關, 甚至油蒿的種內關聯關系出現負相關趨勢。這可能與種群在小尺度下呈現出聚集分布, 大尺度下呈現出隨機或均勻分布有密切聯系[40]。雖然目前有研究表明, 種群的競爭壓力主要是因為種內競爭產生的[41], 但油蒿與楊柴的種內關聯關系在較大尺度范圍內沒有明顯的排斥競爭的現象, 這可能是由于同一種群具有相同的生物學特性造成的。一方面, 同種植被的生境偏好和環境適應性相同, 對于資源的利用具有相似性, 產生生態位重疊, 有利于種群內部互惠互利、共存發展[40]。另一方面, 徑級較小的植株對于環境資源(水分、土壤、養分、地形等)的競爭力較弱[42], 同時種群呈現聚集分布, 形成群體效應[43], 從而徑級大、競爭實力強的植株在一定范圍內對生境的改變使得徑級較小的植株更加適合生存, 彼此交錯分布, 充分共享和利用資源[13, 44]。
該研究還發現灌木(油蒿、楊柴)的生長發育階段越接近, 個體間的正相關性越強; 反之, 植株的生長發育階段差距較大, 個體間的正相關性就越弱, 甚至在大尺度下表現為無相關或負相關。由此可見, 形體大小相近的個體對于種群的協調發展更有利, 而形體差距較大的個體之間可能由于對資源的需求和競爭不能達到平衡穩定狀態, 從而產生種內競爭,呈現出負關聯關系[13]。除此之外, 檸條營養株和生殖株個體之間主要表現為無關聯關系, 這可能由于檸條主要呈現為隨機或均勻分布, 不同齡級之間保持一定距離, 彼此之間互不干擾、獨立生長, 在長時間的群落演替過程中占據適宜的生態位, 減緩了種群內資源爭奪的壓力[13, 44]。
4 結論
研究基于固沙灌木個體的位置信息和生長發育階段信息, 分析了毛烏素沙地固定沙丘上油蒿、楊柴和檸條種群的空間分布格局及種間、種內關聯性。空間分布格局研究表明: 三個種群在小尺度下呈現聚集分布, 大尺度下為隨機或均勻分布; 在研究尺度范圍內, 楊柴和檸條主要呈現隨機分布。這主要是由種子擴散機制、生境偏好、群體效應的生存策略以及密度制約造成的。
油蒿與楊柴兩個種群種間關聯研究表明: 油蒿和楊柴在較小尺度下負相關, 其他尺度下主要表現為無相關關系。種間負相關可能是由于生態位重疊程度小, 對于有限的資源種間競爭激烈引起的。
三個種群的種內關聯研究表明: 檸條的種內關聯關系基本呈無關聯, 油蒿和楊柴的種內關系基本呈正相關, 隨著尺度進一步增加, 會呈現負相關或無關聯。除此之外, 同一種群個體的生長發育階段越接近, 個體間的正相關性越強。
研究結果進一步加深了我們對于毛烏素沙地優勢種群空間格局的認識, 有利于更好地理解毛烏素沙地固沙灌木種群的生物學特性以及形成機制, 為毛烏素沙地防風固沙建設提供了一定參考依據。本研究采用的點格局方法主要利用個體的位置和生長發育階段信息進行分析, 對于個體內部以及個體間差異的關注度不夠。因此, 未來可對樣地內固沙灌木進行動態監測, 結合個體自身的生物學特性以及生境因素, 進一步闡明種群空間格局形成機制。
[1] 楊君瓏, 王輝, 王彬, 等. 子午嶺油松林灌木層主要樹種的空間分布格局和種間關聯性研究[J]. 西北植物學報, 2007, 27(4): 4791–4796.
[2] 郭忠玲, 馬元丹, 鄭金萍, 等. 長白山落葉闊葉混交林的物種多樣性、種群空間分布格局及種間關聯性研究[J]. 應用生態學報, 2004, 15(11): 2013–2018.
[3] FANGLIANG H, LEGENDRE P, LAFRANKIE J V. Distribution patterns of tree species in a Malaysian tropical rain forest[J]. Journal of Vegetation Science, 1997, 8(1): 105–114.
[4] DRUCKENBROD D L, SHUGART H H, DAVIES I. Spatial pattern and process in forest stands within the Virginia piedmont[J]. Journal of Vegetation Science, 2005, 16(1): 37–48.
[5] CONDIT R, ASHTON P S, BAKER P, et al. Spatial patterns in the distribution of tropical tree species[J]. Science, 2000, 288(5470): 1414–1418.
[6] 張金屯. 植物種群空間分布的點格局分析[J]. 植物生態學報, 1998, 22(4): 57–62.
[7] 楊洪曉, 張金屯, 吳波, 等. 毛烏素沙地油蒿種群點格局分析[J]. 植物生態學報, 2006, 30(4): 563–570.
[8] BRISSON J, REYNOLDS J F. The effect of neighbors on root distribution in a creosote bush() population[J]. Ecology, 1994, 75(6): 1693–1702.
[9] 鄭元潤. 不同方法在沙地云杉種群分布格局分析中的適用性研究[J]. 植物生態學報, 1997, 21(5): 89–93.
[10] 戴小華, 余世孝. GIS支持下的種群分布格局分析[J]. 中山大學學報(自然科學版), 2003, 42(1): 75–78.
[11] 尤海舟, 賈成, 樊華, 等. 格局分析的最新方法——點格局分析[J]. 四川林業科技, 2009, 30(6): 106–110.
[12] 王本洋, 余世孝. 種群分布格局的多尺度分析[J]. 植物生態學報, 2005, 29(2): 235–241.
[13] 拓鋒, 劉賢德, 劉潤紅, 等. 祁連山大野口流域青海云杉種群空間格局及其關聯性[J]. 植物生態學報, 2020, 44(11): 1172–1183.
[14] HILL M O. The intensity of spatial pattern in plant communities[J]. The Journal of Ecology, 1973, 61(1): 225–235.
[15] 沈渭壽. 毛烏素沙地3種飛播植物種群分布格局的研究[J]. 中國沙漠, 1998, 18(4): 78–84.
[16] 劉法, 楊海龍, 高甲榮, 等. 毛烏素沙地羊柴種群點格局分析[J]. 中國水土保持科學, 2011, 9(4): 98–103.
[17] 崔強, 高甲榮, 李柏, 等. 寧夏毛烏素沙地3種常用固沙植物種群點格局分析[J]. 生態與農村環境學報, 2011, 27(3): 58–63.
[18] RIPLEY B D. Modelling spatial patterns[J]. Journal of the Royal Statistical Society: Series B (Methodological), 1977, 39(2): 172–192.
[19] 張新時. 毛烏素沙地的生態背景及其草地建設的原則與優化模式[J]. 植物生態學報, 1994, 18(1): 1–16.
[20] 郭柯, 董學軍, 劉志茂. 毛烏素沙地沙丘土壤含水量特點——兼論老固定沙地上油蒿衰退原因[J]. 植物生態學報, 2000, 24(3): 275–279.
[21] 吳波, 慈龍駿. 毛烏素沙地景觀格局變化研究[J]. 生態學報, 2001, 21(2): 191–196.
[22] 魏寧寧, 母艷梅, 姜曉燕, 等. 毛烏素沙地油蒿-楊柴灌叢生態系統蒸散組分分配及其影響因子[J]. 應用生態學報, 2021, 32(7): 2407–2414.
[23] LEWIS A, BRIDLE S. Cosmological parameters from CMB and other data: A Monte-Carlo approach[J]. Physical Review D, 2002, 66(10): 103511.
[24] VEBLEN T T, ASHTON D H, SCHLEGEL F M. Tree regeneration strategies in a lowland Nothofagus-dominated forest in south-central Chile[J]. Journal of Biogeography, 1979, 6(4): 329–340.
[25] HUBBELL S P. Neutral theory and the evolution of ecological equivalence[J]. Ecology, 2006, 87(6): 1387–1398.
[26] 馮麗, 張景光, 張志山, 等. 沙坡頭人工固沙區油蒿繁殖分配的研究[J]. 中國沙漠, 2008, 28(3): 473–477.
[27] 王剛, 梁學功. 沙坡頭人工固沙區的種子庫動態[J]. 植物學報, 1995, 37(3): 231–237.
[28] 韓路, 王海珍, 彭杰, 等. 塔里木河上游天然胡楊林種群空間分布格局與動態研究[J].西北植物學報, 2007, 27(8): 1668–1673.
[29] 肖春旺, 周廣勝, 馬風云,. 施水量變化對毛烏素沙地優勢植物形態與生長的影響[J]. 植物生態學報, 2002, 26(1): 69–76.
[30] 陳玉福, 董鳴. 毛烏素沙地根莖灌木羊柴的基株特征和不同生境中的分株種群特征[J]. 植物生態學報, 2000, 24(1): 40–45.
[31] 彭舜磊, 閆小婷, 齊光, 等. 伏牛山栓皮櫟群落優勢種群種間關聯動態分析[J]. 森林與環境學報, 2018, 38(3): 326–333.
[32] WARD J S, PARKER G R, FERRANDINO F J. Long-term spatial dynamics in an old-growth deciduous forest[J]. Forest Ecology and Management, 1996, 83(3): 189–202.
[33] 李智叁, 李鳳日. 烏蘭布和沙漠天然白刺種群及主要伴生種種間關聯性研究[J]. 植物研究, 2008, 28(1): 98–103.
[34] 張德魁, 王繼和, 馬全林, 等. 油蒿研究綜述[J]. 草業科學, 2007(8): 30–35.
[35] 黃正英, GUTTERMAN Y. 油蒿與中國和以色列沙漠中的兩種蒿屬植物種子萌發策略的比較[J]. 植物學報, 2000, 42(1): 71–80.
[36] 馬驥, 李俊禎, 孔紅. 我國沙區6種蒿屬植物的種子微形態特征[J]. 中國沙漠, 2002, 22(6): 586–590.
[37] 張志山, 樊恒文, 趙金龍, 等. 沙漠人工植物群落的根系分布及動態[J]. 中國沙漠, 2006, 26(4): 637–643.
[38] 唐海萍, 史培軍, 李自珍. 沙坡頭地區不同配置格局油蒿和檸條水分生態位適宜度研究[J]. 植物生態學報, 2001, 25(1): 6–10.
[39] 王磊, 孫啟武, 郝朝運, 等. 皖南山區南方紅豆杉種群不同齡級立木的點格局分析[J]. 應用生態學報, 2010, 21(2): 272–278.
[40] 張金屯, 孟東平. 蘆芽山華北落葉松林不同齡級立木的點格局分析[J]. 生態學報, 2004, 24(1): 35–40.
[41] STOLL P, PRATI D. Intraspecific aggregation alters competitive interactions in experimental plantcommunities[J]. Ecology, 2001, 82(2): 319–327.
[42] 張健, 郝占慶, 宋波, 等. 長白山闊葉紅松林中紅松與紫椴的空間分布格局及其關聯性[J]. 應用生態學報, 2007, 18(8): 1681–1687.
[43] 趙峰俠, 尹林克. 荒漠內陸河岸胡楊和多枝檉柳幼苗種群空間分布格局及種間關聯性[J]. 生態學雜志, 2007, 26(7): 972–977.
[44] 涂洪潤, 李嬌鳳, 劉潤紅, 等. 桂林巖溶石山檵木種群空間格局及其關聯性[J]. 應用生態學報, 2019, 30(8): 2621–2630.
Spatial distribution pattern and spatial correlation of main sand-fixing shrubs in fixed dunes in Yanchi region, Mu Us sandy land
NIU Chengxu1, ZHANG Dinghai1,*, ZHANG Zhishan2, WANG Yanli3
1.Center for Quantitative Biology, College of Science, Gansu Agricultural University, Lanzhou 730070, China 2. Shapotou Desert Research and Experimental Station, Northwest Institute of Eco-Environment and Resources, Lanzhou 730000, China 3. College of Forestry, Gansu Agricultural University, Lanzhou 730070, China
The purpose of this research on the spatial distribution pattern and correlation of populations is to predict the succession trend of populations and vegetation reconstruction which has a great significance meaning for further research investigation. Based on the geographic location and growth stage data of sand-fixing shrubs on fixed sand dunes in the Mu Us Sandy Land, the spatial distribution pattern and correlation of three dominant populations of(AO),(HM) and(CK) were analyzed by using univariate and bivariate paired correlation functions and Monte Carlo fitting test. The results are as follows. (1) The three populations were aggregated at small scale and randomly or uniformly distributed at large scale; the populations of HM and CK showed aggregation distribution within the spatial scale of the study. (2) The interspecies relationships showed thatAO and HM were negatively correlated at the scale less than 5 meters and basically uncorrelated at other scales. (3) The intraspecific correlation among different growth and development stages of the populations showed thatthe intraspecific relationships of the 3 populations were basically positively correlated at small scales. At large scales, there was a negative correlation between the intraspecifc relationships of AO,while there was no correlation between the intraspecifc relationships of HM and CK.
Mu Us sandy land; sand-fixing shrubs; point pattern analysis; spatial distribution pattern; spatial correlation
10.14108/j.cnki.1008-8873.2024.01.001
Q948.15
A
1008-8873(2024)01-001-09
2021-07-30;
2021-09-30
國家自然科學基金項目(42361016); 甘肅省科技創新計劃—“西部之光”人才計劃項目(22JR9KA032);甘肅省科技計劃項目(21JR7RA831)
牛程旭(1997—), 女, 河北沽源人, 碩士研究生, 主要從事應用統計學研究, E-mail: 286116811@qq.com
通信作者:張定海, 男, 副教授, 碩士導師, 主要從事生態水文模型及統計學在生態學、經濟學中的應用, E-mail: zhangdh@gsau.edu.cn
牛程旭, 張定海, 張志山, 等. 毛烏素沙地鹽池地區固定沙丘主要固沙灌木的空間分布格局與空間關聯性研究[J]. 生態科學, 2024, 43(1): 1–9.
NIU Chengxu, ZHANG Dinghai, ZHANG Zhishan, et al. Spatial distribution pattern and spatial correlation of main sand-fixing shrubs in fixed dunes in Yanchi region, Mu Us sandy land[J]. Ecological Science, 2024, 43(1): 1–9.
