陸地棉重組自交系再生能力和遺傳轉化效率篩選
2024-04-28 01:17:44張獻龍林忠旭
作物學報 2024年5期
關鍵詞:體系
樂 愉 王 濤 張獻龍 林忠旭
陸地棉重組自交系再生能力和遺傳轉化效率篩選
樂 愉 王 濤 張獻龍 林忠旭*
華中農業大學植物科學技術學院 / 作物遺傳改良全國重點實驗室, 湖北武漢 430070
轉基因工程育種是棉花種質創新的有效手段, 為了拓展陸地棉可再生基因型, 豐富棉花轉基因受體, 本研究以湖北省高產闊葉棉品種‘鄂棉22’ (E22)為母本、高再生能力的雞腳葉品種‘豫早1號’ (YZ1)為父本, 通過單籽傳法構建了F9重組自交系群體(YE); 利用棉花中比較成熟的IBA+KT (IK)和2,4-D+KT (DK) 2種不同激素組合的培養體系, 分別對重組自交系群體的164個家系的愈傷組織誘導率(CIF)、愈傷組織繼代繁殖力(CSC)、愈傷組織的出胚率(CRE)以及愈傷組織出胚時間(CET)進行比較, 共獲得12個可再生的闊葉棉家系; 利用目前成熟的棉花遺傳轉化體系對可再生闊葉棉家系進行遺傳轉化效率分析, 同時對其進行田間農藝性狀考察, 最終獲得了一個遺傳轉化效率為82.9%并且農藝性狀更優良的家系YE3。本研究為棉花的遺傳轉化及基因功能研究拓展了陸地棉基因型種質資源。
棉花; 重組自交系; 體細胞胚胎發生; 再生能力; 遺傳轉化效率
棉花是世界上重要的纖維和油料作物, 在我國的經濟發展中有著舉足輕重的地位。傳統的雜交育種法已經難以滿足現代棉花遺傳改良的需要, 轉基因生物技術育種是棉花種質創新的熱點[1]。而現代分子生物技術應用的先決條件在于具備不同品種的組織培養和植株再生的高效體系, 因此在棉花的遺傳改良過程中, 豐富的棉花遺傳轉化受體材料以及較高的再生能力和遺傳轉化效率至關重要。
棉花組織培養體系的構建起步較晚, 但發展迅速。1979年, Price和Smith利用懸浮培養技術成功地誘導出了棉花體細胞胚[2], 1983年, Davidonis對陸地棉Coker 310 (珂字棉)經過2年多的繼代培養, 成功獲得第1株棉花再生植株[3]。歷經十幾年的研究探索, 棉花組織培養體系逐漸完善, 隨后進入快速發展時期, 由于棉花體細胞胚胎發生受基因型限制, 很多體細胞胚胎發生的陸地棉多為珂字棉或有其遺傳背景的品種[4-6]。2006年, 金雙俠等[7]通過對多種基因型棉花材料進行離體培養, 篩選到了2個體細胞胚胎發生能力明顯高于珂字棉的基因型材料‘豫668’ (Y668)和‘豫早1號’ (YZ1); 2008年, 陳天子等[8]利用新疆4個主栽棉花品種‘新陸中20’、‘新陸早24’、‘新陸早33’和‘03298’為材料, 通過不同濃度的激素組合成功地誘導體細胞胚發生并且在6~8個月獲得大批量再生苗。葛書婭等[9]設計了6種不同激素組合的培養體系, 成功建立了新疆推廣的陸地棉品種‘新陸早45號’的再生體系。對棉花不同品種的再生條件的摸索, 拓展了陸地棉再生基因型, 為棉花的遺傳轉化豐富了受體材料。
植物轉基因技術已經成為植物分子育種中最常用的手段, 棉花轉基因后能否快速地獲得再生植株, 除了受到組織培養再生體系的影響, 關鍵影響因素是轉基因受體的可再生能力[10]。再生能力受基因型決定, 不同基因型間再生能力差異很大, 且多為數量性狀[11]。張獻龍等[12]提出棉花的基因型是影響體細胞胚胎發生和植株再生的首要因素, 而其他外界條件是次要的。通過對4個栽培品種的比較發現, 再生能力在栽培種之間也存在較大的差異: 陸地棉>亞洲棉>海島棉>非洲棉, 而同一栽培種的不同棉花品種間也存在差異, 例如陸地棉中以珂字棉的再生能力最強, 珂字棉>岱字棉>斯字棉[13]。2022年, Ge等[14]以棉花種子的頂端分生組織干細胞為外植體, 利用農桿菌和超聲波處理將外源基因整合到干細胞中, 通過誘導干細胞產生不定芽進而發育成再生植株, 建立了無基因型限制的棉花遺傳轉化體系。然而, 通過體細胞胚胎發生再生植株仍然是棉花轉基因研究的重要方法。通過對再生能力相關的基因進行定位和克隆, 用以改造農藝性狀優良但再生能力低的基因型, 是實現作物轉基因生產的有效手段[15]。由于棉花體細胞胚胎發生和植株再生對基因型的依賴性較大, 根據已報道的不同棉花品種的再生培養體系, 選擇與親本相關的再生體系對構建的群體進行培養, 通過對再生能力相關性狀的數據統計, 嘗試定位與體細胞胚發生和再生能力相關的位點, 然后有針對性的將這些位點導入到農藝性狀優良的品種中去, 可用于進一步改良這些優良品種[16]。
為了拓展陸地棉可再生的基因型, 豐富陸地棉的轉基因受體材料, 本研究以湖北省高產棉花品種‘鄂棉22’ (Emian 22, E22)和高再生能力的品種‘豫早1號’ (Yuzao 1, YZ1)為親本, 構建了包含164個家系的重組自交系群體(YE), 采用IBA+KT (IK)和2,4-D+KT (DK) 2種培養體系對YE家系的再生能力進行鑒定并對可再生闊葉棉家系進行遺傳轉化效率分析, 以期篩選出農藝性狀優良且遺傳轉化效率較高的材料。
1 材料與方法
1.1 試驗材料
以‘鄂棉22’ (E22)為母本、‘豫早1號’ (YZ1)為父本雜交獲得F1。E22是由湖北省黃岡市農業科學院選育, 產量較高, 纖維品質良好, 但再生能力較差, 難以獲得再生植株。YZ1再生能力高[7,17], 可在短時間內獲得再生植株, 但產量低, 纖維品質一般。F1自交獲得F2, 之后通過單籽傳法自交9代獲得純合株系, 該重組自交系群體包含164個株系, 以YE表示。
1.2 試驗方法
1.2.1 組織培養方法 基于本實驗室優化的棉花組織培養體系[18], 采用IBA+KT (IK)和2,4-D+KT (DK) 2套培養體系, 分別對164個YE家系及親本E22和YZ1的下胚軸進行愈傷組織的誘導、胚性愈傷組織的分化和再生培養。
1.2.2 培養基的選擇 本試驗中所用的培養體系是經過多次優化后的、適合棉花組織培養的培養體系, 包括無菌苗萌發培養基、愈傷組織誘導培養基、體細胞胚分化培養基、生根培養基。采用IK和DK 2套體系進行培養。
無菌苗萌發培養基: 1/2 MS大量元素, 附加15 g L–1葡萄糖, 2.5 g L–1的Phytagel。
愈傷組織誘導培養基: MS無機鹽, B5維生素作為基本培養基(以下簡稱MSB), 附加不同種類的激素組合。
體細胞胚分化培養基: MSB基本培養基(KNO3加倍, 去掉NH4NO3)并附加Gln 2.0 g L–1和Asn 1.0 g L–1。
生根培養基: 1/2 MS無機鹽加B5有機物, 附加15 g L–1葡萄糖, 2.5 g L–1的Phytagel。
所有的激素和氨基酸在滅菌前加入, 在121℃, 1.1 kg cm–2壓力下高溫高壓滅菌15 min。所有培養物均置于(28±2)℃, 光照強度(冷光源135mmol m–2s–1), 每天光照14 h的條件下進行培養。
1.2.3 愈傷組織的誘導 無菌苗的培養: 硫酸脫掉短絨后的種子剝掉種皮后浸泡于10%的升汞溶液8~10 min, 再用無菌水沖洗3~5次, 接種于無菌苗培養基上, 露白之后將根用無菌的鑷子插入培養基中, 并剝去內種皮, 置于20℃的暗培養條件下生長。
愈傷組織誘導: 取無菌苗下胚軸切成0.5~ 0.8 cm小段分別接種于2種愈傷組織誘導培養基上, 每個家系培養3瓶, 每瓶初始放置10段下胚軸, 并稱取0 d的重量; 放置于光照培養室進行培養, 培養每隔25 d后采用各自所屬體系進行繼代培養, 統計愈傷組織的誘導率(callus induction rate, CIF); 第1次25 d繼代時稱取每個家系的增殖重量, 求取平均值, 計算愈傷組織繼代繁殖力(callus secondary fecundity, CSC)。
1.2.4 愈傷組織的繼代 隨著愈傷組織的快速增長, 培養基空間過于擁擠, 所以在繼代過程中隨機減少愈傷的塊數, 每瓶培養基只保留6塊愈傷組織, 在繼代過程中統計愈傷組織最早出胚時間(callus embryo time, CET)和愈傷組織的出胚塊數以及瓶數, 從而計算愈傷組織的出胚率(embryo rate of callus, CRE)。
1.2.5 愈傷組織的分化 觀察到分化出胚性愈傷組織后立即轉入分化培養基中培養, 繼續繼代培養直到分化成胚, 再選擇發育正常的子葉胚轉移到生根培養基上, 促使幼苗生根。
1.2.6 衡量棉花再生能力性狀的統計 本次試驗所統計的與棉花體細胞再生能力的性狀有: 愈傷組織誘導率(CIF)、愈傷組織繼代繁殖力(CSC)、愈傷組織最早出胚時間(CET)、愈傷組織的出胚率(CRE) 4個指標, 計算方法如下: 愈傷組織誘導率(CIF): 出愈傷塊數/誘導愈傷總塊數×100; 愈傷組織繼代繁殖力(CSC): 繼代25 d后的愈傷重/繼代0 d的愈傷重×100; 愈傷組織出胚時間(CET): 單個家系中最早出現胚性愈傷的時間; 愈傷組織的出胚率(CRE): 出現胚性愈傷的塊數/繼代總塊數×100。
1.2.7 遺傳轉化效率分析 以12個可再生闊葉棉家系為轉化受體材料, 每一個受體保證10瓶無菌苗, 每瓶4粒種子的下胚軸用于切苗侵染; 在農桿菌侵染后的第1個月, 每個愈傷組織誘導培養皿有35~40段生長健康的下胚軸段, 每個受體獲得10~12個培養皿不等, 不同的培養皿是不同的重復。采用IK培養體系, 以紅色熒光蛋白(red fluorescence protein, RFP)為報告基因, 紅色熒光蛋白由華中農業大學金雙俠教授提供, 利用農桿菌介導的棉花遺傳轉化方法進行棉花遺傳轉化[18]; 再利用體式顯微鏡在紅色熒光波長下觀察轉基因植株是否有紅色熒光, 遺傳轉化效率計算方法為陽性單株/總轉化單株×100。
1.2.8 農藝性狀考察 12個可再生闊葉棉家系以及親本E22和YZ1于2019年種植于湖北武漢、2020年及2021年種植于湖北鄂州試驗基地。待棉花完全吐絮時, 收取植株中部吐絮正常的棉花, 軋花后對衣分含量以及棉花纖維品質性狀進行考察。
2 結果與分析
2.1 2種培養體系下YE群體的再生能力鑒定
2.1.1 體細胞胚胎發生的差異 不同的棉花品種在同一培養體系下, 其再生時間存在明顯的差別, 而同一品種選擇的培養體系不同, 再生能力也存在差異。因此, 針對高再生能力的家系選擇合適的培養體系就顯得至關重要。
棉花的植株再生過程通常包括外植體接種、愈傷組織的誘導、胚性愈傷組織的分化、胚狀體的形成和植株再生4個過程。胚性愈傷組織轉移到分化培養基上培養2~3個月后會得到大量胚狀體, 對胚狀體進行誘導后得到再生幼苗。但是在利用IK和DK體系培養YE家系的過程中發現, 在IK培養體系下(圖1)有個別家系(YE3)在2~3個月的時候未見大量胚性愈傷, 卻率先發現了零星的子葉胚(圖1-C),通過對子葉胚的誘導, 能夠順利得到再生植株, 可在原來培養周期的基礎上進一步縮短再生周期。這些家系在DK培養體系(圖2)下并未出現這種先出現子葉胚的情況, 因此在培養體系的選擇上, IK體系比DK體系更適合YE群體。
2.1.2 再生能力的比較 利用IK和DK培養體系對164個YE家系和親本進行組織培養, 本研究對CIF、CSC、CET、CRE 4個指標進行了數據統計。由于培養體系的優化, 在2種培養體系下所有的家系都可以誘導出愈傷組織, 因此所有家系的CIF都接近100%, 本研究可通過其他3個指標來衡量不同家系的再生能力。

圖1 IK體系下棉花體細胞胚胎發生和植株再生流程
A: 棉花下胚軸; B: 1~2個月的愈傷組織; C: 胚性愈傷組織, 伴有子葉型胚狀體; D: 大量胚狀體形成; E: 植株再生; F: 水培后可移栽。
A: hypocotyl; B: the induced callus tissue of 1–2 months; C: the embryonic callus with cotyledons; D: a large amount of embryoid formation; E: plant regeneration; F: hydroponics can be transplanted.

圖2 DK體系下棉花體細胞胚胎發生和植株再生流程
A: 棉花下胚軸; B: 1~2個月的愈傷組織; C: 胚性愈傷組織, 無胚狀體; D: 大量胚狀體形成; E: 植株再生; F: 水培后可移栽。
A: hypocotyl; B: the induced callus tissue of 1–2 months; C: the embryonic callus with cotyledons; D: a large amount of embryoid formation; E: plant regeneration; F: hydroponics can be transplanted.
由IK和DK體系下愈傷組織增殖頻率分布圖(圖3)可知, YE家系在DK培養體系下的繼代繁殖力大部分在4~7 g, 而在IK培養體系下的繼代繁殖力大部分在3~5 g, 在DK培養體系下的繼代繁殖力高于IK體系。對于在IK和DK 2種培養體系下均可出胚的家系, 本研究對其相關性狀的統計分析發現, 對于出胚時間(圖4-A), 在IK體系下, 出胚時間主要集中在60~80 d, 而在DK體系下, 出胚時間主要集中在80~100 d, IK體系的出胚時間要比DK體系的出胚時間短; 對胚性愈傷組織出胚率(圖4-B)的比較可以看出, 出胚率大多處于60%~100%, 2種體系沒有明顯差異。綜合胚性愈傷組織增殖、愈傷組織出胚時間和胚性愈傷組織出胚率可知, 過度地愈傷增殖并不會加快胚性愈傷組織的出現, 對出胚率也沒有明顯影響。
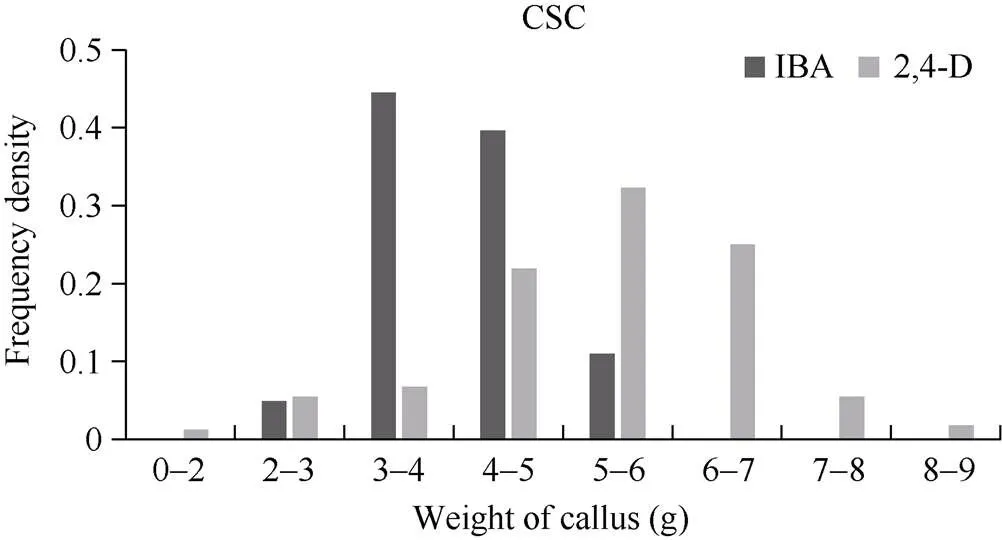
圖3 IK和DK體系下愈傷組織增殖的頻率分布圖
CSC: 愈傷組織繼代繁殖力。CSC: callus secondary fecundity.
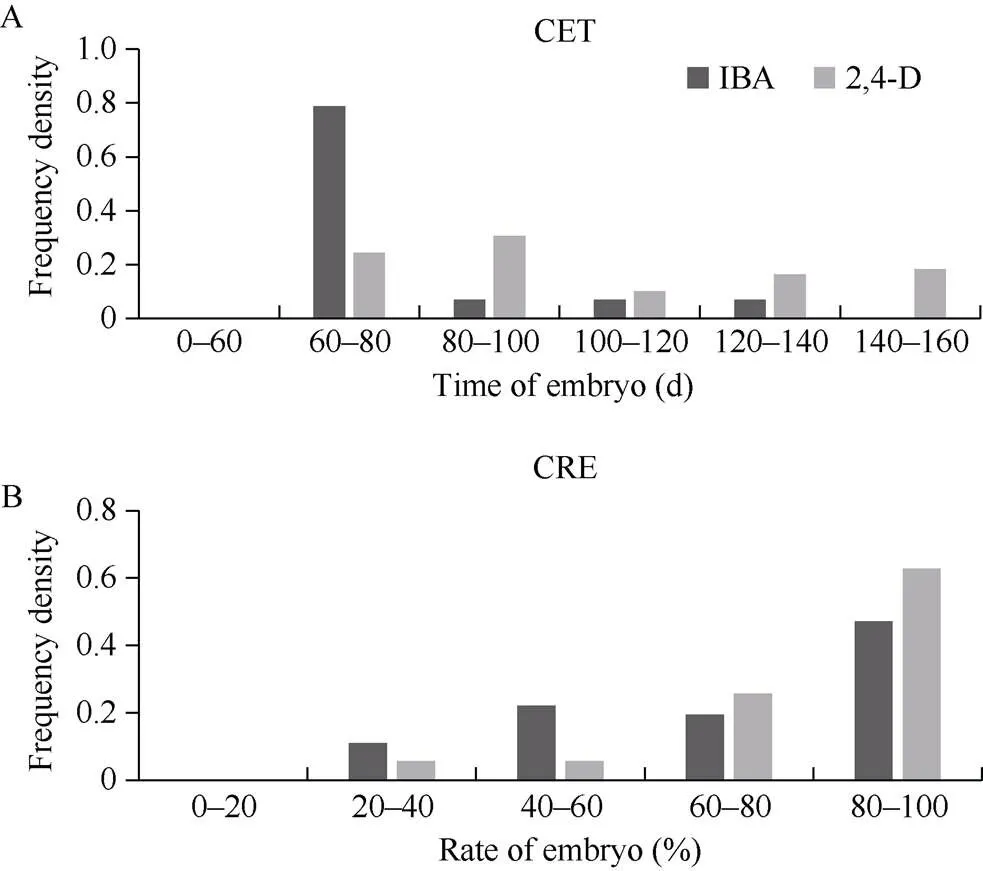
圖4 IK和DK體系下可出胚家系再生相關性狀的頻率分布圖
A: CET, 愈傷組織最早出現的時間; B: CRE, 愈傷組織出胚率。
A: CET, callus embryo time; B: CRE, the embryo rate of callus.
2.1.3 YE家系再生植株統計 本研究對在2種體系下獲得的再生苗進行統計發現, 共有36個家系在IK體系下獲得再生苗, 其中22個是闊葉棉家系; 共有35個家系在DK體系下獲得再生苗, 其中19個是闊葉棉家系; 共有25個家系在2個培養體系下均能獲得再生苗, 其中闊葉棉家系有12個(表1), 圖5所示為部分再生植株。本研究選擇在2個體系下均可以獲得再生苗的闊葉棉家系繼續后續的遺傳轉化分析, 這些闊葉棉家系在IK體系下的CIF為100%, CSC不低于2.8 g, CER不低于50%, CET不晚于120 d。

圖5 部分再生植株
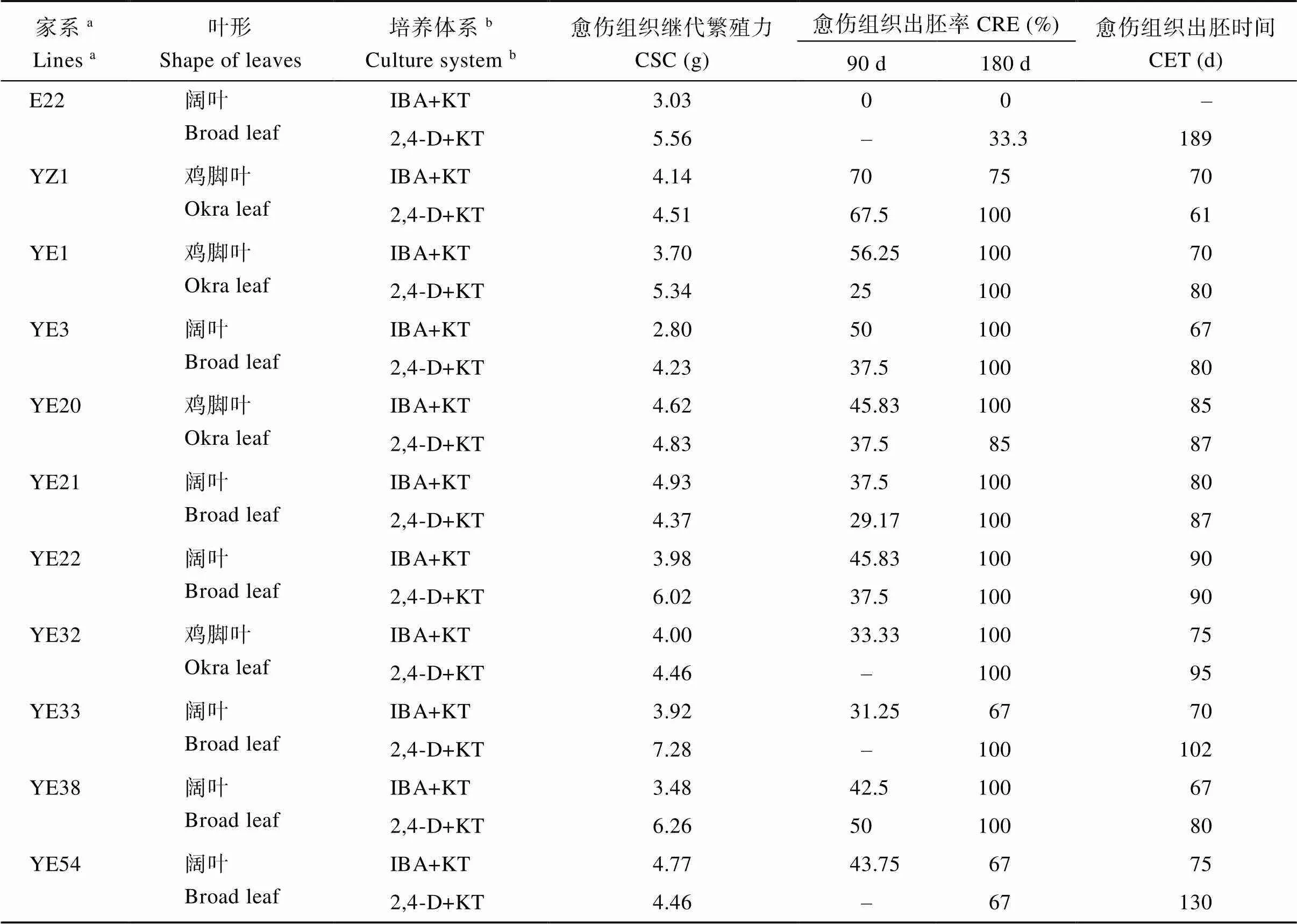
表1 再生YE家系CSC、CRE和CET比較

(續表1)
aYE表示以E22和YZ1為親本獲得的重組自交系群體的家系編號;bIBA+KT表示吲哚丁酸+激動素激素組合, 2,4-D+KT表示2,4-二氯苯氧乙酸+激動素激素組合。
aYE: the recombinant inbred lines obtained with E22 and YZ1 as the parents.bIBA+KT: hormone combination of indole butyric acid + kinetin, 2,4-D+KT: hormone combination of 2,4-dichlorophenoxyacetic acid + kinetin. CSC: callus secondary fecundity; CRE: the embryo rate of callus; CET: callus embryo time.
2.2 可再生闊葉棉遺傳轉化效率鑒定
本研究對篩選得到的所有可再生闊葉棉家系利用IK體系進行遺傳轉化發現, 并不是所有的可再生闊葉棉家系在經過農桿菌侵染后都能獲得再生植株。YE81和YE86在侵染90 d后開始出現大面積的農桿菌污染, 下胚軸壞死; YE21無法誘導出愈傷組織, 下胚軸發黑壞死; YE22、YE33、YE38、YE111、YE138和YE142可以誘導出愈傷組織, 但是后面無法誘導出胚性愈傷組織; YE87可以誘導出胚性愈傷組織, 但是胚狀體發育畸形, 無法獲得再生植株; 只有YE3和YE54能完成體細胞胚胎發生及植株再生過程, YE3在培養7個月后開始獲得再生苗, YE54只獲得了陰性單株, 沒有獲得陽性單株。圖6中的ABCD和EFGH分別為在白光和紅色熒光條件下YE3轉基因胚性愈傷組織(圖6-A)、體細胞胚(圖6-B)和轉基因植株葉片(圖6-C, D), 其中圖6-D和6-F所示左邊為陽性植株葉片, 右邊為陰性植株葉片。最終統計表明以YE3為受體獲得的轉基因總植株數為41, 陽性單株數為34, 其遺傳轉化效率達到82.9%。

圖6 YE3轉基因植株陽性鑒定
A和E: 胚性愈傷組織; B和F: 體細胞胚; C、G、D和F: 葉片; 標尺為1000mm。
A and E: embryonic callus; B and F: embryo; C, G, D, and H: leaf. Bar: 1000mm.
2.3 可再生闊葉棉家系農藝性狀考察
本研究考察了12個可再生的闊葉棉家系的纖維品質及纖維產量相關性狀, 包括纖維長度、纖維強度、馬克隆值、整齊度、伸長率和衣分。由表2可知, 在纖維長度方面, 除YE22明顯低于YZ1外, 其他可再生闊葉棉家系均接近或高于YZ1; 在纖維強度方面, 所有家系均高于YZ1; 馬克隆值和伸長率在E22和YZ1之間沒有差異, 可再生闊葉家系中的馬克隆值和伸長率變化較大; YZ1的整齊度高于E22, 可再生闊葉棉家系YE3、YE21、YE81、YE138和YE142的整齊度均高于YZ1; 在產量性狀衣分上, 除了YE81和E22接近, 其他闊葉棉家系均高于YZ1。由此可見, 可再生闊葉棉中大部分家系表現出優于高再生能力材料YZ1的農藝性狀, 說明這些可再生闊葉家系中同時還攜帶了與優良農藝性狀相關的遺傳信息。

表2 12個可再生闊葉家系農藝性狀考察
aYE表示以E22和YZ1為親本獲得的重組自交系群體的家系編號。
aYE represents the recombinant inbred lines obtained with E22 and YZ1 as the parents.
YE3在纖維品質性狀如纖維長度、纖維強度、纖維整齊度、纖維伸長率方面要明顯優于高再生能力的材料YZ1; 在產量性狀上, YE3的衣分為40.53%, 與E22的43.72%接近, 遠遠高于YZ1的35.43%。綜上, 在主要農藝性狀方面, YE3表現出優于高再生材料YZ1的潛力, 表明YE3中攜帶了YZ1高再生能力相關的遺傳位點, 同時又保留了高產陸地棉品種的優異農藝性狀。
3 討論
3.1 高再生能力家系的篩選
同一個家系在不同培養體系下的再生能力存在差異。李雪林等[19]利用DK和IK體系對多個棉花品種的體細胞胚胎發生能力進行比較時發現, DK體系下大鈴棉和邯8959兩個材料只能分化出胚性愈傷, 但在IK體系下胚性愈傷組織能夠萌發出胚狀體, 并再生出完整植株。本研究利用DK和IK體系對YE家系的再生能力統計過程中也發現, 有些家系在2個體系下都可再生(表1); 但也有一部分家系只在其中一個體系下可再生, 如YE16、YE34和YE129等10個家系只能在DK體系下獲得再生植株, YE30、YE44、YE80等11個家系只能在IK體系下可以獲得再生植株。本研究認為在2個體系下都能獲得再生植株的YE家系才是高再生能力的代表, 至少說明其基因型響應2種激素的能力較強。
3.2 高遺傳轉化效率及農藝性狀優良材料的篩選
篩選再生能力強以及遺傳轉化效率高且綜合農藝性狀優良的闊葉棉材料對于棉花轉基因工程尤為重要。YZ1雖然具有較高的再生能力, 但以它作為轉基因受體的弊端很明顯, 作為超雞腳葉的典型代表, YZ1的纖維產量和品質不佳, 以它為受體的轉基因植株很難滿足當前的生產需要。E22適應范圍廣, 其纖維產量和纖維品質優于YZ1, 但該材料無法通過組織培養技術誘導體細胞胚胎的發生及植株再生,因此我們無法以它作為轉基因受體材料進行定向改良研究。為了獲得高再生能力并且產量和品質優良的材料, 以滿足當前生產上的遺傳改良需要, 本研究以YZ1和E22為親本, 通過多代自交獲得了穩定遺傳的重組自交系群體, 避免了像F2群體只能用一代的弊端, 可以常年獲得外植體對其進行組織培養和遺傳轉化研究。
作為維管束植物最主要的營養器官, 葉片對于光合產物的積累、養分分配以及水分運輸起著重要作用[20]; 葉形和空間分布會影響光能利用效率, 在很大程度上決定著作物的產量和品質[21]。目前生產上主栽陸地棉品種為闊葉棉, 闊葉棉葉面積大, 光合作用強; 雞腳葉葉面積小, 不利于光合作用, 并且與纖維產量和品質不利性狀關聯, 不適于大面積種植[22]。因此, 本研究選擇可再生的闊葉棉家系進行遺傳轉化效率的研究。
本研究共獲得了12個在IK和DK 2種培養體系下均可再生的闊葉棉家系, 選用更適合于YE家系的IK培養體系進行培養, 利用農桿菌介導法進行遺傳轉化, 農桿菌介導外源基因進入植物基因組需要編碼毒力蛋白(vir)的基因, 這些毒蛋白基因的表達受到受傷的植物釋放的酚類物質的誘導[23]。農桿菌介導棉花遺傳轉化受到多因素影響[24], 內在因素包括基因型和外植體, 外在因素包括培養體系、農桿菌菌株和濃度以及浸染時間、酚類物質乙酰丁香酮(AS)的濃度、共培養的時間以及抗生素的濃度等, 本研究在對可再生闊葉棉家系進行遺傳轉化時, 除了基因型的差異外, 其他因素都是保持一致的。遺傳轉化結果表明, 并不是所有能夠再生的闊葉棉家系都能夠經過農桿菌的浸染并完成轉化過程而得到轉基因植株, 可能是由于基因型差異引起, 基因型不同的受體在受到農桿菌侵染時, 傷口釋放的酚類物質可能也會不同, 從而影響對毒力蛋白的誘導和T-DNA的轉移能力, 針對不同基因型的材料, 可能需要優化遺傳轉化方案才能達到理想的結果。
本研究選擇以RFP為報告基因, 因為RFP的可視性強, 可肉眼觀察轉基因植株的顏色以篩選陽性轉化事件, 同時可利用熒光體式顯微鏡觀察以明確篩選結果。通過統計陽性轉基因植株所占比例, 篩選出了高遺傳轉化效率的材料YE3, 該材料能在胚性愈傷組織誘導階段率先誘導出子葉胚, 子葉胚進一步分化成再生苗, 這一點特性可縮短再生苗的培養周期; 田間農藝性狀考察表明YE3有更優于YZ1的優良農藝性狀。綜上, 本研究表明YE3在棉花轉基因育種中具有很強的實用性。
4 結論
本研究利用DK和IK 2種培養體系對YE重組自交系群體進行再生能力的鑒定, 篩選出了12個可再生闊葉棉; 通過遺傳轉化效率分析以及農藝性狀考察, 最終篩選到一個遺傳轉化效率高于80%、且主要農藝性狀更優良的材料YE3, 本研究拓展了陸地棉遺傳轉化的基因型, 豐富了轉基因受體材料。后續研究可以將具有YZ1等位基因的位點作為具有調控棉花再生能力的候選位點, 同時結合轉錄組分析以及功能基因組挖掘棉花再生相關候選基因, 為棉花體細胞胚胎發生及再生研究提供理論基礎。
[1] Tanveer K, Siva R V, Sadhu L. High-frequency regeneration via somatic embryogenesis of an elite recalcitrant cotton genotype (L.) and efficient-mediated transformation, 2010, 101: 323–330.
[2] Price H J, Roberta H S. Somatic embryogenesis in suspension cultures ofanderss,1979, 145: 305–307.
[3] Davidonis G H, Hamilton R H. Plant regeneration from callus tissue ofL, 1983, 32: 89–93.
[4] Kumar S, Pental D. Regeneration of Indian cotton variety MCU-5 through somatic embryogenesis, 1998, 74: 538–540.
[5] Kumria R, Sunnichan V G, Das D K, Gupta S K, Reddy V S, Bhatnagar R K, Leelavathi S. High-frequency somatic embryo production and maturation into normal plants in cotton () through metabolic stress, 2003, 21: 635–639.
[6] Rauf S, Hafeez-ur-Rahman. A study ofregeneration in relation to doses of growth regulators in hybrids of upland cotton, 2005, 83: 209–215.
[7] Jin S X, Zhang X L, Nie Y C, Guo X P, Liang S G, Zhu H G. Identification of a novel elite genotype forculture and genetic transformation of cotton, 2006, 50: 519–524.
[8] 陳天子, 吳慎杰, 李飛飛, 郭旺珍, 張天真. 新疆棉花4個主栽品種的體細胞胚胎發生及植株再生作物學報, 2008, 34: 1374–1380. Chen T Z, Wu S J, Li F F, Guo W Z, Zhang T Z.regeneration of four commercial cotton (L.) cultivars grown in Xinjiang, China., 2008, 34: 1374–1380 (in Chinese with English abstract).
[9] 葛書婭, 沈秋平, 何積明, 呂尊富, 鄭海彪, 李飛飛. 陸地棉新陸早45號體細胞胚胎發生及再生體系的建立作物學報, 2018, 30: 492–497. Ge S Y, Shen Q P, He J M, Lyu Z F, Zheng H B, Li F F. A regeneration system for cotton variety Xinluzao 45somatic embryogenesis., 2018, 30: 492–497 (in Chinese with English abstract).
[10] 羅曉麗, 姜艷麗, 肖娟麗, 武宗信, 張安紅, 王志安, 吳家和. 早熟棉體細胞胚胎發生和植株再生體系的建立西北植物學報, 2011, 31: 609–615. Luo X L, Jiang Y L, Xiao J L, Wu Z X, Zhang A H, Wang Z A, Wu J H. Establishment of somatic cell embryogenesis and plant regeneration system of early cotton., 2011, 31: 609–615 (in Chinese with English abstract).
[11] 王永芳, 刁現民. 植物再生相關基因的QTL定位及克隆應用研究進展河北農業科學, 2010, 14(11): 85–88.
Wang Y F, Diao X M. Advances of QTL localization and cloning of genes related to regeneration ability of plant tissue culture., 2010, 14(11): 85–88 (in Chinese with English abstract).
[12] 張獻龍, 孫濟中, 劉金蘭. 陸地棉體細胞胚胎發生與植株再生遺傳學報, 1991, 18: 461–467. Zhang X L, Sun J Z, Liu J L. Somatic embryogenesis and plant regeneration in upland cotton., 1991, 18: 461–467 (in Chinese with English abstract).
[13] 董合忠. 不同基因型棉花下胚軸離體培養胚狀體發生的研究萊陽農學院學報, 1991, 8(2): 97–101. Dong H Z. Cotton somatic embryogenesis of different genotypes., 1991, 8(2): 97–101 (in Chinese with English abstract).
[14] Ge X Y, Xu J T, Yang Z E, Yang X F, Wang Y, Chen Y L, Wang P, Li F G. Efficient genotype-independent cotton genetic transformation and genome editing, 2022, 65: 907–917.
[15] Bolibok H, Rakoczy-Trojanowska M. Genetic mapping of QTLs for tissue-culture response in plants, 2006, 149: 73–83.
[16] Zhao L N, Zhou H J, Lu L X, Liu L L, Li X H, Lin Y J, Yu S B. Identification of quantitative trait loci controlling rice mature seed culturability using chromosomal segment substitution lines, 2009, 28: 247–56.
[17] Jin S X, Zhang X L, Liang S G, Nie Y C, Guo X P, Huang C. Factors affecting transformation efficiency of embryogenic callus of upland cotton () with, 2005, 81: 229–237.
[18] Li J Y, Wang M J, Li Y J, Zhang Q H, Lindsey K, Daniell H, Jin S X, Zhang X L. Multi-omics analyses reveal epigenomics basis for cotton somatic embryogenesis through successive regeneration acclimation process, 2019, 17: 435–450.
[19] 李雪林, 王翠玲, 孟超敏. 高頻體細胞胚胎發生的優異棉花種質材料篩選分子植物育種, 2012, 10: 683–688. Li X L, Wang C L, Meng C M. Screening elite cotton germplasm with high frequency somatic embryogenesis., 2012, 10: 683–688 (in Chinese with English abstract).
[20] Tsukaya H. Mechanism of leaf-shape determination, 2006, 57: 477–496.
[21] Wang H F, Kong F J, Zhou C E. From genes to networks: The genetic control of leaf development, 2021, 63: 1181–1196.
[22] Zhu Q H, Zhang J, Liu D X, Stiller W, Liu D J, Zhang Z S, Llewellyn D, Wilson L. Integrated mapping and characterization of the gene underlying the okra leaf trait inL., 2016, 67: 763–774.
[23] Gelvin S B. Integration ofT-DNA into the plant genome, 2017, 51: 195–217.
[24] 李靜, 張換樣, 朱永紅, 吳慎杰, 焦改麗. 農桿菌介導棉花遺傳轉化的影響因素南方農業, 2020, 14(8): 8–12. Li J, Zhang H Y, Zhu Y H, Wu S J, Jiao G L. Influencing factors of-mediated genetic transformation in cotton., 2020, 14(8): 8–12 (in Chinese with English abstract).
Screening of regeneration capacity and genetic transformation efficiency in recombinant inbred lines ofL
LE Yu, WANG Tao, ZHANG Xian-Long, and LIN Zhong-Xu*
College of Plant Science & Technology, Huazhong Agricultural University / National Key Laboratory of Crop Genetic Improvement, Wuhan 430070, Hubei, China
Transgenic engineered breeding is an effective way of cotton germplasm innovations. In order to expand the renewable genotypes of upland cotton and to enrich cotton transgenic receptors, we constructed an F9recombinant inbred population (YE) by the single-seed descent method using the high-yielding broad leaf cotton variety Emian 22 (E22) from Hubei province as the male parent and the high regeneration-capable okra leaf variety Yuzao 1 (YZ1) as the female parent. We compared the callus induction rate (CIF), callus secondary fecundity (CSC), embryo rate of callus (CRE), and callus embryo time (CET) of 164 recombinant inbred lines using two different hormone combinations of IBA+KT (IK) and 2,4-D+KT (DK), respectively, which were mature tissue culture systems in cotton. A total of 12 regenerable broad leaf lines were obtained and evaluated for the genetic transformation efficiency using the current mature genetic transformation system in cotton; meanwhile, agronomic traits of these lines were investigated. Finally, a family line YE3, with a genetic transformation efficiency of 82.9% and better agronomic traits, was obtained. This study develops a new genotype of upland cotton, which will facilitate the genetic transformation and gene function research of cotton.
cotton; RILs; somatic embryogenesis; regeneration capacity; transformation efficiency
10.3724/SP.J.1006.2024.34167
本研究由國家自然科學基金項目(32172025)資助。
This study was supported by the National Natural Science Foundation of China (32172025).
林忠旭, E-mail: linzhongxu@mail.hzau.edu.cn
E-mail: leyu_hzau@163.com
2023-10-16;
2024-01-12;
2024-02-07.
URL: https://link.cnki.net/urlid/11.1809.S.20240205.1152.008
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/).
猜你喜歡
商品與質量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
中國衛生(2015年12期)2015-11-10 05:13:40
現代企業(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11
