CsAHL25通過影響CsHB1、LEC1/B3基因表達調控柑橘體細胞胚發生
2024-04-30 04:30:41葉長寧徐夢夢劉蘭蘭付玉潔葛曉霞
果樹學報 2024年4期
葉長寧 徐夢夢 劉蘭蘭 付玉潔 葛曉霞

摘? ? 要:【目的】基于柑橘體細胞胚發生相關基因CsHB1的啟動子篩選其上游轉錄因子,以期為柑橘體細胞胚發生分子機制研究提供可靠的候選基因。【方法】利用CsHB1啟動子(-1018~-558 bp)進行酵母單雜篩庫實驗,篩選出CsHB1上游轉錄因子CsAHL25;利用亞細胞定位實驗,確定CsAHL25在細胞中的位置;通過酵母單雜點對點、雙熒光素酶實驗驗證CsAHL25對CsHB1表達的影響;利用qRT-PCR探究CsAHL25基因在柑橘體細胞胚誘導過程中的表達模式;在柑橘愈傷組織中瞬時表達該基因,并檢測體細胞胚發生相關基因的表達變化。【結果】CsAHL25在柑橘體細胞胚誘導過程中呈現先上升后下降的表達模式,該蛋白定位在細胞核中,能與CsHB1啟動子結合并下調CsHB1的表達。瞬時表達CsAHL25會導致CsHB1表達量下調,及CsABI3、CsFUS3、CsLEC1、CsL1L等促進體細胞發生的LEC1/B3基因表達量上調。【結論】CsAHL25能直接下調CsHB1的表達,并使LEC1/B3基因表達量上升。CsAHL25可能通過調整CsHB1、LEC1/B3基因的表達促進體細胞胚發生。
關鍵詞:柑橘:體細胞胚發生;HD-ZIP;AT-HOOK
中圖分類號:S666 文獻標志碼:A 文章編號:1009-9980(2024)04-0579-11
CsAHL25 regulates citrus somatic embryogenesis by affecting the expression of CsHB1 and LEC1/B3 genes
YE Changning1, 3, XU Mengmeng1, 3, LIU Lanlan1, 3, FU Yujie1, 3, GE Xiaoxia2, 3*
(1College of Horticulture and Forestry Sciences, Huazhong Agricultural University, Wuhan 430070, Hubei, China; 2Journal Center of Academy of Science and Technology Development, Huazhong Agricultural University, Wuhan 430070, Hubei, China; 3 Center of Applied Boitechnology, Wuhan Institute of Bioengineering, Wuhan 430415, Hubei, China)
Abstract: 【Objective】Somatic embryogenesis (SE) is widely used in the conservation and utilization of plant germplasm resources. However, there is significant variation in the somatic embryogenesis (SE) capacity of calls derived from different citrus varieties. Furthermore, their SE capacity gradually diminishes during culture, posing a significant hindrance to the conservation and utilization of citrus germplasm resources. CsHB1, an HD-ZIP II gene associated with enhancing SE, was isolated from a citrus variety exhibiting robust SE capabilities. In this study, we harnessed the promoter of CsHB1 (pCsHB1) to search for upstream transcription factors to provide reliable candidate genes for the study on plant somatic embryogenesis. 【Methods】 To identify the upstream transcription factors of CsHB1, we cloned pCsHB1 (-1018 to -558 bp) into pAbAi and utilized a yeast one-hybrid (Y1H) assay to obtain the candidate transcription factor CsAHL25 from a yeast library. Using SMART, candidate genes were analyzed for domains and named based on annotations in the Citrus Pangenome Breeding Database. The expression pattern of this gene was measured by qRT-PCR in various somatic embryo developmental stages of citrus, aiming to deduce the function of CsAHL25. The gene was cloned and inserted into pRI121, transferred into GV3101 and Marker mixed annotated Nicotiana benthamiana. After 2 d, the localization in the cells was observed using the laser scanning confocal microscopy. CsAHL25 was cloned and inserted into pGADT7 and transfected into Y1HGold with pCsHB1-AbAi for the Y1H assay. A Y1H assay was performed to determine whether the two were complementary or not based on the growth of the yeast cells in the screening medium. The gene was cloned and inserted into the overexpression vector pCMBAI1300-35S, and pCsHB1 (-2377-0 bp) was cloned and inserted into pGreenII 0800-LUC. The two vectors were then separately transferred into GV3101 and mixed to transiently transform N. benthamiana, with empty vector used as a control. After 2 d, the fluorescence of LUC was observed using an in vivo Plant Fluorescent Imaging System, and the LUC/REN ratio was calculated. This was followed by a comparison with the control to determine the role of this gene in the downstream gene regulation of pCsHB1. To explore the function of this gene, we transiently expressed the gene in the callus of Citrus sinensis ‘Next, and qRT-PCR was used to detect the expression of somatic embryogenesis-related genes. 【Results】 A candidate transcription factor, named CsAHL25, which is involved in the regulation of CsHB1 expression, was identified from the results of Y1H screening. Sequence analysis revealed that CsAHL25 possesses a typeⅠ AT-HOOK domain and a type A PPC domain and belongs to the AHL15-29 subfamily of AT-HOOK. Subcellular localization analysis demonstrated that, similar to other AHL transcription factors, CsAHL25 is a nucleus-localized transcription factor. CsAHL25 exhibited high expression levels at 60 d and 120 d of somatic cell embryo induction. The expression pattern of CsAHL25 suggested that this gene may play a role in SE. The Y1H results showed that yeast cells containing CsAHL25 and pCsHB1 were able to grow well in SD-Leu/ABA200, indicating that CsAHL25 was bound to pCsHB1. The results of plant fluorescent imaging indicated that 1300+pCsHB1-LUC exhibited higher LUC values than CsAHL25-1300+pCsHB1-LUC. The LUC/REN results were consistent with the Plant Fluorescent Imaging outcomes, with the strongest LUC-related activity observed in 1300+pCsHB1-LUC. These results showed that CsAHL25 was bound to the integral pCsHB1 and repressed its transcription. To further investigate the function of CsAHL25 during SE, we performed transient transfection of CsAHL25 in the callus of C. sinensis ‘Anliucheng, and then analyzed gene expression by using qRT-PCR. The results showed that the expression of CsHB1 was significantly downregulated, while the LEC1/B3 genes promoting somatic embryogenesis, such as CsLEC1, CsL1L, CsFUS3 and CsABI3, were significantly upregulated. 【Conclusion】 The results of this study indicated that CsAHL25 was an upstream transcription factor of CsHB1, which can inhibit CsHB1 expression, and transient expression of CsAHL25 can cause upregulation of the expression of LEC1/B3 genes. Based on the expression pattern of CsAHL25, we studied the functions of HD-ZIP, LEC1/B3 and AHL25 in relation to citrus somatic embryogenesis. Finally, we hypothesized that CsAHL25 regulated citrus somatic embryogenesis.
Key words: Citrus; Somatic embryogenesis; HD-ZIP; AT-HOOK
植物體細胞胚發生是植物體細胞形成胚胎的過程,其發育過程與合子胚相似,均會經歷球形胚、魚雷形胚、子葉形胚等發育階段[1]。隨著體細胞胚誘導技術的發展,該技術已經成為植物種質資源保存與創制、重要經濟植物大規模生產的重要技術手段之一[2]。柑橘是中國的重要經濟作物,近年來,柑橘的種質資源發掘和遺傳改良研究為中國柑橘產業發展提供了有力支持[3]。隨著誘導出來的柑橘品種的胚性愈傷越來越多,利用體細胞進行柑橘種質資源的保存和育種已成為柑橘種質資源利用的重要手段 [3-4]。然而不同柑橘品種的體細胞胚發生能力有著巨大的差異,部分品種至今無法獲得胚性愈傷[5],這對柑橘種質資源保存和育種造成阻礙,因此研究柑橘體細胞胚發生機制有著重要的生物學意義和應用價值。
在體細胞胚發生的過程中,轉錄因子可以通過影響信號轉導調控體細胞胚發生[6]。前期研究表明,LEC[7-9]、FUS[10-12]、ABI[13]、HD-ZIP[14-16]轉錄因子在擬南芥、龍眼、冷杉、柑橘、油棕、紫花苜蓿等多種植物體細胞胚發生過程中起重要作用。其中HD-ZIP轉錄因子是一類植物特有的轉錄因子,具有亮氨酸拉鏈(ZIP)和與之緊密結合的同源結構域,根據HD-ZIP結構域的同源性、蛋白結構和功能可以將HD-ZIP轉錄因子分為4類,其中存在CPSCE結構域的HD-ZIP蛋白被歸類于Ⅱ型HD-ZIP蛋白[17]。在植物胚胎發育的過程中,Ⅱ型HD-ZIP具有調控生長素轉運、維持分生組織、保持植物細胞全能性和控制子葉發育的功能[18- 19]。
AT-HOOK是一類能與DNA序列中富含AT序列區域結合的轉錄因子[20]。其蛋白包含AT-HOOK結構域與PPC結構域兩種保守結構,根據保守結構域的數量和種類可以將AT-HOOK轉錄因子分為兩個亞家族,其中Ⅰ型AT-HOOK蛋白(AHL15-29)包含一個Ⅰ型AHL結構域和一個A型PPC結構域[21-22]。該亞家族成員AHL15、AHL19、AHL20是作用在植物胚胎發生早期的重要轉錄因子,Ⅰ型AT-HOOK基因的表達受生長素和BBM轉錄因子的調控,且具有促進植物體細胞胚發生的功能[23]。雖然前人研究已證實Ⅰ型AT-HOOK轉錄因子具有促進植物胚胎發生的作用,然而Ⅰ型AT-HOOK轉錄因子調控體細胞發生的分子機制仍未被報道。
在對柑橘體細胞胚發生的研究中,前人分離并鑒定出促進柑橘體細胞胚發生的Ⅱ型HD-ZIP轉錄因子CsHB1[24-25]。為了探索柑橘體細胞胚發生相關基因CsHB1的上游調控網絡,筆者對調控CsHB1基因表達的轉錄因子進行挖掘,發現一個屬于Ⅰ型AT-HOOK亞家族的基因CsAHL25,并對其功能進行初步驗證,完善AT-HOOK轉錄因子的調控網絡,為柑橘體細胞胚發生研究提供潛在的候選基因,以期推進柑橘體細胞胚發生的分子機制研究。
1 材料和方法
1.1 試驗材料
MT培養基繼代保存的紐荷爾臍橙(Citrus sinensis ‘Newhall)、暗柳橙(C. sinensis ‘Anliucheng)胚性愈傷組織。
1.2 CsHB1啟動子片段誘餌菌株AbA表達水平檢測
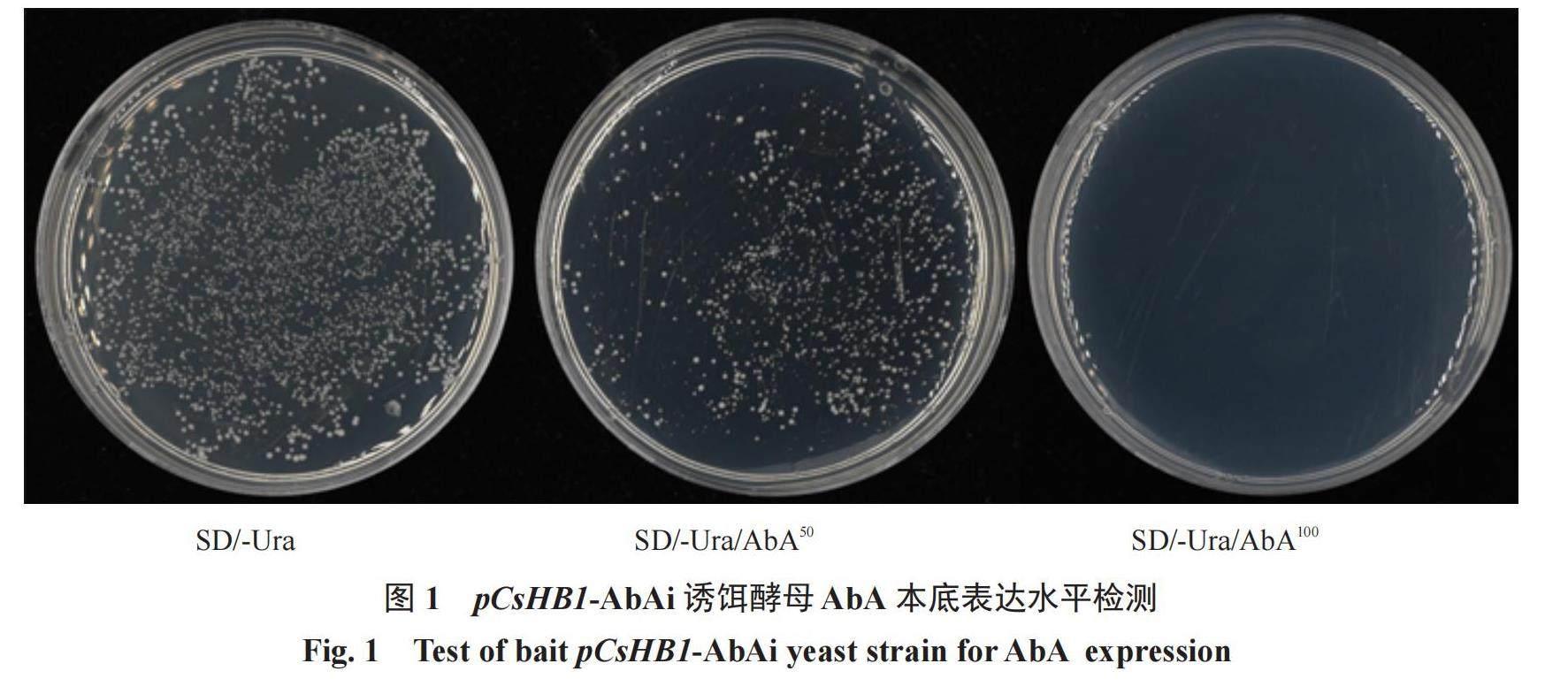
根據距CsHB1基因-1018~-558 bp的啟動子片段序列和pAbAi載體序列設計引物(表1),使用Phanta Max聚合酶(諾唯贊,南京)從暗柳橙DNA中擴增目的片段,使用ClonExpress? Ⅱ同源重組試劑盒(諾唯贊,南京)將目的片段克隆到pAbAi載體中,用BstBⅠ限制性內切酶(新景,杭州)酶切后,使用酵母轉化試劑盒(酷來搏,北京)將載體轉入Y1H Gold酵母細胞中,獲得含有pCsHB1-AbAi的酵母細胞。使用0.9%的NaCl 溶液懸浮酵母細胞(OD600=0.002),取100 μL懸浮菌液分別涂在0、50、100 ng·mL-1 AbA的培養基上,30 ℃倒置培養2?3 d,篩選最適濃度作為酵母單雜篩庫和酵母單雜點對點實驗的AbA濃度。
1.3 酵母單雜篩選CsHB1的上游轉錄因子
使用酵母轉化試劑盒(酷來搏,北京)將各體細胞胚誘導時期的暗柳橙愈傷組織cDNA混合文庫質粒轉入含有pCsHB1-AbAi的Y1H Gold酵母細胞中,使用SD/-Leu/AbA200培養基篩選陽性克隆,使用Taq酶(翌圣,上海)進行陽性克隆鑒定,將鑒定片段大小500?2000 bp的PCR產物進行測序后,使用CPBD數據庫(Citrus Pan-genome to Breeding Database,http://citrus.hzau.edu.cn/)進行比對,分析候選蛋白。
1.4 CsAHL25結構域分析
參考CPBD數據庫中甜橙二代基因組注釋數據和使用SMART(SMART:Main page)工具分析CsAHL25蛋白的氨基酸序列以確定其結構域。
1.5 CsAHL25系統發育樹建立
使用NCBI(https://ncbi.nlm.nih.gov/gene)下載擬南芥(Arabidopsis thaliana)、水稻(Oryza sativa)的Ⅰ型AT-HOOK氨基酸序列。使用MEGA X軟件將擬南芥、水稻的AT-HOOK和CsAHL25的氨基酸序列進行多序列比對,利用鄰接法構建進化樹,Bootstrapping參數值設置為1000次,使用iTOL網站(https://itol.embl.de/)進行數據可視化分析。
1.6 RNA的提取以及cDNA合成
收集甘油培養基(20 mL·L-1)誘導0 d(ALC 0 d)、60 d(ALC 60 d)、120 d(ALC 120 d)的暗柳橙愈傷組織,使用RN38 EASY spin plus植物RNA提取試劑盒(艾德萊,北京)提取組織RNA后,用RT SuperMix反轉錄試劑盒(諾唯贊,南京)合成cDNA。
1.7 實時熒光定量PCR分析
使用Primer Premier 5軟件設計CsAHL25基因的定量引物(表1),以Actin基因作為內參基因。使用LC480實時熒光定量儀器(Roched,美國)和qPCR Master Mix試劑盒(諾唯贊,南京)進行qRT-PCR試驗。
1.8 CsAHL25亞細胞定位分析
根據CsAHL25 CDS序列和pRI121載體序列設計引物(表1),從暗柳橙胚性愈傷組織cDNA中擴增序列,將去除終止密碼子的CsAHL25 CDS序列克隆到pRI121載體中,獲得pRI121-CsAHL25-GFP重組蛋白質粒后轉入GV3101農桿菌中。將含有pRI121-CsAHL25-GFP 農桿菌液和核定位marker H2B-RFP菌液混合后注射煙草葉片,2 d后利用TCS SP8激光共聚焦顯微鏡(Leica,德國)觀察熒光信號并拍照。
1.9 酵母單雜驗證CsAHL25與CsHB1啟動子互作
將CsAHL25 CDS序列克隆到pGADT7載體中(引物見表1),獲得獵物載體pGADT7- CsAHL25并轉入含有pCsHB1-AbAi的酵母細胞中。以空載獵物載體為陰性對照、pAbAi-P53+pGADT7-P53為陽性對照,將獲得的陽性菌株按照10-1梯度稀釋并分別接種在SD/-Leu、SD/-Leu/AbA200培養基上,通過觀察酵母的生長狀態判斷CsAHL25轉錄因子和CsHB1啟動子的互作情況。
1.10 LUC活體成像、雙熒光素酶驗證CsAHL25對CsHB1啟動子活性的影響
將CsHB1啟動子(-2377~0 bp)克隆到pGreenⅡ 0800-LUC載體中(引物見表1),獲得報告子載體pGReenⅡ 0800-LUC-pCsHB1并轉入GV3101(pSoup)農桿菌中,將CsAHL25 CDS序列克隆到pCAMBIA1300-35S載體中作效應子(引物見表1),并轉入GV3103農桿菌中。以報告子空載pGReen Ⅱ 0800-LUC、效應子空載 pCAMBIA1300-35S作為對照,使用煙草瞬時注射將報告子、效應子菌液按體積比1∶5混合后注射到煙草葉片中,2 d后使用熒光素鉀鹽試劑盒(翌圣,上海),通過NightSHADE LB 985植物活體成像系統(Berthold、德國)觀察其LUC熒光強度并拍照,使用雙熒光素酶試劑盒(翌圣,上海),通過Infinite? 200多功能酶標儀(TECAN,瑞士)檢測其雙熒光素酶活性,計算LUC/REN的比值,得到LUC的相對活性。通過LUC熒光強度和LUC/REN的相對比值判斷CsAHL25轉錄因子對CsHB1啟動子活性的影響。
1.11 CsAHL25瞬時超量表達分析
參考張印[26]的方法使用農桿菌介導法將pCAMBIA1300-35s-CsAHL25農桿菌轉入紐荷爾臍橙的愈傷組織中,以轉pCAMBIA1300-35s載體的愈傷組織為空白對照,在含有AS的MT培養基上培養3 d后,提取愈傷組織的RNA進行熒光定量試驗(引物見表1)。
1.12 數據分析
利用GraphPad 8軟件、采用t-test進行顯著性分析并作圖。
2 結果與分析
2.1 CsHB1上游轉錄因子的篩選
為了篩選調控CsHB1表達的轉錄因子,利用CsHB1啟動子片段(-1018~-558 bp)作為誘餌進行酵母單雜篩庫實驗。酵母AbA本底表達水平檢測表明,在SD/-Ura/AbA100固體培養基上,含有pCsHB1-AbAi的Y1H Gold酵母細胞無法正常生長(圖1),最終確定200 ng·mL-1作為酵母單雜篩庫的AbA質量濃度值。酵母單雜篩庫結果表明,共獲得56個酵母克隆,使用PCR鑒定后得到39個插入片段大小在500~2000 bp之間,且條帶單一的cDNA片段。測序結果使用CPBD數據庫進行Blastx比對分析,去除假陽性克隆后,初步獲得21個可能與CsHB1啟動子片段結合的蛋白(表2),包括含有AT-HOOK結構域的核定位蛋白、含有RING指和CHY鋅指結構的蛋白1、晚期胚胎發生豐富蛋白D-7等。
2.2 CsAHL25基因系統發育與表達模式分析
根據陽性克隆出現次數和相關報道,選擇注釋為含有AT-HOOK結構域的核定位蛋白Cs8g_pb000150進行研究。Cs8g_pb000150基因CDS長度為918 bp,編碼305個氨基酸,進一步分析發現在其61~77 aa處存在一個Ⅰ型AHL結構域、89~214 aa處存在一個A型PPC結構域(圖2-A),其結構域具有Ⅰ型AT-HOOK轉錄因子的特征。該蛋白與擬南芥、水稻Ⅰ型AT-HOOK轉錄因子進化關系分析表明,該蛋白與AtAHL25、OsAHL25在同一個進化支中(圖2-B)。根據CPBD數據庫注釋與進化分析將該蛋白命名為CsAHL25。
qRT-PCR檢測CsAHL25基因在體細胞胚誘導過程中的表達模式,發現在暗柳橙愈傷生胚誘導過程中,CsAHL25的相對表達量隨著誘導時間的延長呈先上升后下降的趨勢(圖2-C),其表達模式暗示CsAHL25基因可能在柑橘的體細胞誘導過程中發揮作用。
2.3 CsAHL25蛋白定位分析
為了檢測CsAHL25蛋白在細胞中的定位情況,在煙草葉片中瞬時表達CsAHL25-GFP熒光蛋白和H2B-RFP核marker蛋白,激光共聚焦觀察發現與空載對照組相比,CsAHL25-GFP熒光信號集中在細胞核中,并與核marker(H2B-RFP)熒光信號均重疊(圖3),結果表明CsAHL25蛋白定位于細胞核中,具有AT-HOOK轉錄因子的定位特征。
2.4 CsAHL25轉錄因子與CsHB1啟動子互作分析
為了確認CsAHL25轉錄因子與CsHB1啟動子的結合,使用酵母單雜點對點實驗對CsAHL25轉錄因子與pCsHB1啟動子片段(-1018~-558 bp)的結合進行驗證,含有pGADT7-CsAHL25和pCsHB1-AbAi質粒的酵母細胞和陽性對照能在互作篩選培養基(SD/-Leu/AbA200)上正常生長,陰性對照無法正常生長(圖4-A),結果表明在酵母細胞中,CsAHL25具有結合pCsHB1啟動子片段的能力。
為了進一步驗證CsAHL25轉錄因子與pCsHB1啟動子(-2377~0 bp)的互作,進行植物活體成像實驗和雙熒光素酶實驗,驗證其互作關系。植物活體成像結果顯示,pCAMBIA1300-35S空載+pCsHB1-LUC組合的LUC熒光值顯著高于CsAHL25-1300+pCsHB1-LUC組合與陰性對照(圖4-B)。雙熒光素酶活性測定結果與植物活體成像結果相同,pCAMBIA1300-35S空載+pCsHB1-LUC組合的LUC相對活性顯著高于CsAHL25-1300+pCsHB1-LUC組合(圖4-C)。上述實驗結果表明,CsAHL25轉錄因子能與pCsHB1啟動子結合并抑制下游基因的表達。
2.5 CsAHL25瞬時表達愈傷系中體細胞胚發生相關基因表達分析
為了進一步探索CsAHL25基因的功能,在紐荷爾臍橙的愈傷組織中瞬時表達CsAHL25。以轉pCAMBIA1300-35S空載的愈傷組織為對照,分析已報道的體細胞胚發生相關基因的表達量變化。結果表明,CsAHL25相對表達量均高于或極顯著高于對照組(圖5-A),且CsHB1表達量均極顯著低于對照組(圖5-B)。CsLEC1、CsL1L、CsFUS3、CsABI3等參與柑橘體細胞胚發生的LEC1/B3基因相對表達量相較對照組均顯著上升,且其表達量變化趨勢與CsAHL25基本一致(圖5-C~F)。以上結果表明,CsAHL25轉錄因子能下調CsHB1表達,并影響LEC1/B3調控網絡相關基因表達。
3 討 論
筆者篩選并鑒定到一個直接調控柑橘體細胞發生相關基因CsHB1的Ⅰ型AT-HOOK轉錄因子CsAHL25。CsAHL25是經過酵母單雜篩庫篩選得到的轉錄因子,是調控CsHB1表達的候選基因之一。經過酵母單雜點對點和雙熒光素酶實驗驗證,發現CsAHL25可以直接與CsHB1啟動子結合,進而下調CsHB1的表達。前期研究發現,Ⅰ型AT-HOOK轉錄因子具有調控植物體細胞胚胎發生的功能,通過過表達AHL15基因,可以誘導擬南芥幼苗直接形成體細胞胚胎,且ahl15 ahl19 amiRAHL20三重突變的擬南芥植株完全無法誘導出體細胞胚[22]。本研究中鑒定出的柑橘Ⅰ型AT-HOOK轉錄因子CsAHL25,具有和柑橘體細胞胚發生相關的表達模式,該轉錄因子能夠調控體細胞胚發生相關基因的表達,從而影響柑橘體細胞胚發生,與已報道的擬南芥Ⅰ型AT-HOOK轉錄因子功能相似。Ⅰ型AT-HOOK轉錄因子能夠通過調控GA3OX1基因表達影響GA的合成[27]、通過調控PFI基因表達影響下胚軸的伸長[28]、通過調控SPL基因表達影響植物的壽命[29]。目前Ⅰ型AT-HOOK轉錄因子促進植物體細胞胚胎發生的分子機制尚未明確,且暫無研究表明Ⅰ型AT-HOOK轉錄因子調控Ⅱ型HD-ZIP基因表達,本研究中初步證明,Ⅰ型AT-HOOK轉錄因子調控Ⅱ型HD-ZIP基因的表達,完善了Ⅰ型AT-HOOK轉錄因子的下游調控網絡。
此外,筆者利用瞬時表達實驗發現,除了直接下調CsHB1基因的表達外,CsAHL25還影響了LEC1/B3基因的表達。瞬時表達CsAHL25基因會導致CsLEC1、CsL1L、CsFUS3、CsABI3基因的表達量上升,對體細胞胚發生相關基因呈現不同的調控方式。LEC1-FUS3-LEC2-ABI3基因共同構成一個LEC1/B3結構域調控網絡,該網絡通過調控體細胞胚的形態構成,進而影響植物體細胞胚的發生[30-32]。前人通過分析柑橘體細胞胚發生過程中基因的表達模式,明確LEC1、LEC1 Like、FUS3、ABI3等LEC1/B3調控網絡基因在保持柑橘愈傷胚性、促進其胚胎發育中起到了重要的作用[33-34]。進一步研究發現,在柑橘愈傷組織分化的過程中,CsFUS3基因相對表達量逐漸上升,超表達CsFUS3會引起愈傷細胞形態變化、激活體細胞胚發生[35]。過表達CsL1L基因也能夠使柑橘的營養組織產生體細胞胚[36]。本研究表明,瞬時表達CsAHL25上調LEC1、L1L、FUS3、ABI3基因的表達量,說明CsAHL25可能通過影響LEC1/B3表達,調控體細胞胚發生的功能。LEC1/B3調控網絡基因能夠具有細胞發生形態轉變、調控體細胞胚形態構建的作用[32,35],而Ⅱ型HD-ZIP基因作用在植物胚胎發生前期,具有維持植物胚胎中干細胞存在的功能[19,37],二者作用在植物胚胎發生過程中的不同方面,其相互關系尚不明確,有待深入研究。此外,研究發現CsFUS3基因可以下調細胞中GA的含量,導致ABA/GA比例上升,從而促進柑橘體細胞胚發育[35],且AHL25基因也具有下調植物中GA含量的功能[27],CsAHL25是否可以通過影響柑橘體內ABA/GA比例來促進柑橘體細胞胚發生還需進一步探討。
4 結 論
筆者通過酵母單雜點對點實驗、雙熒光素酶實驗等篩選到柑橘體細胞胚發生相關基因CsHB1的上游抑制因子CsAHL25,通過瞬時表達CsAHL25確認其能夠下調CsHB1表達量并上調LEC1、LEC1 Like、FUS3、ABI3等LEC1/B3基因相對表達量。筆者認為該基因具有激活柑橘體細胞胚發生的功能,為柑橘體細胞胚發生研究提供了一個潛在的候選基因。
參考文獻 References:
[1] MORDHORST A P,TOONEN M A J,DE VRIES S C,MEINKE D. Plant embryogenesis[J]. Critical Reviews in Plant Sciences,1997,16(6):535-576.
[2] SIDDIQUI Z H,ABBAS Z K,ANSARI M W,KHAN M N. The role of miRNA in somatic embryogenesis[J]. Genomics,2019,111(5):1026-1033.
[3] 郭文武,葉俊麗,鄧秀新. 新中國果樹科學研究70年:柑橘[J]. 果樹學報,2019,36(10):1264-1272.
GUO Wenwu,YE Junli,DENG Xiuxin. Fruit scientific research in New China in the past 70 years:Citrus[J]. Journal of Fruit Science,2019,36(10):1264-1272.
[4] 鄧秀新. 中國柑橘育種60年回顧與展望[J]. 園藝學報,2022,49(10):2063-2074.
DENG Xiuxin. A review and perspective for citrus breeding in China during the last six decades[J]. Acta Horticulturae Sinica,2022,49(10):2063-2074.
[5] 劉丹. 柑橘優異資源胚性愈傷組織誘導及體細胞雜種創制[D]. 武漢:華中農業大學,2019.
LIU Dan. Induction of nucellar embryogenic callus and generation of somatic hybrid by protoplast fusion in citrus[D]. Wuhan:Huazhong Agricultural University,2019.
[6] ELHITI M,STASOLLA C. Transduction of signals during somatic embryogenesis[J]. Plants,2022,11(2):178.
[7] 蔡英卿,賴鐘雄,陳義挺,林玉玲,李惠華,張妙霞. 龍眼胚性愈傷組織LEC1基因cDNA克隆以及在體胚發生過程中的表達分析[J]. 福建農林大學學報(自然科學版),2011,40(5):494-500.
CAI Yingqing,LAI Zhongxiong,CHEN Yiting,LIN Yuling,LI Huihua,ZHANG Miaoxia. Cloning of LEC1 gene from embryogenic callus and its expression analysis during somatic embryogenesis in longan[J]. Journal of Fujian Agriculture and Forestry University (Natural Science Edition),2011,40(5):494-500.
[8] VETRICI M A,YEVTUSHENKO D P,MISRA S. Overexpression of douglas-fir LEAFY COTYLEDON1 (PmLEC1) in Arabidopsis induces embryonic programs and embryo-like structures in the lec1-1 mutant but not in wild type plants[J]. Plants,2021,10(8):1526.
[9] KIM H U,JUNG S J,LEE K R,KIM E H,LEE S M,ROH K H,KIM J B. Ectopic overexpression of castor bean LEAFY COTYLEDON2 (LEC2) in Arabidopsis triggers the expression of genes that encode regulators of seed maturation and oil body proteins in vegetative tissues[J]. FEBS Open Bio,2013,4:25-32.
[10] LEDWO? A,GAJ M D. LEAFY COTYLEDON1,FUSCA3 expression and auxin treatment in relation to somatic embryogenesis induction in Arabidopsis[J]. Plant Growth Regulation,2011,65(1):157-167.
[11] WANG F F,PERRY S E. Identification of direct targets of FUSCA3,a key regulator of Arabidopsis seed development[J]. Plant Physiology,2013,161(3):1251-1264.
[12] LIU Z,GE X X,QIU W M,LONG J M,JIA H H,YANG W,DUTT M,WU X M,GUO W W. Overexpression of the CsFUS3 gene encoding a B3 transcription factor promotes somatic embryogenesis in Citrus[J]. Plant Science,2018,277:121-131.
[13] CHEN B J,FIERS M,DEKKERS B J W,MAAS L,VAN ESSE G W,ANGENENT G C,ZHAO Y,BOUTILIER K. ABA signalling promotes cell totipotency in the shoot apex of germinating embryos[J]. Journal of Experimental Botany,2021,72(18):6418-6436.
[14] KHIANCHAIKHAN K,AROONLUK S,VUTTIPONGCHAIKIJ S,JANTASURIYARAT C. Genome-wide identification of homeodomain leucine zipper (HD-ZIP) transcription factor,expression analysis,and protein interaction of HD-ZIP IV in oil palm somatic embryogenesis[J]. International Journal of Molecular Sciences,2023,24(5):5000.
[15] GE X X,LIU Z,WU X M,CHAI L J,GUO W W. Genome-wide identification,classification and analysis of HD-ZIP gene family in citrus,and its potential roles in somatic embryogenesis regulation[J]. Gene,2015,574(1):61-68.
[16] HU X,ZHANG C R,XIE H,HUANG X,CHEN Y F,HUANG X L. The expression of a new HD-Zip Ⅱ gene,MSHB1,involving the inhibitory effect of thidiazuron on somatic embryogenic competence in alfalfa (Medicago sativa L. cv. Jinnan) callus[J]. Acta Physiologiae Plantarum,2012,34(3):1067-1074.
[17] TRON A E,BERTONCINI C W,CHAN R L,GONZALEZ D H. Redox regulation of plant homeodomain transcription factors[J]. The Journal of Biological Chemistry,2002,277(38):34800-34807.
[18] TURCHI L,CARABELLI M,RUZZA V,POSSENTI M,SASSI M,PE?ALOSA A,SESSA G,SALVI S,FORTE V,MORELLI G,RUBERTI I. Arabidopsis HD-Zip Ⅱ transcription factors control apical embryo development and meristem function[J]. Development,2013,140(10):2118-2129.
[19] ROODBARKELARI F,GROOT E P. Regulatory function of homeodomain-leucine zipper (HD-ZIP) family proteins during embryogenesis[J]. The New Phytologist,2017,213(1):95-104.
[20] PRAVEEN S,PAWAR V,AHLAWAT Y S. Somatic embryogenesis and plant regeneration in Kinnow mandarin[J]. Journal of Plant Biochemistry and Biotechnology,2003,12(2):163-165.
[21] ARAVIND L,LANDSMAN D. AT-hook motifs identified in a wide variety of DNA-binding proteins[J]. Nucleic Acids Research,1998,26(19):4413-4421.
[22] ZHAO J F,FAVERO D S,PENG H,NEFF M M. Arabidopsis thaliana AHL family modulates hypocotyl growth redundantly by interacting with each other via the PPC/DUF296 domain[J]. Proceedings of the National Academy of Sciences of the United States of America,2013,110(48):E4688-E4697.
[23] KARAMI O,RAHIMI A,MAK P,HORSTMAN A,BOUTILIER K,COMPIER M,VAN DER ZAAL B,OFFRINGA R. An Arabidopsis AT-hook motif nuclear protein mediates somatic embryogenesis and coinciding genome duplication[J]. Nature Communications,2021,12:2508.
[24] 謝幸男,賴曉娜,戰爽,程來超,許全全,葛曉霞. 柑橘體細胞胚發生基因CsHB1特異肽段多克隆抗體的制備及其蛋白動態檢測[J]. 果樹學報,2017,34(9):1069-1075.
XIE Xingnan,LAI Xiaona,ZHAN Shuang,CHENG Laichao,XU Quanquan,GE Xiaoxia. Preparation of CsHB1 polyclonal antibody and its protein dynamic changes during somatic embryogenesis in Citrus[J]. Journal of Fruit Science,2017,34(9):1069-1075.
[25] 劉娉婷. 基于柑橘強胚性材料開展CsHB1促體細胞胚發生方式的研究[D]. 武漢:華中農業大學,2022.
LIU Pingting. Study the CsHB1 promotion of somatic embryogenesis uasing citrus strong embryogenic materials[D]. Wuhan:Huazhong Agricultural University,2022.
[26] 張印. 柑橘原花青素積累及ABA代謝的調控機制研究[D]. 武漢:華中農業大學,2021.
ZHANG Yin. Research on the regulation mechanism of proanthocyanidin accumulation and ABA metabolism in citrus[D]. Wuhan:Huazhong Agricultural University,2021.
[27] MATSUSHITA A,FURUMOTO T,ISHIDA S,TAKAHASHI Y. AGF1,an AT-hook protein,is necessary for the negative feedback of AtGA3ox1 encoding GA 3-oxidase[J]. Plant Physiology,2007,143(3):1152-1162.
[28] XIAO C W,CHEN F L,YU X H,LIN C T,FU Y F. Over-expression of an AT-hook gene,AHL22,delays flowering and inhibits the elongation of the hypocotyl in Arabidopsis thaliana[J]. Plant Molecular Biology,2009,71(1):39-50.
[29] RAHIMI A,KARAMI O,BALAZADEH S,OFFRINGA R. miR156-independent repression of the ageing pathway by longevity-promoting AHL proteins in Arabidopsis[J]. The New Phytologist,2022,235(6):2424-2438.
[30] HORSTMAN A,LI M F,HEIDMANN I,WEEMEN M,CHEN B J,MUINO J M,ANGENENT G C,BOUTILIER K. The BABY BOOM transcription factor activates the LEC1-ABI3-FUS3-LEC2 network to induce somatic embryogenesis[J]. Plant Physiology,2017,175(2):848-857.
[31] LOTAN T,OHTO M A,YEE K M,WEST M A L,LO R,KWONG R W,YAMAGISHI K,FISCHER R L,GOLDBERG R B,HARADA J J. Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells[J]. Cell,1998,93(7):1195-1205.
[32] KWONG R W,BUI A Q,LEE H,KWONG L W,FISCHER R L,GOLDBERG R B,HARADA J J. LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development[J]. The Plant Cell,2003,15(1):5-18.
[33] GE X X,CHAI L J,LIU Z,WU X M,DENG X X,GUO W W. Transcriptional profiling of genes involved in embryogenic,non-embryogenic calluses and somatic embryogenesis of Valencia sweet orange by SSH-based microarray[J]. Planta,2012,236(4):1107-1124.
[34] 繆星辰. 椪柑體細胞胚胎發生關鍵基因的挖掘與鑒定[D]. 揚州:揚州大學,2023.
MIAO Xingchen. Mining and identification of key genes involved in somatic embryogenesis in ponkan[D]. Yangzhou:Yangzhou University,2023.
[35] 劉政. 柑橘珠心胚起始轉錄組分析及體細胞胚發生相關基因CsFUS3功能鑒定[D]. 武漢:華中農業大學,2015.
LIU Zheng. Transcriptional analysis of citrus nucellar embryo initiation and functional characterization of CsFUS3 gene preferentially expressed during somatic embryogenesis[D]. Wuhan:Huazhong Agricultural University,2015.
[36] ZHU S P,WANG J,YE J L,ZHU A D,GUO W W,DENG X X. Isolation and characterization of LEAFY COTYLEDON 1-LIKE gene related to embryogenic competence in Citrus sinensis[J]. Plant Cell,Tissue and Organ Culture,2014,119(1):1-13.
[37] TURCHI L,BAIMA S,MORELLI G,RUBERTI I. Interplay of HD-Zip Ⅱ and Ⅲ transcription factors in auxin-regulated plant development[J]. Journal of Experimental Botany,2015,66(16):5043-5053.
