玉米多葉矮化突變體lyd1的鑒定與基因克隆
2024-04-28 01:13:44劉孝偉牛群凱時子文侯雨微馮開潔榮廷昭曹墨菊
作物學(xué)報 2024年5期
蘇 帥 劉孝偉 牛群凱 時子文 侯雨微 馮開潔 榮廷昭 曹墨菊
玉米多葉矮化突變體的鑒定與基因克隆
蘇 帥 劉孝偉 牛群凱 時子文 侯雨微 馮開潔 榮廷昭 曹墨菊*
四川農(nóng)業(yè)大學(xué)玉米研究所 / 農(nóng)業(yè)部西南玉米生物學(xué)與遺傳育種重點實驗室, 四川成都 611130
玉米株高降低通常是由節(jié)間數(shù)目減少、節(jié)間長度變短或二者共同作用所致。而本研究在基因編輯后代中發(fā)現(xiàn)的玉米多葉矮化突變體卻表現(xiàn)為節(jié)間數(shù)目顯著增加, 株高顯著降低。株高僅為93.10 cm, 與野生型KN5585的株高159.95 cm相比, 降低了41.79%, 差異達(dá)到極顯著水平。然而葉片數(shù)平均達(dá)到27.8片, 相較野生型平均17.8片葉, 增加56.18%, 差異達(dá)到極顯著水平。遺傳分析表明,的突變表型由1對隱性核基因控制, 通過圖位克隆將控制多葉矮化性狀的基因定位于玉米3號染色體標(biāo)記Indel10和Indel11之間, 物理距離0.74 Mb。對定位區(qū)間內(nèi)13個基因(不包含假基因)的序列進行測序, 發(fā)現(xiàn)僅在第4外顯子出現(xiàn)1個A堿基的替換, 其他基因無差異。編碼一個RNA結(jié)合蛋白, 氨基酸的替換發(fā)生在第3個RNA結(jié)合結(jié)構(gòu)域內(nèi)(RRM3), 導(dǎo)致天冬氨酸轉(zhuǎn)變?yōu)槔i氨酸。突變體的突變位點與已報道的、、、不同,的發(fā)現(xiàn)為進一步解析玉米葉片和節(jié)間發(fā)育平衡的遺傳機制提供了寶貴的材料。
玉米; 葉片數(shù)量; 節(jié)間長度; 基因定位
玉米(L.)是我國糧食和飼料作物, 在保障國家糧食安全方面發(fā)揮重要作用。現(xiàn)階段玉米產(chǎn)量的增長主要通過提升種植密度, 適當(dāng)降低株高以增強抗倒伏能力是提高種植密度的關(guān)鍵[1-2]。玉米節(jié)間數(shù)量(葉片數(shù)量)和節(jié)間長度是決定株高的關(guān)鍵因素, 因此對于玉米葉片數(shù)量和節(jié)間長度以及協(xié)調(diào)二者平衡相關(guān)基因的挖掘具有重要的理論和實踐意義。
玉米節(jié)間長度由節(jié)間居間分生組織細(xì)胞的分裂、分化和伸長決定。植物激素作為重要的信號分子, 參與玉米節(jié)間長度的調(diào)控。[3]、[4-5][6]編碼赤霉素合成途徑的中間產(chǎn)物,[7-9]、[9]編碼DELLA蛋白作為GA信號轉(zhuǎn)導(dǎo)的負(fù)調(diào)控轉(zhuǎn)錄因子, 通過調(diào)控赤霉素的生物合成或信號轉(zhuǎn)導(dǎo)影響玉米節(jié)間長度。生長素以極性運輸?shù)姆绞綄?jié)間長度產(chǎn)生影響,編碼生長素極性運輸?shù)鞍譖GP, 通過促進節(jié)間生長素的積累抑制節(jié)間的伸長, 且對穗下部節(jié)間長度影響較大[10-11]。[12]、[13]位于油菜素內(nèi)酯生物合成的上游,[14]位于油菜素內(nèi)酯生物合成的下游,參與油菜素內(nèi)酯信號轉(zhuǎn)導(dǎo)途徑[15], 上述基因突變后均會對玉米節(jié)間長度產(chǎn)生影響。編碼乙烯生物合成中的關(guān)鍵限速酶ACC合成酶, 通過增強蛋白的穩(wěn)定性, 促進乙烯合成, 抑制節(jié)間伸長[16]。玉米葉片由莖頂端分生組織的周緣區(qū)以一定的時間間隔和空間間隔分化形成[17], 但具體的分子機制尚未解析。迄今已經(jīng)報道的葉片數(shù)量相關(guān)突變體主要與生長素和細(xì)胞分裂素有關(guān)。玉米基因編碼IAA生物合成途徑中的色氨酸轉(zhuǎn)移酶,突變體展示出葉片數(shù)目顯著減少而導(dǎo)致的矮化表型[18]。編碼細(xì)胞分裂素誘導(dǎo)的A型反應(yīng)調(diào)節(jié)因子, 玉米突變體葉片數(shù)增多, 但對株高沒有顯著影響。在突變體中, 玉米IAA外排載體在初始葉原基的表達(dá)量大幅下降, 其生長素水平也明顯降低[19]。
RNA結(jié)合蛋白是含有一個或多個RNA結(jié)合結(jié)構(gòu)域, 通過結(jié)合RNA并改變結(jié)合RNA的命運或功能的蛋白質(zhì)[20-21]。RNA結(jié)合蛋白的研究主要在哺乳動物和細(xì)菌系統(tǒng)中進行, 植物RNA結(jié)合蛋白研究較少。是在酵母中發(fā)現(xiàn)的一個編碼RNA結(jié)合蛋白的基因, 含有3個RNA結(jié)合結(jié)構(gòu)域, 促進酵母細(xì)胞減數(shù)分裂S期DNA的合成和減數(shù)第一次分裂的有序進行[22]。水稻基因編碼Mei2-like蛋白, 該基因突變后植株矮小, 葉片數(shù)增加, 植株早熟[23]。是在玉米中的同源基因, 該基因在葉原基起始處表達(dá)量最低, 其突變體株高降低, 葉片數(shù)目增多, 葉脈紊亂, 植株雄穗缺失, 原雄穗發(fā)育處展示出雌性化的特征[33-34]。盡管在植物中很多基因已經(jīng)被發(fā)現(xiàn), 且均與酵母基因類似, 含有3個RNA結(jié)合結(jié)構(gòu)域, 但在植物中未找到酵母下游基因的同源物, 表明該基因在酵母和植物中的功能和作用機制并不相同[24]。在植物中基因下游結(jié)合的RNA的挖掘, 以及3個RNA結(jié)合結(jié)構(gòu)域在發(fā)揮功能過程中的作用仍需要進一步深入研究。
本研究以多葉矮化突變體()為研究對象, 對突變體的葉片數(shù)和節(jié)間長度等表型特征進行了系統(tǒng)調(diào)查, 采用分子標(biāo)記技術(shù)對突變基因進行了定位, 并對候選基因進行預(yù)測和分析, 確定了關(guān)鍵候選基因, 該基因編碼RNA結(jié)合蛋白。本研究為進一步闡明玉米葉片數(shù)量和節(jié)間長度的平衡關(guān)系以及RNA結(jié)合蛋白對玉米生長發(fā)育的影響提供了寶貴材料。
1 材料與方法
1.1 試驗材料
玉米多葉矮化突變體、玉米自交系B73、Mo17和KN5585。突變體選自基因()的CRISPR/Cas9敲除后代, 對突變體中的進行測序, 結(jié)果顯示突變體中未發(fā)生編輯, 即該突變體的突變表型與無關(guān)。
1.2 突變體lyd1的表型鑒定和遺傳分析
2020年6月在四川農(nóng)業(yè)大學(xué)溫江試驗基地發(fā)現(xiàn)該多葉矮化突變體, 將其自交保種。2020年12月在四川農(nóng)業(yè)大學(xué)云南試驗基地種植、Mo17和B73, 并將分別與Mo17和B73雜交得到F1。2021年3月在四川農(nóng)業(yè)大學(xué)溫江試驗基地種植F1, 自交獲得F2種子。2022年3月在四川農(nóng)業(yè)大學(xué)溫江試驗基地種植F2群體。成熟期統(tǒng)計F2分離群體中正常和多葉矮化植株的分離比并進行卡平方測驗。
2021年3月將野生型KN5585和突變體種植于四川農(nóng)業(yè)大學(xué)溫江試驗基地, 野生型KN5585二葉期后, 每間隔3 d, 統(tǒng)計野生型KN5585和突變體各30株完全展開葉數(shù)量。在植株成熟期, 田間隨機選取野生型KN5585和各15株, 考察株高、穗位、葉片長度、葉片寬度、雄穗分支數(shù)、地上部分各節(jié)間長度和散粉期等重要農(nóng)藝性狀。使用Microsoft Excel 2016進行數(shù)據(jù)處理和繪圖。
1.3 莖稈細(xì)胞學(xué)觀察
植株散粉后, 選取野生型KN5585和地上部分最長節(jié)間(均為雌穗下方第2節(jié), 野生型KN5585為第11節(jié), 突變晰完體為第19節(jié))的中部, 縱切并將其固定于FAA固定液中。將樣品依次進行軟化、洗滌、脫水、透明、包埋、切片、烤片、染色, 在顯微鏡下進行節(jié)間細(xì)胞觀察, 野生型KN5585和突變體分別選取細(xì)胞形態(tài)清整的3張切片, 共10個視野進行照相。通過ImageJ軟件進行細(xì)胞長度測量, 并根據(jù)實際測量得出的最長節(jié)間平均長度進行節(jié)間細(xì)胞數(shù)目的計算, 使用Microsoft Excel 2016進行數(shù)據(jù)處理和繪圖。
1.4 基因定位
利用CTAB法[25]提取玉米自交系Mo17、KN5585、突變體以及[×Mo17] F2分離群體內(nèi)多葉矮化單株的基因組DNA。利用均勻分布在玉米10條染色體上1050對Indel和SSR標(biāo)記, 對Mo17和進行多態(tài)性分析。在[×Mo17] F2群體中, 選取10株多葉矮化單株提取葉片的DNA, 稀釋成相同濃度并等體積混合, 每5株混合為一個池, 構(gòu)建2個突變池。同時, 另外選取10株表型正常的單株提取葉片的DNA, 稀釋成相同濃度并等體積混合為一個正常池。采用BSA (bulk segregant analysis)方法[26]篩選在正常池和突變池之間具有穩(wěn)定性差異的多態(tài)性標(biāo)記, 并利用上述標(biāo)記對F2群體內(nèi)多葉矮化單株進行基因分型, 篩選與突變表型連鎖的多態(tài)性標(biāo)記, 對目標(biāo)基因進行初步定位。利用NCBI在線引物設(shè)計網(wǎng)站根據(jù)初步定位區(qū)段內(nèi)序列信息, 設(shè)計產(chǎn)物片段600 bp左右的引物, 通過PCR擴增出親本基因片段后進行測序, 利用DNAMAN軟件對所擴增的雙親基因序列進行比對分析, 找出雙親間的序列差異位點, 根據(jù)差異位點開發(fā)片段大小為150~300 bp的Indel標(biāo)記。利用新開發(fā)的標(biāo)記, 對擴大的定位群體內(nèi)多葉矮化單株進行基因分型, 縮小定位區(qū)間, 對目的基因進行精細(xì)定位。
1.5 候選基因的預(yù)測與分析
在MaizeGDB和NCBI網(wǎng)站上, 獲取精細(xì)定位區(qū)間內(nèi)基因的注釋信息。根據(jù)精細(xì)定位區(qū)間內(nèi)基因序列和注釋, 設(shè)計基因特異引物, 以野生型和基因組DNA或全長CDS為模板, 利用KOD高保真DNA聚合酶進行PCR擴增。擴增體系為30 μL: 2×PCR buffer for KOD FX 15 μL、正向和反向引物(10 μmol L–1)各2 μL、KOD FX 0.6 μL、dNTPs (0.002 mol L–1) 6 μL、ddH2O 3.4 μL、模板3 μL, PCR程序為: 95℃預(yù)變性5 min; 98℃變性10 s, 58℃退火30 s, 68℃延伸(1 kb min–1), 循環(huán)數(shù)為35次; 68℃ 10 min。PCR擴增產(chǎn)物送公司進行測序并利用DNAMAN軟件進行序列比對, 確定變異位點。
以定位群體內(nèi)關(guān)鍵交換單株DNA為模板, 根據(jù)變異位點附近序列設(shè)計特異性引物, 對變異位點所在的基因片段進行擴增, 長度約600 bp, 變異位點位于擴增片段中間, 并將PCR擴增產(chǎn)物送公司進行測序, 利用DNAMAN軟件進行序列比對, 進行變異位點群體內(nèi)共分離驗證。
1.6 關(guān)鍵候選基因的表達(dá)及其蛋白的亞細(xì)胞定位
為了檢測在不同時期不同組織的表達(dá)模式, 利用TRIzol試劑盒提取野生型KN5585在V3、V7、V11、V17時期的根、莖、葉、雌穗、雄穗的RNA; 同時為了檢測突變體中基因表達(dá)水平變化, 利用TRIzol試劑盒提取野生型KN5585和的V3和V7時期莖尖的RNA, 電泳檢測RNA的完整性, 并將質(zhì)量符合標(biāo)準(zhǔn)的RNA反轉(zhuǎn)錄得到cDNA, 根據(jù)玉米公共數(shù)據(jù)庫MaizeGDB中提供的候選基因cDNA信息, 使用Primer 3在線設(shè)計特異性引物, 引物擴增片段長度在100~200 bp左右。cDNA稀釋至100 ng μL–1作為模板。以作為內(nèi)參。qRT-PCR反應(yīng)在CFX 96 Real-Time System上進行。qRT-PCR反應(yīng)體系為10 μL: Diluted cDNA 1 μL, 2×SYBR Green PCR Mix 5 μL, 上游引物(10 μmol L–1) 0.4 μL, 下游引物(10 μmol L–1) 0.4 μL, ddH2O 3.2 μL。反應(yīng)程序為: 95℃預(yù)變性3 min; 95℃變性10 s, 58℃退火30 s, 72℃延伸15 s, 40個循環(huán), 3次生物重復(fù), 3次技術(shù)重復(fù)。擴增結(jié)束后產(chǎn)生的數(shù)據(jù)導(dǎo)入Microsoft Excel中, 采用2–DDCt方法進行基因相對表達(dá)量的分析。
為了檢測蛋白在細(xì)胞內(nèi)的定位模式, 設(shè)計帶有PCAMBIA2300-eGFP載體同源臂的引物擴增全長轉(zhuǎn)錄本。利用同源重組的方法, 將pCAMBIA2300-35S-eGFP載體和目的片段連接, 并轉(zhuǎn)入大腸桿菌感受態(tài), 挑選陽性克隆檢測并測序比對, 挑選正確序列的單克隆擴繁, 提取質(zhì)粒。將質(zhì)粒轉(zhuǎn)化GV3101農(nóng)桿菌, 200轉(zhuǎn) min–128℃過夜擴繁至OD600為0.6左右, 然后5000轉(zhuǎn) min–1離心5 min收集菌體, 棄上清, 加入適量的注射液。用已滅菌的1 mL注射器將菌體懸浮液注射至4~8周左右的煙草葉背部表皮細(xì)胞層中, 然后在正常條件下培養(yǎng)48 h, 用熒光顯微觀察并采集圖像。
2 結(jié)果與分析
2.1 突變體的表型鑒定及遺傳分析
為了明確突變體的遺傳規(guī)律, 將自交后代于每年的春季在四川、秋季在云南進行多年種植觀察,的突變表型不受環(huán)境影響, 可以穩(wěn)定遺傳。將分別與Mo17和B73雜交, F1代植株均表現(xiàn)正常表型, 將F1自交獲得F2, 在5380株[×Mo17] F2群體中, 正常植株數(shù)量為4083株, 多葉矮化株數(shù)量為1297株, 經(jīng)卡平方檢測, 正常植株與多葉矮化株分離比符合3∶1 (c2C=2.24, 小于c20.05,1=3.84); 在286株[×B73] F2群體中, 正常植株數(shù)量為220株,多葉矮化株數(shù)量為66株, 經(jīng)卡平方檢測, 正常植株與多葉矮化株分離比符合3∶1 (c2C=0.46, 小于c20.05,1=3.84)。不同遺傳背景下的F2群體, 正常植株與多葉矮化植株分離比均符合3∶1, 表明該突變表型是由單個隱性核基因控制。
對突變體的農(nóng)藝性狀進行調(diào)查發(fā)現(xiàn),株高93.10 cm、穗位高43.48 cm, 僅為野生型株高159.95 cm、穗位高64.37 cm的58.21%和67.55% (圖1-A~C), 進一步分析表明穗上部和穗下部節(jié)間縮短是造成株高降低的原因(圖2-A)。生育期內(nèi)完全葉數(shù)量平均為27.80±1.18片, 野生型生育期內(nèi)完全葉數(shù)量平均為(17.80±0.95)片(圖2-B), 差異達(dá)到極顯著水平。野生型從播種至群體內(nèi)50%植株散粉需要(89.03±2.10) d,從播種至群體內(nèi)50%植株散粉需要(88.64±1.79) d, 二者無顯著差異(表1)。綜合葉片數(shù)量及生育期的比較結(jié)果可以發(fā)現(xiàn),相鄰2片葉片完全展開的時間間隔為2.31 d, 野生型相鄰2片葉片完全展開的時間間隔為3.50 d, 兩者差異達(dá)到顯著水平,相鄰葉原基起始所用時間間隔縮短, 葉片起始速率加快。此外葉片長度和寬度較野生型顯著縮小(圖1-D), 穗長、穗粗、行粒數(shù)、穗粒重、百粒重均顯著下降(圖1-F, G), 雄穗分支數(shù)顯著減少, 雄穗基部由未完全伸展的葉片包裹(圖1-E和表1), 這可能也暗示了雄穗基部節(jié)間伸長受阻。

圖1 野生型KN5585和突變體lyd1的性狀比較
A、B、C: 分別表示五葉期、十葉期和成熟期的野生型KN5585和植株; D: 野生型KN5585和成熟期的葉片表型; E: 野生型KN5585和突變體的雄穗表型; F、G: 野生型KN5585和的果穗和籽粒的表型。標(biāo)尺: A 15 cm; B、C、D 30 cm; E 5 cm; F 2 cm。
A, B, and C: represent the 5th leaf stage, 10th leaf stage, and mature stage of wild type KN5585 and mutant, respectively. D: leaves phenotypes of wild type KN5585 and mutantat maturity. E: the tassel phenotype of wild type KN5585 and mutant. F, G: phenotypes of ear and kernel of wild types KN5585 and mutant. Bar: A 15 cm; B, C, D 30 cm; E 5 cm; F 2 cm.

圖2 野生型KN5585和突變體lyd1的節(jié)間、葉片發(fā)育的比較
A: 野生型KN5585和地上部分節(jié)間長度的比較, 0表示雌穗所在節(jié)間、–1表示雌穗下方第一節(jié)、+1表示雌穗上方第一節(jié), 依此類推; B: 野生型KN5585和葉片發(fā)育過程比較。縱坐標(biāo)表示調(diào)查群體內(nèi)植株葉片的平均數(shù), 橫坐標(biāo)表示調(diào)查日期。
A: the comparison of internodes length between the wild type KN5585 and mutant, 0 represents the internode of the ear, –1 represents the first internode below the ear, +1 represents the first internode above the ear, and so on. B: the comparison of leaves development between wild type KN5585 and mutant. The vertical axis shows the average number of leaves in the surveyed population. The horizontal axis indicates the survey date.
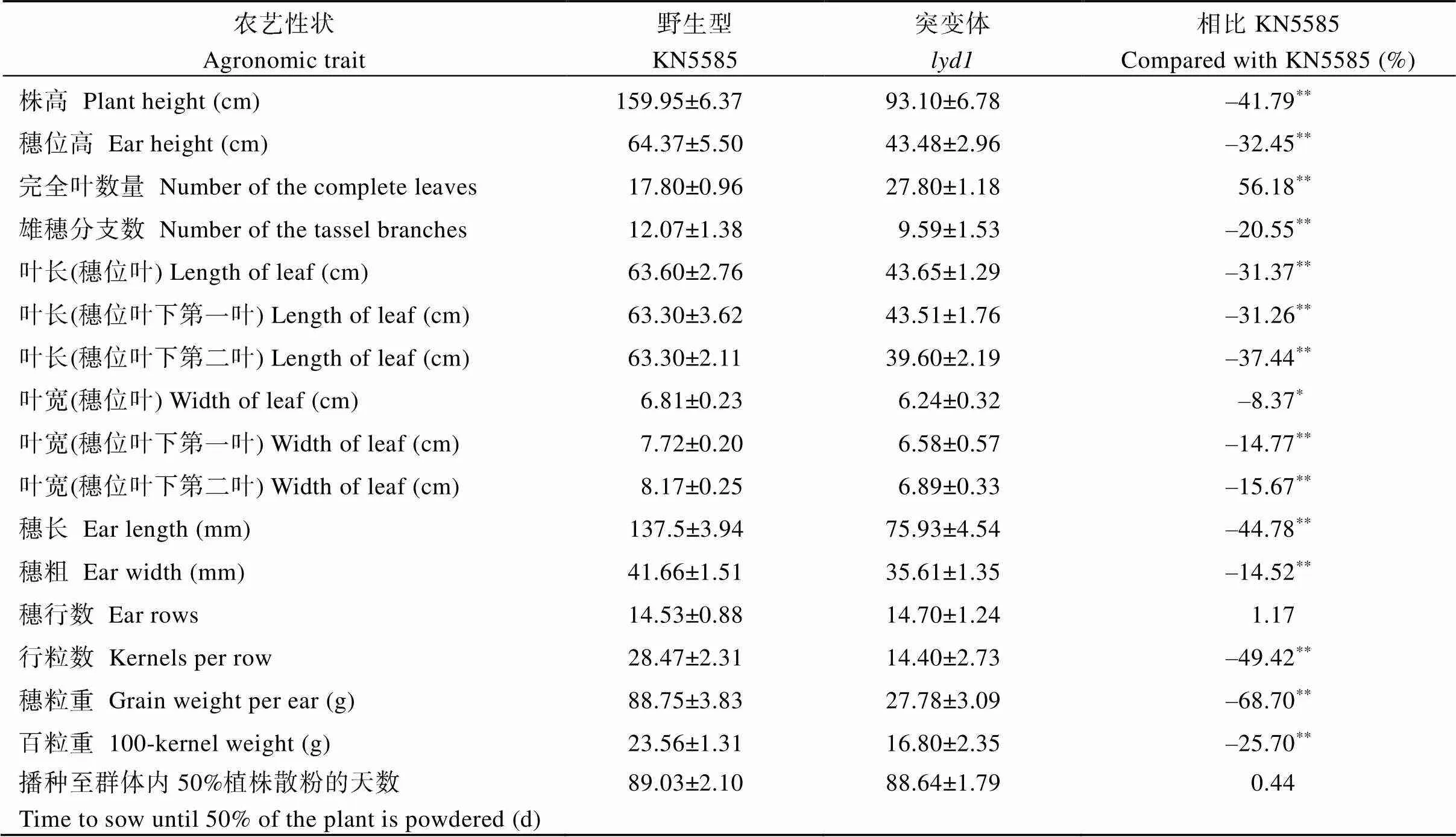
表1 突變體lyd1和野生型KN5585的農(nóng)藝性狀數(shù)據(jù)分析
表中數(shù)據(jù)以“平均值±標(biāo)準(zhǔn)差”形式呈現(xiàn);**表示< 0.01差異極顯著,*表示< 0.05差異顯著。
Data are present “means ± SDs” in the table;**:< 0.01;*:< 0.05.
對和野生型最長節(jié)間進行石蠟切片觀察(圖3-A, B)。結(jié)果表明,最長節(jié)間的10個細(xì)胞縱向長度為17.70 μm, 與野生型KN5585最長節(jié)間的10個細(xì)胞縱向長度22.13 μm相比, 減少了20.02%, 差異達(dá)到極顯著水平;最長節(jié)間細(xì)胞數(shù)目36,003.50個, 野生型KN5585最長節(jié)間細(xì)胞數(shù)目44,984.00個, 差異達(dá)到極顯著水平(圖3-C, D)。因此,節(jié)間縮短是由于節(jié)間細(xì)胞數(shù)目減少和細(xì)胞縱向長度縮短共同導(dǎo)致。

圖3 野生型KN5585和突變體lyd1散粉后雌穗穗位下第二節(jié)節(jié)間的石蠟切片觀察
A: 散粉后野生型KN5585最長節(jié)間(第11葉所在節(jié)間)細(xì)胞觀察; B: 散粉后突變體最長節(jié)間(第19葉所在節(jié)間)細(xì)胞觀察; C, D: 野生型KN5585最長節(jié)間和最長節(jié)間細(xì)胞長度和數(shù)量的比較。標(biāo)尺: 5 μm;**表示< 0.01差異極顯著。
A: the observation of longest intersegmental (the internode where the 11th leaf is located.) cells of wild type KN5585 after loose powder; B: the observation of longest intersegmental (the internode where the 19th leaf is located.) cells of mutantafter loose powder; C, D: the comparison of longest intersegmental cell length and number between wild type KN5585 and mutant. Bar: 5 μm;**:< 0.01
2.2 突變基因的定位
將[×Mo17] F2作為定位群體, 利用均勻分布在玉米10條染色體上的SSR和Indel標(biāo)記對、Mo17進行多態(tài)性分析, 從中共篩選到172對具有雙親多態(tài)性的標(biāo)記。將篩選得到的多態(tài)性標(biāo)記對正常基因池和突變基因池進行BSA混池分析, 結(jié)果顯示位于3號染色體上3對多態(tài)性標(biāo)記Bnlg1019、Indel1、Indel9 (表2)在正常基因池和突變基因池之間存在差異。進一步利用[×Mo17] F2中的93株多葉矮化單株進行基因分型, 發(fā)現(xiàn)標(biāo)記Bnlg1019、Indel1和Indel9的交換單株分別為35、34和9個, 因此將候選基因定位于玉米3號染色體標(biāo)記Indel1和Indel9之間。在Indel1和Indel9之間開發(fā)出8對新的多態(tài)性Indel標(biāo)記, 利用8對Indel標(biāo)記對[×Mo17] F2中93個多葉矮化單株進行基因分型, 發(fā)現(xiàn)Indel2、Indel3、Indel4、Indel5、Indel6、Indel7、Indel10、Indel8 (表2)的交換單株數(shù)分別為30、15、10、7、6、2、0和8個, 將定位區(qū)段鎖定在標(biāo)記Indel7和Indel8之間, 物理距離10.94 Mb (圖4-A)。
為了精細(xì)定位, 在標(biāo)記Indel7和Indel8之間新開發(fā)出5對有多態(tài)性的標(biāo)記。同時擴大定位群體, 利用新開發(fā)的5對多態(tài)性標(biāo)記對[×Mo17] F2群體中的剩余的1204株多葉矮化單株進行基因分型, Indel7、Indel10、SNP1、Indel11、Indel12、Indel13和Indel8 (表2)的交換單株數(shù)分別為35、6、0、11、11、39和89個, 最終將候選基因定位在Indel10和Indel11標(biāo)記之間, 物理距離約0.74 Mb (圖4-B, C)。
2.3 候選基因分析
根據(jù)B73_V5版本的基因注釋信息, 0.74 Mb定位區(qū)間中共有14個候選基因, 其中存在1個假基因(表3)。根據(jù)候選基因的基因組和CDS序列的長度及擴增的難易程度, 對基因組序列較短且較容易擴增的基因擴增其基因組序列, 否則擴增其CDS序列, 并將擴增產(chǎn)物進行測序。結(jié)果顯示KN5585和之間在基因組上、、、、、和均無序列差異, 在全長CDS上、、、、均無序列差異。而()在基因組上存在1個堿基的替換, 為了驗證該堿基替換的真實性, 對全長CDS序列進行擴增, 發(fā)現(xiàn)該位置的堿基替換在的基因組和CDS序列中穩(wěn)定存在(圖5-A)。全長4132 bp, 只有1個轉(zhuǎn)錄本, 包含6個外顯子和5個內(nèi)含子, 編碼664個氨基酸, 具有3個典型的RNA結(jié)合結(jié)構(gòu)域(RRM), 編碼一個RNA結(jié)合蛋白。中堿基替換發(fā)生在第4外顯子上, 位于全長CDS第1478堿基由A堿基轉(zhuǎn)換為T堿基, 導(dǎo)致第492個氨基酸由天冬氨酸轉(zhuǎn)變?yōu)槔i氨酸, 該突變的氨基酸殘基位于RRM3結(jié)構(gòu)域內(nèi)。RRM3在玉米、水稻、擬南芥、小麥、高粱、大豆、藜麥等植物中保守性較高, 并且突變的天冬氨酸也具有較高保守性(圖5-B, C)。
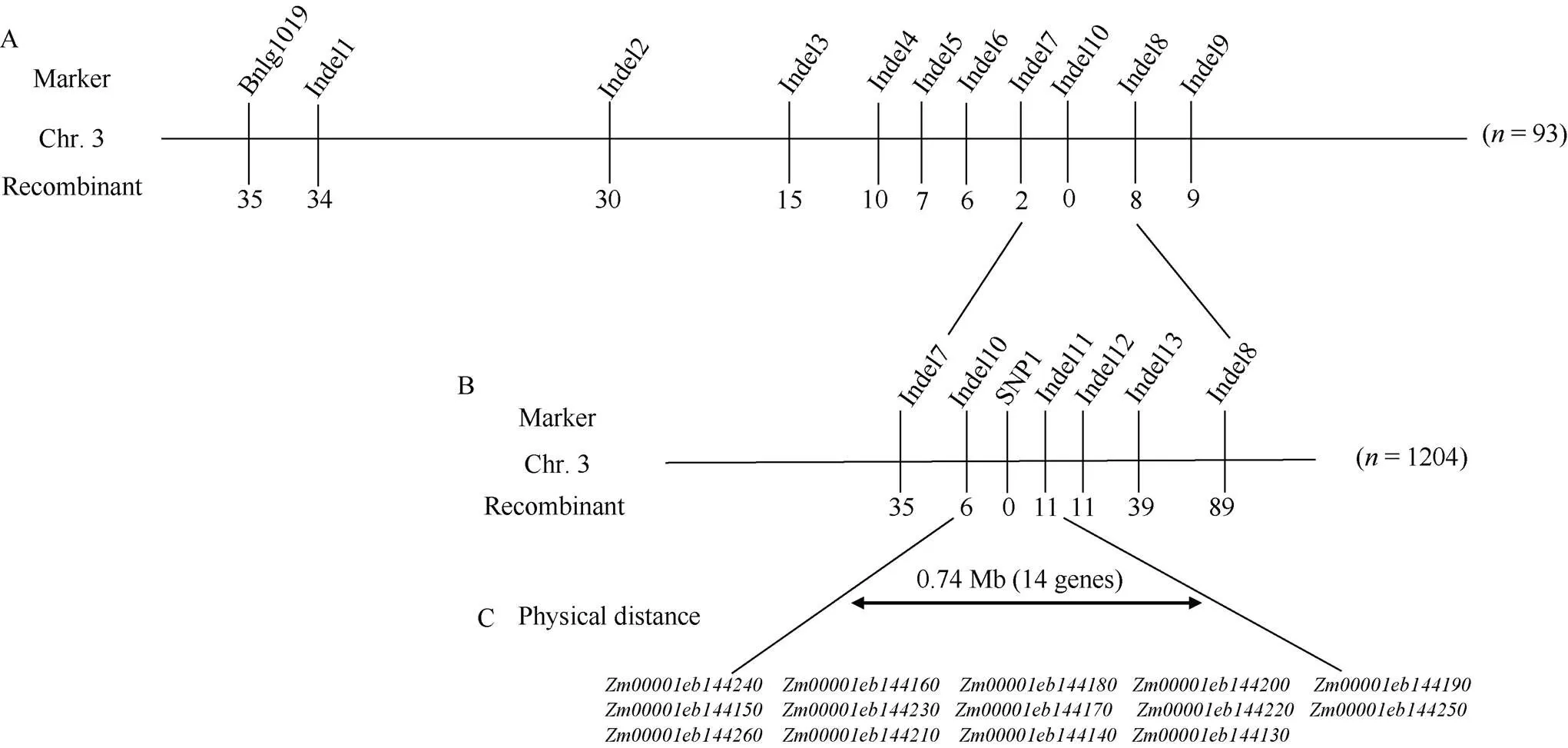
圖4 突變體lyd1的精細(xì)定位
A: 候選基因初步定位在3號染色體Indel7和Indel8分子標(biāo)記之間; B: 候選基因精細(xì)定位在3號染色體Indel10和Indel11分子標(biāo)記之間; C: 精細(xì)定位區(qū)間內(nèi)包含14個候選基因。橫線上方為分子標(biāo)記, 橫線下方為重組單株數(shù)。
A: the candidate gene was preliminarily located between the Indel7 and Indel8 molecular markers on chromosome 3; B: the candidate gene was finely positioned between the Indel10 and Indel11 molecular markers on chromosome 3; C: there are 14 candidate genes in the fine localization interval. Above the horizontal line is the molecular marker, below the horizontal line is the number of recombinant individual plants.
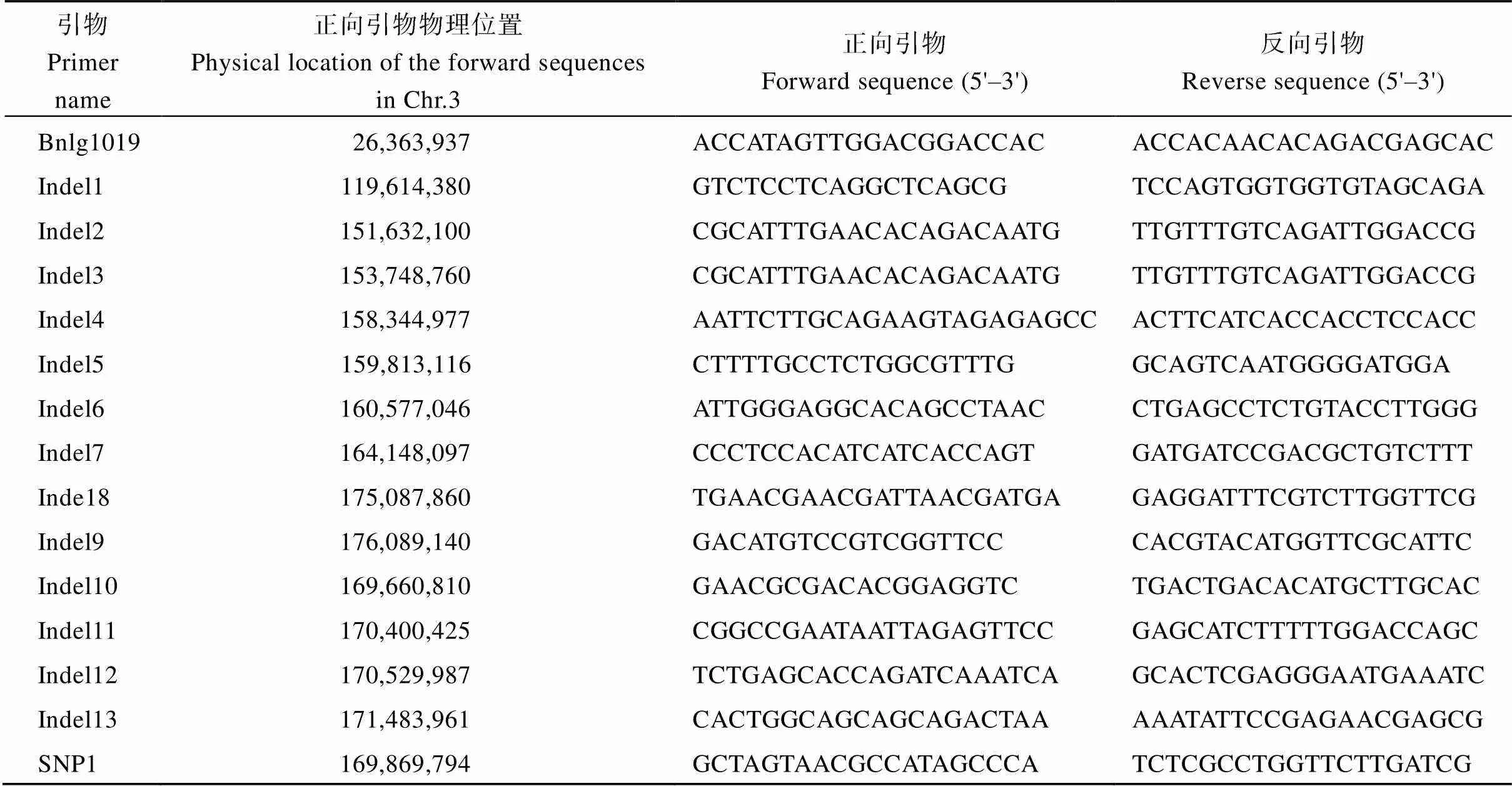
表2 基因定位相關(guān)標(biāo)記信息
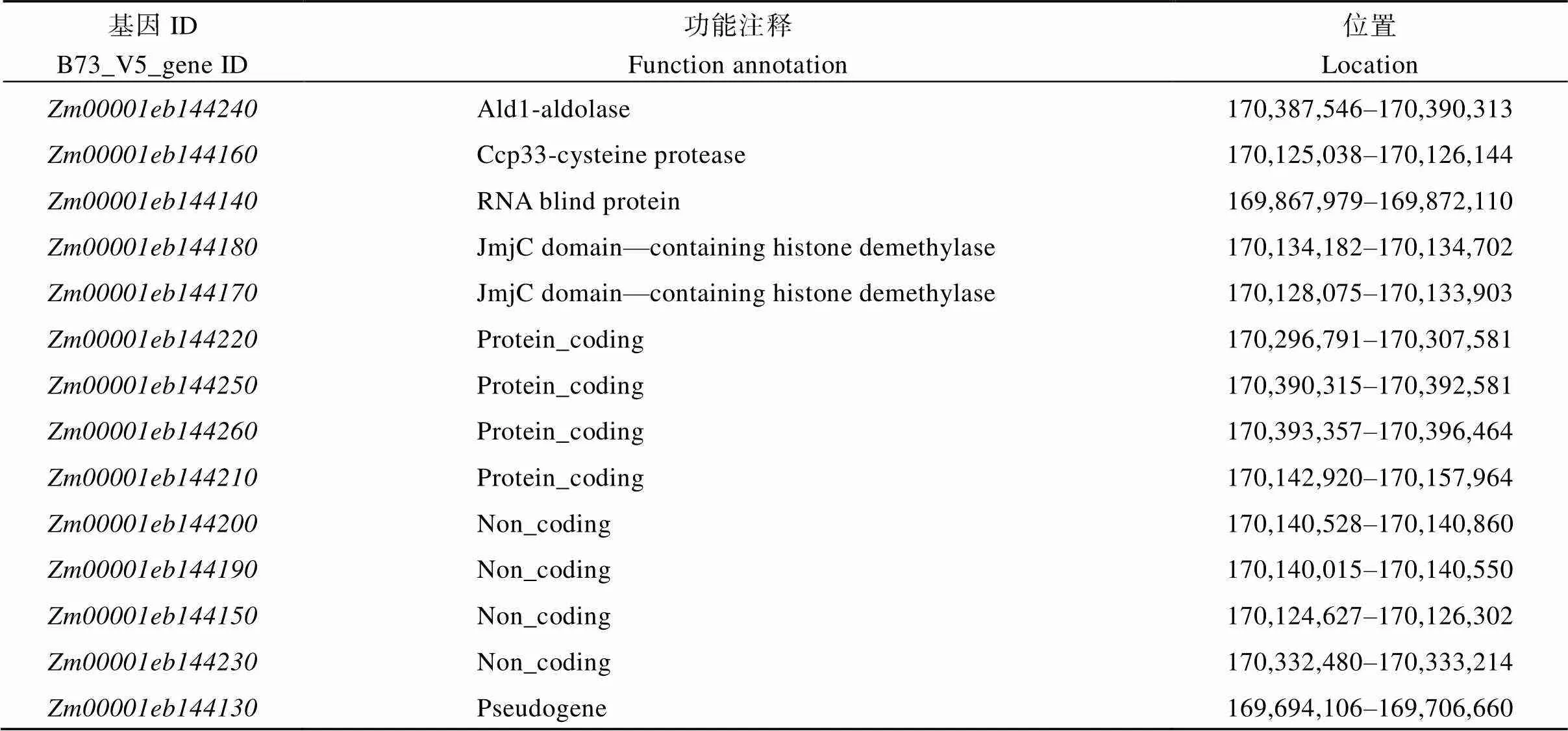
表3 定位區(qū)間內(nèi)候選基因功能注釋
根據(jù)在和野生型差異位點設(shè)計特異性SNP引物(SNP1), 以標(biāo)記Indel10和Indel11在[×Mo17] F2定位群體中檢測到的17個關(guān)鍵交換單株的基因組DNA為模板, 進行擴增測序, 測序結(jié)果顯示17個單株在該位點都顯示的T堿基(圖6), 以上結(jié)果表明該位置發(fā)生的堿基替換與多葉矮化突變表型共分離。
2.4 ZmTE1的表達(dá)分析
利用qRT-PCR分析了在野生型的表達(dá)特征, 結(jié)果顯示在莖尖和幼嫩的雌穗中表達(dá)量較高, 在葉片、莖、雄穗中表達(dá)量很低, 在根中幾乎不表達(dá)(圖7-A)。同時分析了V3和V7時期在野生型與莖尖中的表達(dá)情況, 結(jié)果顯示2個時期在突變體和野生型之間的表達(dá)量均未達(dá)顯著差異(圖7-B), 說明A堿基到T堿基的替換可能不影響該基因在莖尖的表達(dá)。
為進一步了解蛋白的定位情況, 對蛋白進行亞細(xì)胞定位分析, 結(jié)果顯示GFP-的GFP信號與核、膜Marker的信號很好地重疊(圖8-A), 表明蛋白定位在細(xì)胞核和細(xì)胞膜上, 這與RNA結(jié)合蛋白在轉(zhuǎn)錄水平對細(xì)胞核內(nèi)的mRNA進行加工的功能一致。
3 討論
玉米的株高通常由節(jié)間的數(shù)目和節(jié)間長度決定, 一般來說節(jié)間數(shù)目(葉片數(shù)目)越多, 節(jié)間越長植株越高。目前已報道的玉米矮化突變體, 有些葉片數(shù)目不變, 株高降低由節(jié)間長度縮短導(dǎo)致, 如[10-11]、[27]、[7-9]、[28]過表達(dá)株系[29]等, 有些為葉片數(shù)目減少和節(jié)間長度縮短, 但葉片數(shù)減少是導(dǎo)致株高降低的主要原因, 如[18]、過表達(dá)株系[30]等。此外還有些玉米矮化突變體, 表現(xiàn)為節(jié)間數(shù)量的增加, 其株高降低是由節(jié)間長度縮短導(dǎo)致, 如突變體葉片數(shù)較野生型增加3.2片[31];突變體葉片數(shù)較野生型增加1.6片[32];突變體葉片數(shù)較野生型增加1.75片[16]。而本研究發(fā)現(xiàn)的多葉矮化突變體葉片數(shù)較野生型KN5585增加10片葉, 增加了56.18%, 節(jié)間嚴(yán)重縮短, 為研究節(jié)間數(shù)目和節(jié)間長度的平衡提供了寶貴材料。
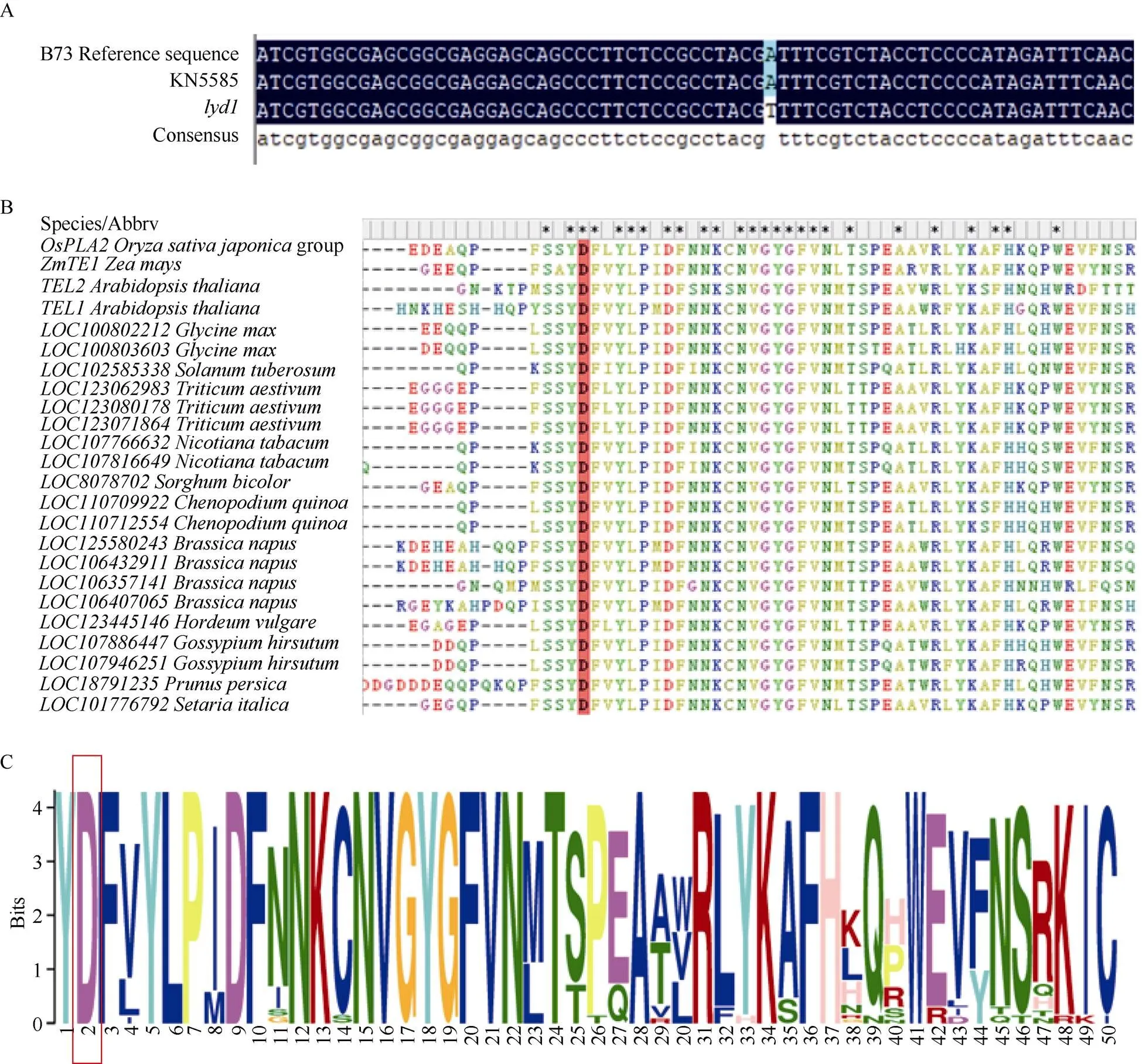
圖5 lyd1突變位點及突變位點保守性分析
A: 突變體與野生型KN5585的CDS差異序列比較; B, C:蛋白序列突變氨基酸殘基保守性分析
A: the comparison of CDS difference sequence ofin mutantand wild type KN5585; B, C: the conservative analysis of amino acid residues mutated inprotein sequence.
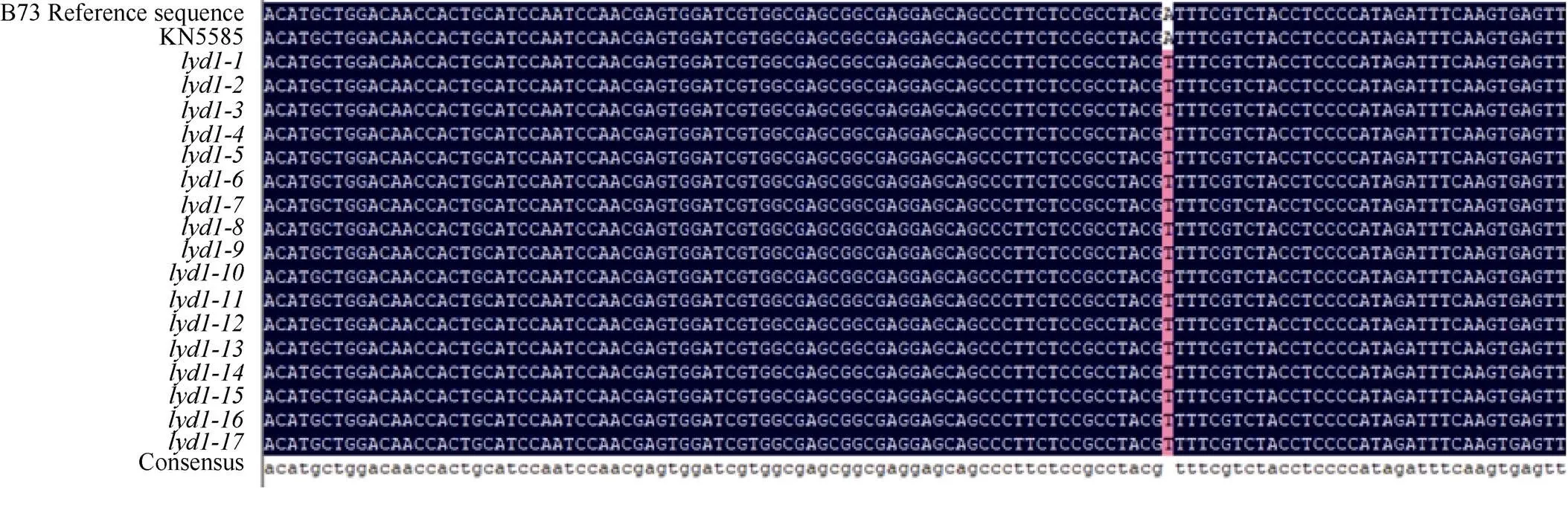
圖6 野生型KN5585和突變體lyd1的序列差異位點群體驗證
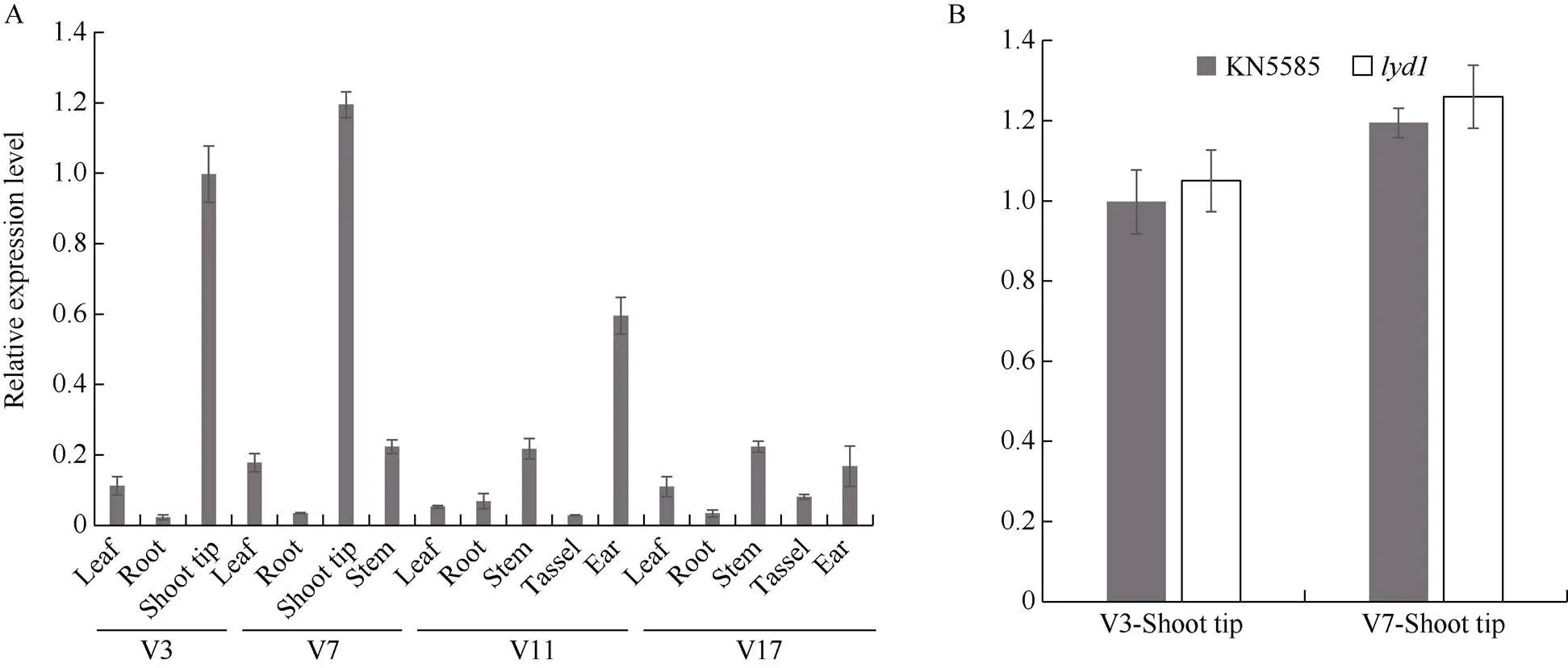
圖7 ZmTE1組織表達(dá)分析
A:在野生型KN5585不同組織中的表達(dá); B:在野生型KN5585和突變體中的表達(dá)比較。V3、V7、V11、V17分別表示玉米植株分別有3、7、11、17片完全展開葉所處的時期。
A: the relative expression level ofgenes in different tissues of wild type KN5585; B: the comparison ofexpression in wild type KN5585 and mutant. V3, V7, V11, and V17 represent the stage when the maize has 3, 7, 11, and 17 fully unfolded leaves, respectively.
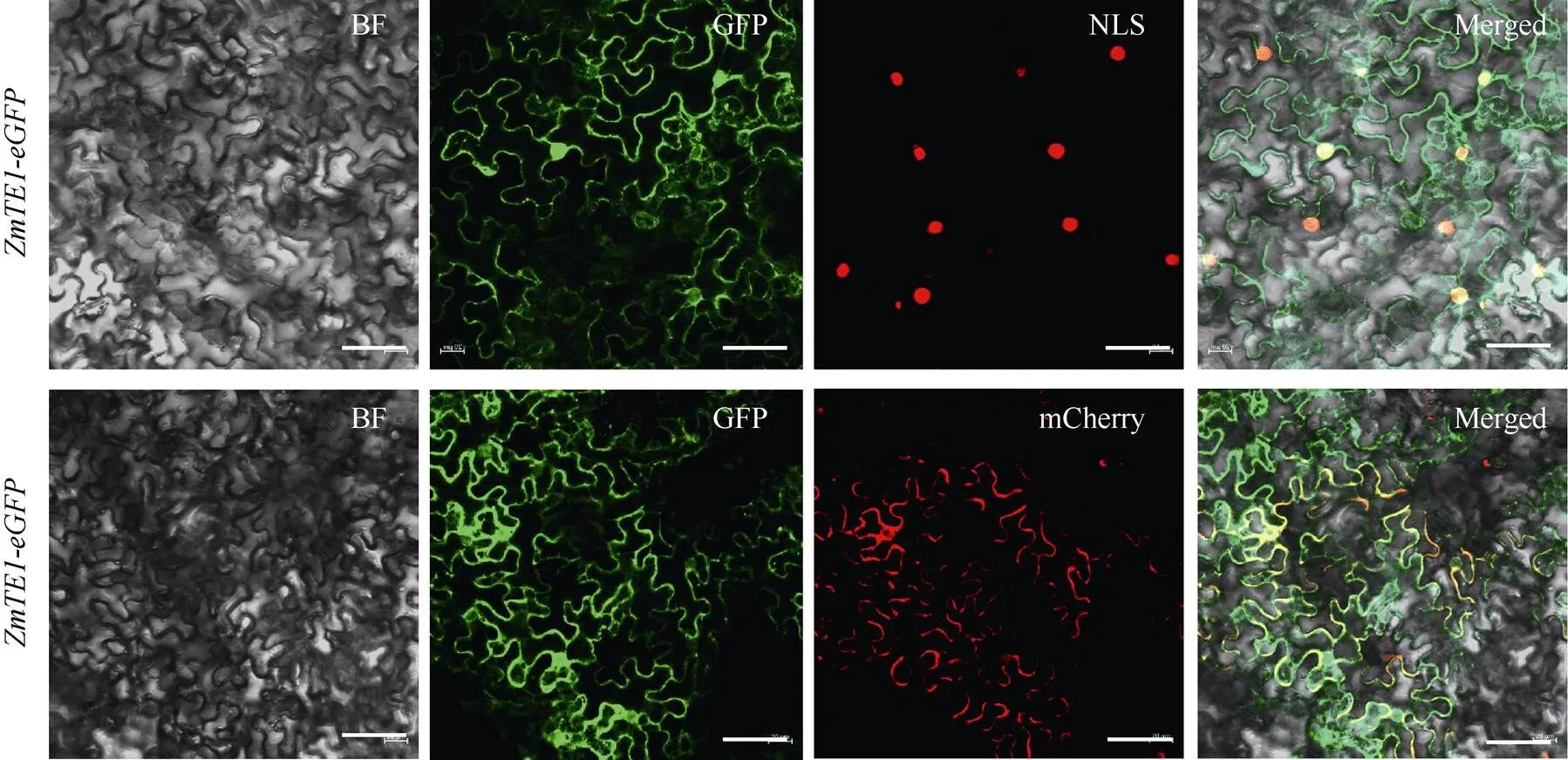
圖8 ZmTE1的亞細(xì)胞定位
PC2300-eGFP-與NLS (細(xì)胞核標(biāo)記)和mCherry (細(xì)胞膜標(biāo)記)共定位, 標(biāo)尺為50 μm。
PC2300-eGFP-are colocalized with NLS (nuclear marker) and mCherry (cell membrane marker). Bar: 50 μm.
控制突變表型的基因被定位在玉米3號染色體, 位于標(biāo)記Indel10和Indel11之間, 物理距離0.74 Mb, 將確定為的關(guān)鍵候選基因。與已報道的、、突變體的突變位點不同,和突變體中轉(zhuǎn)座子插入在第3外顯子,突變體中轉(zhuǎn)座子插入在第1內(nèi)含子[33],突變體中第1外顯子單堿基的替換導(dǎo)致蛋白翻譯提前終止[34], 上述已報道的突變體植株生活力較弱, 難以完成正常生長發(fā)育, 給后續(xù)研究帶來困難, 并且由于突變位點的不同導(dǎo)致株高降低的程度存在差異, 突變體僅在第4外顯子上發(fā)生1處堿基替換, 植株矮化程度較輕, 且并未出現(xiàn)雌穗和雄穗發(fā)育缺失, 能夠正常生長。RNA結(jié)合蛋白通過在轉(zhuǎn)錄水平上改變與之結(jié)合RNA的命運或功能, 調(diào)控植物的生長發(fā)育[35]。RRM是特征最明確的RNA結(jié)合基序, 其拓?fù)浣Y(jié)構(gòu)為βαββαβ, 與RNA的特異性識別通常發(fā)生在β-sheet的表面[36]。編碼含有3個RRM結(jié)構(gòu)域的RNA結(jié)合蛋白, 但該基因下游的靶標(biāo)RNA尚未找到。突變體中發(fā)生突變的天冬氨酸殘基具有較高的保守性, 且出現(xiàn)在保守性較高的RRM3結(jié)構(gòu)域內(nèi), 位于與RNA的特異性識別相關(guān)的β-sheet的表面, 因此我們推測中的點突變可能通過影響與下游RNA的結(jié)合能力進而影響其他基因的表達(dá)。由此可見,突變體的發(fā)現(xiàn)對于從RRM3結(jié)構(gòu)域出發(fā)探索下游靶標(biāo)RNA, 以及解析3個RRM在多葉矮化表型產(chǎn)生過程中所扮演的角色具有重要研究意義。
4 結(jié)論
本研究發(fā)現(xiàn)了一個玉米多葉矮化突變體, 其株高較野生型KN5585顯著下降41.79%, 葉片數(shù)較野生型KN5585顯著增加56.18%。該突變表型受一對隱性核基因控制, 被定位在3號染色體分子標(biāo)記Indel10和Indel11之間, 物理距離0.74 Mb。進一步對定位區(qū)間內(nèi)候選基因測序, 發(fā)現(xiàn)第4外顯子出現(xiàn)1個堿基替換, 其他基因無差異, 因此, 將確定為突變表型的關(guān)鍵候選基因, 該基因編碼一個含有3個RNA結(jié)合結(jié)構(gòu)域的RNA結(jié)合蛋白, 突變位點位于第3個RNA結(jié)合結(jié)構(gòu)域。突變體的發(fā)現(xiàn)為探究玉米葉片數(shù)目(節(jié)間數(shù)目)與節(jié)間長度的平衡以及二者對株型發(fā)育的影響提供了寶貴的試驗材料。
[1] Tester M, Langridge P. Breeding technologies to increase crop production in a changing world., 2010, 327: 818–822.
[2] Haarhoff S J, Swanepoel P A. Plant population and maize grain yield: a global systematic review of rainfed trials., 2018, 5: 1819–1829.
[3] Bensen R J, Johal G S. Cloning and characterization of the maizegene., 1995, 7: 75–84.
[4] Chen Y, Hou M M, Liu L J, Wu S, Shen Y, Ishiyama K, Kobaya, Shi M, McCarty D R, Tan B C. The maizeencodes a gibberellin 3-oxidase and is dual localized to the nucleus and cytosol., 2014, 166: 2028–2039.
[5] Teng F, Zhai L H, Liu R X, Bai W, Wang L Q, Huo D G, Tao Y S, Zheng Y L, Zhang Z X., a candidate gene for a major QTL,, for plant height in maize., 2013, 73: 405–416.
[6] Winkler R G, Helentjaris T. The maizegene encodes a cytochrome P450-mediated early step in gibberellin biosynthesis., 1995, 7: 1307–1317.
[7] Cassani E, Bertolini E, Cerino Badone F, Landoni M, Gavina D, Sirizzotti A, Pilu R. Characterization of the first dominant dwarf maize mutant carrying a single amino acid insertion in the VHYNP domain of thegene., 2009, 24: 375–385.
[8] Harberd N P, Freeling M. Genetics of dominant gibberellin- insensitive dwarfism in maize., 1989, 121: 827–838.
[9] Lawit S J, Wych H M, Xu D, Kundu S, Tomes D T. Maize DELLA proteinsandas modulators of plant development., 2010, 51: 1854–1868.
[10] Multani D S, Briggs S P, Chamberlin M A, Blakeslee J J, Murphy A S, Johal G S. Loss of an MDR transporter in compact stalks of maizeand sorghummutants., 2003, 302: 81–84.
[11] Zhang X, Hou X, Liu Y, Zheng L, Yi Q, Zhang H, Huang X, Zhang J, Hu Y, Yu G, Liu H, Li Y, Huang H, Zhan F, Chen L, Tang J, Huang Y. Maize brachytic2 () suppresses the elongation of lower internodes for excessive auxin accumulation in the intercalary meristem region., 2019, 19: 589.
[12] Hartwig T, Chuck G S, Fujioka S, Klempien A, Weizbauer R, Potluri D P V, Choe S, Johal G S, Schulz B. Brassinosteroid control of sex determination in maize., 2011, 108: 19814–19819.
[13] Best N B, Hartwig T, Budka J, Fujioka S, Johal G, Schulz B, Dilkes B P.encodes a maize ortholog of thebrassinosteroid biosynthesis gened, identifying developmental interactions between brassinosteroids and gibberellins., 2016, 171: 2633–2647.
[14] Makarevitch I, Thompson A, Muehlbauer G J, Springer N M.gene in maize encodes a brassinosteroid C-6 oxidase., 2012, 7: e30798.
[15] Kir G, Ye H, Nelissen H, Neelakandan A K, Kusnandar A S, Luo A, Inzé D, Sylvester A W, Yin Y, Becraft P W. RNA interference knock down ofin maize reveals novel functions for brassinosteroid signaling in controlling plant architecture., 2015, 169: 826–839.
[16] Li H, Wang L, Liu M, Dong Z, Li Q, Fei S, Xiang H, Liu B, Jin W. Maize plant architecture is regulated by the ethylene biosynthetic gene., 2020, 183: 1184–1199.
[17] Schaller G E, Bishopp A, Kieber J J. The Yin-Yang of hormones: cytokinin and auxin interactions in plant development., 2015, 27: 44–63.
[18] Phillips K A, Skirpan A L, Liu X, Christensen A, Slewinski T L, Hudson C, Barazesh S, Cohen J D, Malcomber S, Mcsteen P.encodes a grass-specific tryptophan aminotransferase required for vegetative and reproductive development in maize., 2011, 23: 550–566.
[19] Lee B H, Johnston R, Yang Y, Gallavotti A, Kojima M, Traven?olo B A, Costa Lda F, Sakakibara H, Jackson D. Studies ofmutants of maize indicate complex interactions between auxin and cytokinin signaling in the shoot apical meristem., 2009, 150: 205–216.
[20] 張在寶, 李婉杰, 李九麗, 張弛, 胡夢輝, 程琳, 袁紅雨. 植物RNA結(jié)合蛋白研究進展. 中國農(nóng)業(yè)科學(xué), 2018, 51: 4007–4019. Zhang Z B, Li W J, Li J L, Zhang C, Hu M H, Cheng L, Yuan H Y. The research progress of plant RNA binding proteins., 2018, 51: 4007–4019 (in Chinese with English abstract).
[21] Cho H, Cho H S, Hwang I. Emerging roles of RNA-binding proteins in plant development., 2019, 51: 51–57.
[22] Jeffares D C, Phillips M J, Moore S, Veit B. A description of the Mei2-like protein family; structure, phylogenetic distribution and biological context., 2004, 214: 149–158.
[23] Kawakatsu T, Itoh J, Miyoshi K, Kurata N, Alvarez N, Veit B, Nagato Y.regulates leaf initiation and maturation in rice., 2006, 18: 612–625.
[24] Anderson G H, Alvarez N D, Gilman C, Jeffares D C, Trainor V C, Hanson M R, Veit B. Diversification of genes encoding mei2–like RNA binding proteins in plants.,2004, 54: 653–670.
[25] 王關(guān)林, 方宏筠. 植物基因工程(第2版). 北京: 科學(xué)出版社, 2002. pp 742–744. Wang G L, Fang H Y. Plant Gene Engineering, 2nd edn. Beijing: Science Press, 2002. pp 742–744 (in Chinese).
[26] Michelmore R W, Paran I, Kesseli R V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations., 1991, 88: 9828–9832.
[27] Avila L M, Cerrudo D, Swanton C, Lukens L., a putative inositol polyphosphate 5-phosphatase, is required for internode elongation in maize., 2016, 67: 1577–1588.
[28] Zhang D, Sun W, Singh R, Zheng Y, Cao Z, Li M, Lunde C, Hake S, Zhang Z.regulates shoot architecture and meristem determinacy in maize., 2018, 30: 360–374.
[29] Li W, Ge F, Qiang Z, Zhu L, Zhang S, Chen L, Wang X, Li J, Fu Y. Maizeencodes a microtubule-associated protein that controls plant and ear height., 2020, 18: 1345–1347.
[30] Heuer S, Hansen S, Bantin J, Brettschneider R, Kranz E, L?rz H, Dresselhaus T. The maize MADS box geneaffects node number and spikelet development and is co-expressed withduring flower development, in egg cells, and early embryogenesis., 2001, 127: 33–45.
[31] Lyu H K, Zheng J, Wang T Y, Fu J J, Huai J L, Min H W, Zhang X, Tian B H, Shi Y S, Wang G Y. The maize, a novel allele of, is required for maize internode elongation., 2014, 84: 243–257.
[32] Bommert P, Je B I, Goldshmidt A, Jackson D. The maize Gα genefunctions in clavata signalling to control shoot meristem size., 2013, 502: 555–558.
[33] Veit B, Briggs S P. Regulation of leaf initiation by thegene of maize., 1998, 393: 166–168.
[34] Wang F, Yu Z, Zhang M, Wang M, Lu X, Liu X, Li Y, Zhang X, Tan B C, Li C, Ding Z.promotes plant height by regulating intercalary meristem formation and internode cell elongation in maize., 2022, 20: 526–537.
[35] Hentze M W, Castello A, Schwarzl T, Preiss T. A brave new world of RNA-binding proteins., 2018, 19: 327–341.
[36] 唐蜻. 植物RNA結(jié)合蛋白的研究進展. 安徽農(nóng)業(yè)科學(xué), 2010, 38(1): 38–41. Tang Q. The research progress of plant RNA binding proteins., 2010, 38(1): 38–41 (in Chinese with English abstract).
Identification and gene cloning of leafy dwarf mutantin maize
SU Shuai, LIU Xiao-Wei, NIU Qun-Kai, SHI Zi-Wen, HOU Yu-Wei, FENG Kai-Jie, RONG Ting-Zhao, and CAO Mo-Ju*
Maize Research Institute, Sichuan Agricultural University / Key Laboratory of Biology and Genetic Improvement of Maize in Southwest Region, Ministry of Agriculture and Rural Affairs, Chengdu 611130, Sichuan, China
The decrease of plant height in maize is usually caused by the decrease in the number of internodes, the shortening of internodes or the combination of both. However, in this study, the mutant() found in the progeny of gene editing, exhibited more leaves and shorter stature. Quantitative measurements indicated the plant height of mutantwas only 93.10 cm, the plant height of wild-type KN5585 was 159.95 cm. The plant height was significantly reduced by 41.79% in mutantcompared with the wild type KN5585. The wild type KN5585 produced an average of 17.8 leaves at maturity stage, whereas mutantsproduced 27.8 leaves. The number of leaves were significantly increased by 56.18% in mutantcompared with the wild type. Genetic analysis showed that the mutation phenotype ofwas controlled by a pair of recessive nuclear genes. We applied a map-based cloning strategy to identify the gene responsible for thephenotype. The gene was located between Indel10 and Indel11 on maize chromosome 3, and the physical distance was 0.74 Mb. Gene sequencing analysis of 13 genes (excluding pseudogenes) within the interval revealed that one base A was substituted in the fourth exon of, and there was no significant difference in other genes.encoded an RNA-binding protein. The amino acid substitution was in the third RNA binding domain (RRM3), resulting in the conversion of aspartic acid to valine. The mutation sites of the mutantwere different from,,, andin previously reported. The discovery ofprovides valuable materials for further analysis of the genetic mechanism of the balance between leaves and internodes development in maize.
maize; the number of leaves; the length of internodes; gene mapping
10.3724/SP.J.1006.2024.33044
本研究由四川省科技計劃項目(2021YFYZ0011, 2021YFYZ0017, MZGC20230108)和四川農(nóng)業(yè)大學(xué)學(xué)科建設(shè)專項研究支持計劃項目資助。
This study was supported by the Sichuan Science and Technology Program (2021YFYZ0011, 2021YFYZ0017, MZGC20230108) and the Specific Research Supporting Program for Discipline Construction at Sichuan Agricultural University.
曹墨菊, E-mail: caomj@sicau.edu.cn
E-mail: 1018714902@qq.com
2023-07-31;
2024-01-12;
2024-02-08.
URL: https://link.cnki.net/urlid/11.1809.S.20240206.1114.002
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
